
Effects of UV-B radiation intensity and timing on epidemiological components of wheat stripe rust
2018-12-11 08:38WANGHuiQlNFengCHENGPeiMAZhanhongWANGHaiguang
Journal of Integrative Agriculture 2018年12期
WANG Hui , QlN Feng CHENG Pei MA Zhan-hong WANG Hai-guang
1 Department of Plant Pathology, China Agricultural University, Beijing 100193, P.R.China
2 Rizhao Agricultural Bureau of Shandong Province, Rizhao 276800, P.R.China
Abstract Stripe rust caused by Puccinia striiformis f. sp. tritici is an important wheat disease worldwide that is greatly influenced by environmental conditions. Ultraviolet B (UV-B) radiation is one important environmental factor affecting the occurrence and epidemiology of wheat stripe rust. Investigating UV-B radiation effects on the epidemiology of stripe rust may be conducive to monitoring and predicting this disease. In this study, wheat seedlings were exposed to UV-B radiation during different periods under laboratory conditions and radiation effects on epidemiological components of wheat stripe rust were investigated.Results showed that incubation period was shortened, and the infection efflciency, sporulation quantity and disease index increased when UV-B radiation was performed only pre-inoculation. When the UV-B radiation was performed only postinoculation or both pre- and post-inoculation, the incubation period was prolonged, and the infection efflciency, sporulation quantity and disease index were reduced. When healthy wheat seedlings were inoculated using urediospores collected from wheat leaves irradiated by UV-B only post-inoculation or both pre- and post-inoculation, infection efflciency, sporulation quantity and disease index were also reduced. However, in the latter, the disease incubation period did not differ under varying UV-B radiation intensities compared to that when wheat leaves were not treated with UV-B radiation. Overall, the effects of direct exposure of wheat plants to UV-B radiation with different intensities in different periods on epidemiological components of wheat stripe rust were systematically explored, and the results suggest that the effects of UV-B radiation increased gradually with the increase of UV-B radiation intensity. This information provides a basis for monitoring and predicting this disease as well as for conducting further studies on pathogen virulence variation.
Keywords: wheat stripe rust, UV-B radiation, epidemiological component, Puccinia striiformis f. sp. tritici
1. lntroduction
Wheat stripe rust is caused by airbornePuccinia striiformisf. sp.tritici(Pst) and is an important fungal wheat disease worldwide (Li and Zeng 2002; Line 2002; Wanet al. 2004,2007; Chen 2005; Chenet al. 2014; Wanget al. 2014).Wheat stripe rust in China has high epidemic frequency,can be very destructive, and results in great losses in wheat production during epidemic years. In particular, wheat yield losses of 6.0, 3.2, 1.8 and 1.3 million tons resulted from pandemics of wheat stripe rust in China in 1950, 1964,1990 and 2002, respectively (Li and Zeng 2002; Wanet al.2004, 2007). Occurrence and prevalence of this disease is closely related to national food security in China (Li and Zeng 2002; Chenet al. 2014).
AlthoughPstsexual reproduction can occur on alternate hosts (e.g.,Berberisspp.) in China (Zhaoet al. 2013), its contribution to regional epidemics of wheat stripe rust is not clear. Wheat stripe rust pathogen mainly relies on urediospores to complete the annual disease cycle on wheat in China (Li and Zeng 2002; Wanet al. 2007; Chenet al. 2014; Wanget al. 2014). The epidemic of wheat stripe rust depends mainly on the susceptibility of cultivated varieties, the pathogenicity of overwinteringPst, rainfall and temperature (Li and Zeng 2002). Furthermore, susceptibility of cultivated varieties andPstpathogenicity are greatly influenced by environmental factors such as temperature,relative humidity and light (Li and Zeng 2002). Ultraviolet B(UV-B: 280–320 nm) radiation is an important environmental factor that affects both host plants and pathogens (Manning and Tiedemann 1995). In particular, wheat stripe rust pathogen is influenced by UV radiation during long-distance dispersal (Li and Zeng 2002). Moreover, the oversummering areas ofPstin China are mainly located in areas with high altitude, where UV-B radiation has high intensity which may affect the survival and genetic variation ofPst.
Among UV components of solar light, UV-B is only partially absorbed by the atmosphere, thus most UV-B radiation reaches the Earth’s surface, affecting many organisms(Madronichet al. 1998; Costaet al. 2012; Escobar-Bravoet al. 2017; Takshak and Agrawal 2017). UV-B radiation can impact morphology, physiology and biochemistry of host plants (Teramura 1983; Barneset al. 1990; Stridet al.1994; Sharmaet al. 1998; Kakaniet al. 2003; Caiet al. 2007;Wanget al. 2010; Xu and Sullivan 2010; Singhet al. 2011;Lidon 2012; Yuet al. 2013; Robsonet al. 2015), which may result in abnormal plant growth and may also impact plant diseases. UV-B radiation can impact growth, development,and physiological and biochemical characteristics of plant pathogens, and can also impact pathogenicity, thus affecting the occurrence of plant diseases (Manning and Tiedemann 1995; Bragaet al. 2015).
Responses of plant pathogens to UV-B radiation differ(Manning and Tiedemann 1995; Bragaet al. 2015). For most of pathogens, UV-B radiation reduces the germination rate (Aylor and Sanogo 1997; Arabi and Jawhar 2003;Costaet al. 2012; Liet al. 2018), mycelial growth rate(Willocquetet al. 1996; Fourtouniet al. 1998; Fenget al.2010; Costaet al. 2012; Yamagaet al. 2016; Liet al.2018) and sporulation (Fourtouniet al. 1998; Fenget al.2010; de Menezeset al. 2015; Liet al. 2018). However,short-term and low-dose UV-B radiation may increase the mycelial growth rate of some plant pathogens (Wuet al. 2008). The effects of UV radiation onPstand wheat stripe rust were previously reported. Jinget al. (1993)investigated the effects of UV radiation onPstusing artiflcial inoculation of UV-irradiated urediospores of two Chinese physiological races ofPst(CYR19 and CYR29) on the highly susceptible wheat cultivar Huixianhong, and found that UV radiation could severely reduce the germinability of the urediospores and weaken the pathogenicity ofPst.Chenget al. (2014) looked at effects of UV-B radiation on epidemiological components of wheat stripe rust caused by three physiological races ofPst(CYR31, CYR32 and CYR33). The results demonstrated that UV-B radiation prolonged the incubation period, reduced the germination rate and infection efflciency of each physiological race,and caused the decreases in the lesion expansion rate,sporulation quantity and area under disease progress curve(AUDPC). Furthermore, the mutation of pathogenicity or virulence variation inPstmay be induced by UV radiation(Shanget al. 1994; Huanget al. 2005; Wanget al. 2009;Huet al. 2014; Kanget al. 2015). Shanget al. (1994)obtained seven virulence mutant strains from UV-irradiated urediospores of thePstphysiological race CYR29 on seven screening wheat cultivars. Studies on UV-induced virulence mutant strains from thePstphysiological races CYR29(Huanget al. 2005) and CYR23 (Wanget al. 2009) at the molecular level using the random amplifled polymorphic DNA technique showed differences in DNA polymorphisms between the mutants and wild strains.
The effects of plant UV-B radiation exposure on plant diseases are related to the timing of UV-B radiation (Orthet al. 1990). In general, plant diseases can be aggravated by pre-inoculation UV-B radiation, and the effects of post-inoculation UV-B radiation on plant diseases vary by crop cultivar and disease species (Wargentet al. 2006).However, pre-inoculation UV-B radiation can reduce the occurrence of some plant diseases. For example, Wargentet al. (2006) demonstrated that exposure of lettuce to UV-B radiation prior to inoculation with a conidial suspension ofBremia lactucaereduced the occurrence of lettuce downy mildew and the sporulation quantity of the pathogen.Moreover, under different conditions, the effects of UV-B radiation on plant diseases differ by cultivar. Liet al. (2008)conducted a study to investigate the effects of enhanced UV-B radiation timing on stripe rust in three wheat cultivars(Mingxian 169, Dunbanmai and Xiannong 4) infected with CYR32. When pre-inoculation UV-B radiation on wheat seedlings was performed, the disease index increased in the two susceptible cultivars (Mingxian 169 and Dunbanmai),but not in the resistant cultivar (Xiannong 4). No effects were detected on the three cultivars when post-inoculation UV-B radiation was performed. In an experiment conducted by Hanet al. (2011), UV-B radiation on seedlings of four wheat cultivars (Mingxian 169, Jing 9428, Beijing 0045 and Jingdong 8) was performed after a 24 h post-inoculation, and it was demonstrated that UV-B radiation had no effect on wheat stripe rust in terms of infection efflciency, incubation period, lesion expansion rate and AUDPC.
Since occurrence and epidemics of wheat stripe rust are greatly influenced by environmental factors (Li and Zeng 2002), it is of great importance to investigate the effects of environmental factors on this disease in order to improve monitoring, forecasting and management of the disease.As described above, virulence variation of plant pathogens as well as occurrence and development of plant diseases may be greatly influenced by UV-B radiation. Most previous studies have focused on the effects of UV-B radiation on plant pathogens or plant growth, but few studies have been conducted to investigate the effects on diseases after host plants were directly exposed to UV-B radiation. In particular,systematic studies on the effects of direct exposure of wheat plants to UV-B radiation on wheat stripe rust have rarely been reported.
The effects of UV-B radiation intensity and timing on epidemiological components of wheat stripe rust including incubation period, infection efflciency, sporulation quantity and disease index by direct exposure of wheat seedlings to UV-B radiation, were investigated in this study to explore direct exposure effects of wheat hosts to UV-B radiation on the disease, to provide some basis for disease monitoring and prediction, and to better understand virulence variation mechanisms ofPst.
2. Materials and methods
2.1. Materials
CYR33, a dominant physiological race ofPstin China, was stored in liquid nitrogen in the Plant Disease Epidemiology Laboratory, Department of Plant Pathology, China Agricultural University in Beijing, China. Mingxian 169, a wheat cultivar susceptible to all known physiological races ofPstin China, was grown as the experimental host material.Using methods previously described by Chenget al.(2014), urediospores of CYR33 were multiplied on wheat seedlings grown in a climate-controlled growth chamber at a temperature of 11–13°C and 60–70% relative humidity under 12 h light at 10 000 lux per day.
UV-B radiation was performed with 36 W UV313 lamps(Beijing Lighting Research Institute, Beijing, China) in the growth chamber described above. The UV-B radiation intensity was measured using a UV-B 297 radiation meter(Beijing Normal University Photoelectric Instrument Factory,Beijing, China).
2.2 UV-B radiation methods
Plump seeds of wheat cultivar Mingxian 169 were soaked for 24 h in sterile water. Well germinated seeds were selected and sown in 10-cm-diameter pots at approximately 20 seeds per pot. Pots were then incubated in the growth chamber.
Five UV-B radiation treatments (Treatments 1–5) were conducted as described below. Within each treatment,three UV-B intensity sub-treatments were performed and non-UV-B irradiated seedlings were used as controls (CK).Sub-treatments were performed in three replicated sets (flve pots per replicate). The UV-B radiation intensities were set by adjusting the distances between wheat seedlings and UV lamps. Three radiation distances were used in this study, 60,50 and 40 cm, with corresponding UV-B radiation intensities of 12, 17.5 and 23.3 µW cm–2, respectively. Epidemiological components investigated in this study included incubation period, infection efflciency, sporulation quantity and disease index.
Treatment 1: pre-inoculation UV-B radiationThe UV-B radiation began on the 6th day post sowing (6 h per day from 10:00 to 16:00, Beijing Time) and continued for 7 consecutive days. At that time, wheat seedlings were inoculated with aPsturediospore suspension of 0.15 mg mL–1using a spray inoculation method. Inoculated seedlings were immediately transferred into a moist chamber with 100% relative humidity without light and incubated at 11–13°C for 24 h. Seedlings were then transferred into the growth chamber under the conditions described above and incubated.
Treatment 2: post-inoculation UV-B radiationOn the 13th day post sowing, wheat seedlings were inoculated,incubated, and transferred to the growth chamber as described above, where they then underwent UV-B radiation as described in Treatment 1 until investigations of epidemiological components were completed.
Treatment 3: UV-B radiation both pre- and post-inoculationThis treatment was performed as in Treatment 1,but with an additional UV-B treatment applied post inoculation (conducted as described in Treatment 1 until all investigations of epidemiological components were completed) immediately after wheat seedlings were transferred to the growth chamber.
Treatment 4: untreated seedlings inoculated withPsturediospores from leaves treated by UV-B radiation post-inoculationInitially, this treatment was performed as in Treatment 2. On the 8th day post sporulation, fresh urediospores from the surface of diseased leaves were collected and inoculated onto healthy wheat seedlings which were then incubated and transferred to the growth chamber as described above (UV-B radiation treatment was not applied to these plants). These plants were used to investigate epidemiological components.
Treatment 5: untreated seedlings inoculated with Psturediospores from leaves treated by UV-B radiation both pre- and post-inoculationThis treatment was performed as in Treatment 3, but with subsequent inoculation of healthy plants as described in Treatment 4 using urediospores collected from plants in the flrst phase of this treatment.The newly inoculated wheat seedlings were not treated with UV-B radiation and were used to investigate epidemiological components.
2.3. lnvestigation methods of epidemiological components
The epidemiological components, including incubation period, infection efflciency, sporulation quantity and disease index, were investigated using methods previously described by Chenget al. (2014). To assess the incubation period, after chlorotic spots appeared on the inoculated wheat leaves, leaves were observed daily or after UV-B radiation at 16:00 every day, and the day on which the flrst uredinium ruptured was recorded. Sporulation quantity was determined for three replicates of irradiated seedlings for each of the three UV-B intensities and flve treatments.For each replicate, leaves were observed for sporulation and urediospores on an individual leaf marked with a label were collected using a flnger-shaped tube every other day until no more spores were produced. The average sporulation quantity was calculated across replicates. The total sporulation quantity under certain UV-B intensity or the control in each treatment was calculated by summing the average sporulation quantities across the entire sporulation period. Plants were surveyed for disease incidence and severity every 5 days post symptom appearance, and the disease index was calculated, all according to Chenget al.(2014). In this study, all statistical analyses of the signiflcant differences at the 0.05 level were performed.
3. Results
3.1. Effects of UV-B radiation on incubation period of wheat stripe rust

Fig. 1 The effects of different UV-B radition distances (40, 50 and 60 cm) and non-UV-B (CK) on incubation period of wheat stripe rust under different UV-B radiation treatments. Treatment 1,pre-inoculation UV-B radiation; Treatment 2, post-inoculation UV-B radiation; Treatment 3, UV-B radiation both pre- and post-inoculation; Treatment 4, untreated seedlings inoculated with Puccinia striiformis f. sp. tritici (Pst) urediospores from leaves treated by UV-B radiation post-inoculation; Treatment 5, untreated seedlings inoculated with Pst urediospores from leaves treated by UV-B radiation both pre- and post-inoculation.Letters indicate signiflcance groupings at the 0.05 level (n=3).
The effects of different UV-B radiation distances on the incubation period of wheat stripe rust across all treatments are shown in Fig. 1. The incubation period for all non-UV-B irradiated control plants was 12 days. The incubation period was shorter by 2 days and 1 day under UV-B radiation distances of 40 cm and 50 or 60 cm, respectively, compared to the control in Treatment 1 (P≤0.05). Under Treatment 1,there was a signiflcant difference in incubation period under the UV-B radiation distance of 40 cm compared to the UV-B radiation distance of 60 or 50 cm. The incubation period was longer by 2 days and 1 day for 40 cm and 50 or 60 cm, respectively, compared to the control in Treatment 2(P≤0.05). Under Treatment 2, signiflcant differences(P≤0.05) in incubation period were observed under the UV-B radiation distance of 40 cm compared to the UV-B radiation distances of 50 and 60 cm. For Treatment 3, the incubation periods for UV-B irradiated plants under distances of 60 and 50 cm did not differ from the control, but was longer by 1 day under a UV-B radiation distance of 40 cm (P≤0.05).For Treatment 4, the incubation period for UV-B irradiated plants under a distance of 60 cm did not differ from the control, but was longer by 1 day under radiation distance of 50 or 40 cm (P≤0.05). For Treatment 5, the incubation period was 12 days for all UV-B radiation distances.
3.2. Effects of UV-B radiation on infection efficiency of Pst
The effects of different UV-B radiation distances on the infection efflciency ofPstunder each UV-B radiation treatment are shown in Fig. 2. For Treatment 1, infection efflciency signiflcantly increased (P≤0.05) after UV-B radiation in comparison to controls, but there was no signiflcant effect (P>0.05) of UV-B radiation distance on infection efflciency. For Treatment 2, infection efflciency signiflcantly decreased (P≤0.05) after UV-B radiation in comparison to controls, and infection efflciency decreased as UV-B radiation distance decreased. Under Treatment 2,the infection efflciency under the UV-B radiation distance of 50 cm was signiflcantly different from that under the distance of 60 cm, but was not signiflcantly different(P>0.05) from that under the distance of 40 cm. For Treatment 3, infection efflciency gradually decreased with the UV-B radiation distance decreasing, but UV-B radiation only signiflcantly affected (P≤0.05) the infection efflciency under the distance of 40 cm. Treatments 4 and 5 showed the greatest differences compared to controls, with infection efflciency decreasing after UV-B radiation. In both of these treatments, infection efflciency decreased with decreasing UV-B radiation distance. These results show that infection efflciency was reduced by UV-B radiation under Treatments 2–5 in comparison to controls. Infection efflciency at the same distance across treatments was higher in Treatment 3 than in Treatment 2, and in Treatment 5 than in Treatment 4.

Fig. 2 The effects of different UV-B radiation distances (40, 50 and 60 cm) and non-UV-B (CK) on infection efflciency of wheat stripe rust under different UV-B radiation treatments. Treatment 1,pre-inoculation UV-B radiation; Treatment 2, post-inoculation UV-B radiation; Treatment 3, UV-B radiation both pre- and post-inoculation; Treatment 4, untreated seedlings inoculated with Puccinia striiformis f. sp. tritici (Pst) urediospores from leaves treated by UV-B radiation post-inoculation; Treatment 5, untreated seedlings inoculated with Pst urediospores from leaves treated by UV-B radiation both pre- and post-inoculation.Letters indicate signiflcance groupings at the 0.05 level. Data are mean±SD (n=3).
3.3. Effects of UV-B radiation on sporulation quantity
The effects of different UV-B radiation distances on sporulation quantity under each UV-B radiation treatment are shown in Table 1 and Fig. 3. In Treatment 1, the total sporulation quantity across the entire sporulation period under different distances increased with the decrease of UV-B radiation distance. Compared to the control, the total sporulation quantity under the distance of 60 cm deceased,but there was no signiflcant difference (P>0.05) between them. The total sporulation quantity under the distance of 50 or 40 cm signiflcantly increased (P≤0.05) in comparison to that under the distance of 60 cm and the control. In Treatment 2, UV-B radiation decreased sporulation quantity in comparison to controls, and the sporulation quantity on the same day post inoculation gradually decreased with decreasing radiation distance. Signiflcant differences were found among the total sporulation quantities under different UV-B radiation distances and the control. In Treatment 3,UV-B radiation decreased sporulation quantity, and the sporulation quantity on the same day post inoculation gradually decreased as radiation distance decreased. The total sporulation quantity under each radiation distance was signiflcantly lower than the control (P≤0.05). In Treatment 4,sporulation quantity on individual leaves was lower in comparison to controls, and decreased with decreasing UV-B radiation distance. The total sporulation quantities under the distances of 60, 50 and 40 cm gradually decreased with decreasing distance in comparison to the control, and there were signiflcant differences among them. In Treatment 5,sporulation quantity on individual leaves was lower than controls, and decreased with decreasing UV-B radiation distance. The total sporulation quantity under the distance of 60, 50 or 40 cm signiflcantly differed (P≤0.05) from the control. These results demonstrate that total sporulation quantity was reduced by UV-B radiation under Treatments 2–5 in comparison to controls. The total sporulation quantity under the same distance was more in Treatment 3 than in Treatment 2, and more in Treatment 5 than in Treatment 4.

Table 1 Total sporulation quantity under different UV-B radiation distances (40, 50 and 60 cm) in different UV-B radiation treatments
3.4. Effects of UV-B radiation on disease index of wheat stripe rust
Disease indices of wheat stripe rust under each UV-B radiation treatment across different radiation distances are shown in Table 2. In Treatment 1, compared to the control,disease index for UV-B irradiated plants on the 5th day post symptom appearance decreased, and then increased on the 10th and 15th days. On the 20th day, disease index decreased under the distances of 60 and 50 cm, and increased under the distance of 40 cm, in comparison to the control. However, the disease index for UV-B irradiated plants under each distance did not signiflcantly differ(P>0.05) from the control. In Treatment 2, UV-B radiation signiflcantly decreased (P≤0.05) disease index, and disease index on the same day post symptom appearance gradually decreased with decreasing radiation distance. Signiflcant differences were found among the disease indices for UV-B irradiated plants under different distances on the 10th or 15th day. On the 20th day, the disease index for UV-B irradiated plants under the distance of 60 cm signiflcantly differed from that under the distance of 50 or 40 cm. In Treatment 3, disease index for UV-B irradiated plants signiflcantly decreased (P≤0.05) in comparison to controls. On the 10th or 20th day, disease index under the distance of 40 cm signiflcantly differed (P≤0.05) from that under the distance of 60 or 50 cm. On the 15th day, the disease index under the distance of 50 cm did not signiflcantly differ (P>0.05)from that under the distance of 60 or 40 cm, but there was a signiflcant difference between the disease indices under the distances of 60 and 40 cm. In Treatments 4 and 5,UV-B radiation signiflcantly decreased (P≤0.05) disease index, and the disease index on the same day post symptom appearance gradually decreased as the radiation distance decreased. For Treatment 4, on the 5th, 15th or 20th day,the disease index under the distance of 60 cm signiflcantly differed (P≤0.05) from that under the distance of 50 or 40 cm,but there was no signiflcant difference (P>0.05) between the disease indices under the distances of 50 and 40 cm; on the 10th day, there were signiflcant differences among the disease indices under different distances. For Treatment 5,on the 5th or 15th day, the disease index under the distance of 50 cm did not signiflcantly differ (P>0.05) from that under the distance of 60 or 40 cm, but there was a signiflcant difference between the disease indices under the distances of 60 and 40 cm; on the 10th or 20th day, there were no signiflcant differences (P>0.05) among the disease indices under different UV-B radiation distances. These results show that the disease index of wheat stripe rust was reduced by UV-B radiation under Treatments 2–5. The disease index under the same distance was more in Treatment 3 than that on the same day in Treatment 2. The disease index under the distance of 60 cm was less in Treatment 5 than that on the same day in Treatment 4. However, the disease index under the distance of 50 or 40 cm was more in Treatment 5 than that on the same day in Treatment 4.

Fig. 3 The effects of different UV-B radiation distances (40,50 and 60 cm) and non-UV-B (CK) on sporulation quantity of wheat stripe rust under various UV-B radiation treatments. A,Treatment 1 (pre-inoculation UV-B radiation). B, Treatment 2(post-inoculation UV-B radiation). C, Treatment 3 (UV-B radiation both pre- and post-inoculation). D, Treatment 4(untreated seedlings inoculated with Puccinia striiformis f. sp.tritici (Pst) urediospores from leaves treated by UV-B radiation post-inoculation). E, Treatment 5 (untreated seedlings inoculated with Pst urediospores from leaves treated by UV-B radiation both pre- and post-inoculation). Data are mean±SD(n=3).
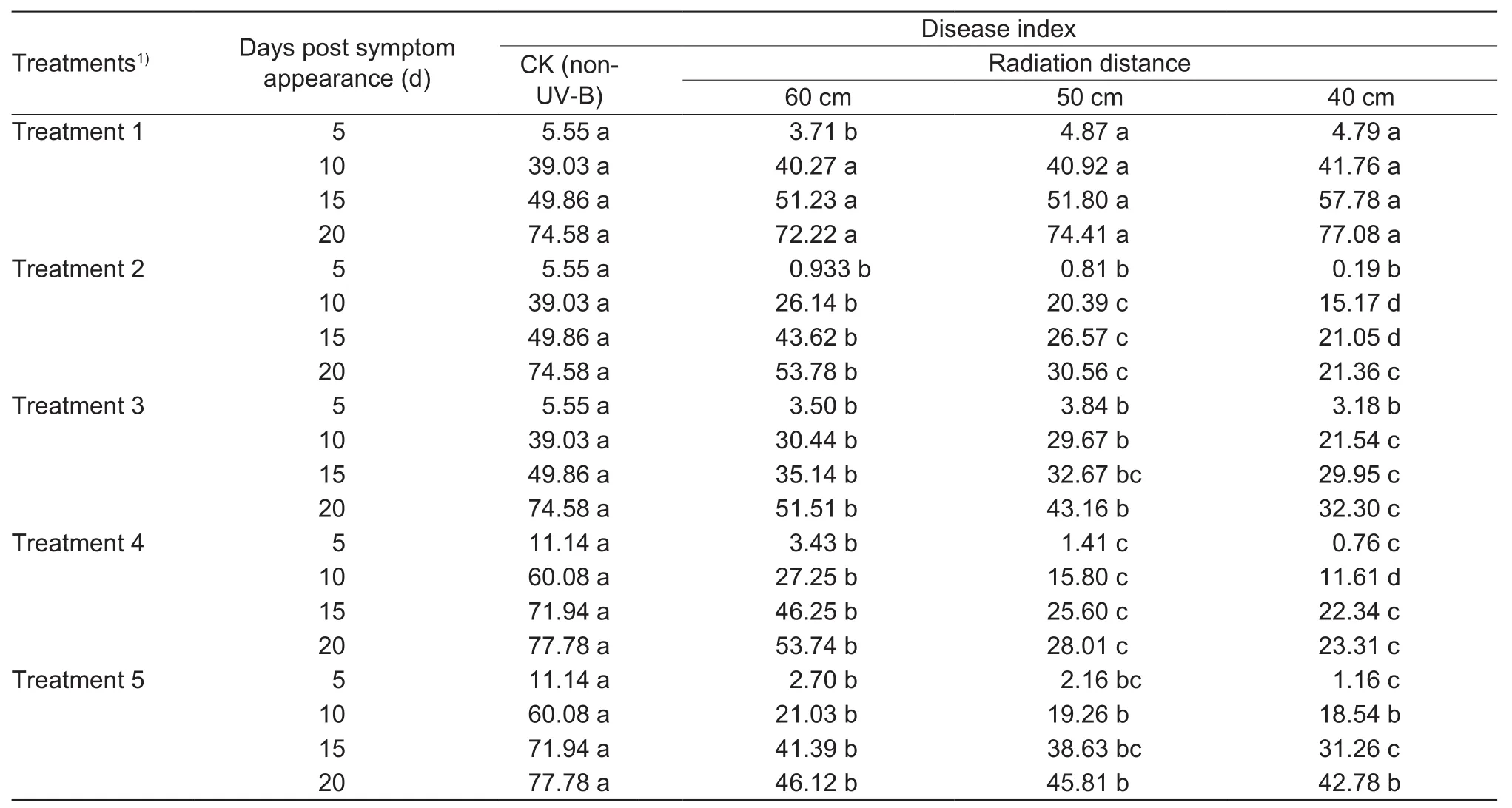
Table 2 Disease index of wheat stripe rust over time for various UV-B radiation treatments
4. Discussion
Caused by an airborne pathogen, wheat stripe rust is greatly influenced by environmental factors, of which UV-B radiation is important (Li and Zeng 2002). Many studies report effects of UV-B radiation on host plants or pathogens individually,but few studies look at the combined effects. In this study,UV-B radiation with different intensities was performed in different periods, and epidemiological components of wheat stripe rust were investigated. In particular, the effects of UV-B radiation pre- and post-inoculation were explored. The results obtained in this study show that the UV-B radiation both pre- and post-inoculation may affect the susceptibility of wheat and the pathogenicity ofPst. Pre-inoculation UV-B radiation may increase the susceptibility of wheat,and then aggravate disease occurrence. Post-inoculation UV-B radiation may reduce the pathogenicity ofPstand decrease disease occurrence. Furthermore, wheat stripe rust was reduced when UV-B radiation was carried out both pre- and post-inoculation. The effect of UV-B radiation on pathogenicity ofPstwas greater than on the susceptibility of wheat.
The exposure of host plants to UV-B radiation prior to inoculation may have different effects on plant diseases(Orthet al. 1990; Wargentet al. 2006; Liet al. 2008). For some diseases, the susceptibility of host plants increased when plants were irradiated with UV-B pre-inoculation (Orthet al. 1990; Liet al. 2008). It was also reported that disease occurrence may be aggravated by pre-inoculation radiation with UV-B (Wargentet al. 2006). In this present study, the susceptibility of wheat to stripe rust was increased by preinoculation UV-B radiation, and the incubation period was shortened and the infection efflciency, sporulation quantity and disease index increased. However, further studies are needed to determine the mechanism by which the preinoculation UV-B radiation aggravates wheat stripe rust.
Studies on the effects of post-inoculation UV-B radiation on wheat stripe rust have also been conducted. Liet al. (2008)showed that wheat stripe rust severity can be reduced by postinoculation UV-B radiation, and they suggested this flnding may be related to direct damage of UV-B radiation toPst.In contrast, Hanet al. (2011) reported that post-inoculation UV-B radiation had no effect on wheat stripe rust. In the present study, post-inoculation UV-B radiation prolonged the incubation period and reduced the infection efflciency,sporulation quantity and disease index, resulting in decreased wheat stripe rust occurrence. This may be due in part to damage ofPstby UV-B radiation. Although UV-B radiation affected both the host plant and the pathogen, the effect of UV-B radiation onPstwas greater than on the host plant.
The results of this study demonstrate that pre-inoculation UV-B radiation on wheat seedlings could reduce damage of post-inoculation UV-B radiation to the pathogenicity ofPstto a certain extent. Pre-inoculation UV-B radiation may cause stress in wheat plants, leading to increased activity of oxygen scavenging systems which reduce the level of reactive oxygen species (ROS) in order to reduce damage from UV-B radiation (Chen and Wang 1999). When UV-B radiation was also conducted post-inoculation, the ROS content may remain low because a large amount of active oxygen scavenging enzymes are already present due to the initial UV-B radiation treatment. However, active oxygen scavenging enzymes in wheat seedlings irradiated with UV-B only post-inoculation may be reduced or may not change (Zhang 2006), thus resulting in ROS accumulation.ROS can inhibit the germination of fungi (Levineet al. 1994).Therefore, damage toPstby UV-B radiation performed both pre- and post-inoculation may be less than that by the UV-B radiation performed only post-inoculation.
Changes in epidemiological components of wheat stripe rust caused by urediospores collected from UV-B irradiated wheat leaves were also investigated in this study. This subject has not been previously studied. When healthy wheat seedlings were inoculated using urediospores collected from wheat leaves irradiated with UV-B only post inoculation, the incubation period was prolonged and the infection efflciency, sporulation quantity, and disease index decreased. Post-inoculation UV-B radiation reduced infectivity ofPst, which may be related to reduction ofPstvitality due to UV-B radiation (Jinget al. 1993; Chenget al.2014). When healthy wheat seedlings were inoculated using the urediospores collected from the wheat leaves irradiated both pre-and post-inoculation,Pstpathogenicity was influenced and wheat stripe rust was reduced.Moreover, under the same UV-B radiation distance, total sporulation quantity was greater on leaves inoculated with urediospores from pre- and post-inoculation UV-B radiation plants compared to those with the post-inoculation treatment only. The effects on infection efflciency and disease index were similar to that on sporulation quantity. This may be due in part to low levels of ROS induced by pre-inoculation UV-B radiation (Chen and Wang 1999), thereby alleviating the damage of ROS toPst.
Plant growth in natural conditions will be influenced by a variety of factors. Hanet al. (2011) performed stress treatments onPst-inoculated wheat seedlings, and showed that UV-B radiation had no effect on wheat stripe rust, but also that the interaction of enhanced CO2concentration and UV-B radiation had no effect on the wheat disease in terms of epidemiological components including infection efflciency, incubation period, lesion expansion rate and AUDPC. To date, most studies related to the effects of UV-B radiation on plant diseases were conducted in controlled environments. However, artiflcial light differs from natural light, and factors such as temperature and humidity may also differ from those in natural environments. Therefore,further studies on the effects of different fleld environmental factors on plant diseases should be conducted in the future.Moreover, mechanisms of the effects of UV-B radiation on wheat stripe rust need further investigation.
5. Conclusion
The effects of the UV-B radiation intensity and timing on aspects of epidemiological components of wheat stripe rust including incubation period, infection efflciency, sporulation quantity and disease index, were investigated in this study.The results showed that pre-inoculation UV-B radiation aggravated wheat stripe rust occurrence, shortened the incubation period, and increased the infection efflciency,sporulation quantity and disease index. However, postinoculation UV-B radiation as well as both pre- and post-inoculation UV-B radiation reduced wheat stripe rust occurrence, prolonged the incubation period, and decreased the infection efflciency, sporulation quantity and disease index. Furthermore, the effects of UV-B radiation both pre- and post-inoculation on epidemiological components of wheat stripe rust were less than that of UV-B radiation post-inoculation only. Under the treatment where untreated healthy seedlings were inoculated withPsturediospores from the leaves treated by UV-B radiation only postinoculation, wheat stripe rust occurrence was reduced with a prolonged incubation period, and decreased infection efflciency, sporulation quantity and disease index. Under the treatment where untreated healthy seedlings were inoculated withPsturediospores from leaves treated by UV-B radiation both pre- and post-inoculation, wheat stripe rust occurrence was also reduced, and infection efflciency,sporulation quantity and disease index were also decreased,but incubation period did not change. Overall, the effects of all flve UV-B radiation treatments on the epidemiological components of wheat stripe rust increased with decreasing UV-B radiation distance (i.e., the increase of UV-B radiation intensity). These results provide a theoretical basis for further studies to monitor and predict wheat stripe rust as well as virulence variation ofPst.
Acknowledgements
This study was supported by the National Key Basic Research Program of China (2013CB127700) and the National Natural Science Foundation of China (31101393).
 Journal of Integrative Agriculture2018年12期
Journal of Integrative Agriculture2018年12期
- Journal of Integrative Agriculture的其它文章
- Analysis of three types of resistance gene analogs in PmU region from Triticum urartu
- Overexpression of the Suaeda salsa SsNHX1 gene confers enhanced salt and drought tolerance to transgenic Zea mays
- Effects of planting methods on yield and quality of different types of japonica rice in northern Jiangsu plain, China
- The role of rhizobacteria in rice plants: Growth and mitigation of toxicity
- Postponed and reduced basal nitrogen application improves nitrogen use efficiency and plant growth of winter wheat
- Effects of variety and chemical regulators on cold tolerance during maize germination