
Pikh,Pi2,Pi9,Pi5 4个稻瘟病抗性基因在贵州禾中的分布
2018-12-11 03:29马继琼孙一丁李进斌陈惠查焦爱霞谭金玉阮仁超许明辉
西南农业学报 2018年11期
马继琼,孙一丁,杨 奕,李进斌,陈惠查,焦爱霞,谭金玉,阮仁超,许明辉*
(1.云南省农业科学院生物技术与种质资源研究所/云南省农业生物技术重点实验室/农业部西南作物基因资源与种质创制重点实验室,云南昆明 650223;2.云南省农业科学院环境资源研究所,云南 昆明 650205;3.贵州省农业科学院农作物品种资源研究所,贵州贵阳 550006)
【研究意义】稻瘟病是全世界范围内水稻生产上最具毁灭性的病害之一。利用抗稻瘟病基因培育抗性品种是防治稻瘟病最经济、有效、环保的手段。广泛筛选鉴定抗稻瘟病基因资源是培育抗稻瘟病品种的基础。【前人研究进展】截至目前,已至少报道了69个抗稻瘟病位点共84个主效基因。其中,Pb1,Pia,Pib,Pid2,Pid3,Pik,Pikh,Pi54,Pik-p,Pish,Pit,Pita,Piz-t,Pi1,Pi2,Pi5,Pi9,Pi21,Pi25,Pi36,Pi37,Pi56,Pi63,PiCO39 等 24 个基因已被成功克隆[1],为利用分子标记检测这些基因提供了可能性。稻瘟病抗病基因Pikh是对稻瘟病菌具有广谱抗性的主效抗病基因,如云南省检测282个稻瘟病菌株的抗性已达81.7%[2],广东省163稻瘟病菌株的抗谱达 89.6%[3],四川、贵州、重庆、福建、吉林等地也具有较强的稻瘟病抗性[4-6],对病菌具有较宽的抗谱,在水稻抗病育种中有较大的利用价值。Pi9抗谱相当广,检测来自13个国家的43个稻瘟病菌株均表现出很高的抗性[7];在云南及黑龙江等地的水稻育种上具有极高价值[2,8],Abdullah 等建议推广种植携带Pi9基因的水稻品种[9]。Pi2的抗谱也较广,检测出对792个中国收集的稻瘟小种中绝大部分均表现抗性[10]。Pi5则在温室人工接种条件下对PO6-6等5个稻瘟病小种表现完全抗性[11]。
【本研究切入点】贵州禾类种质资源是指原产贵州东南部(特别是从江、黎平、榕江、锦屏等县)的一种地域性很强的特殊稻类生态型[12],是在独特的生态条件及当地少数民族(侗族)特殊的耕作制度下,经过长期的自然选择和人工选择所形成的一种稻种,主要分布于林间深谷,其山林郁闭严重,雾多、湿度大,阳光直射时间短,气候一日间寒热变化频繁,从而形成了与自然生态环境高度适应的具有品质优良和抗逆性强为主要特点的稻种生态类型。具有耐阴性、耐冷性和苗期抗旱性极强等特点,并具有广亲和基因[13]。【拟解决的关键问题】文章利用Pikh、Pi2、Pi9、Pi5分子标记对贵州禾进行分析,明确每个地方品种携带这4个抗性基因的状况,以评价贵州禾种质资源利用价值,也将有助于制定新的稻瘟病防治与抗病育种策略。
1 材料与方法
1.1 供试材料
供试材料均为贵州省地方水稻品种“贵州禾”,共82份,来自贵州省黔东南和黔南2个州相互接壤的5个县,其中从江县39份、黎平县28份、荔波县9份、雷山县4份、榕江县2份。均为科技部科技基础性工作专项“贵州农业生物资源调查(2012FY110200)”调查队2012-2015年调查中所收集的品种,目前民间仍有种植,感病对照为“丽江新团黑谷”。
1.2 实验方法
1.2.1 引物合成 PCR特异性引物参考文献[14-16],由生工生物工程(上海)股份有限公司合成,序列详情见表1。
1.2.2 基因组总DNA提取 采用CTAB法提取基因组DNA。
1.2.3 PCR反应 反应体系20 μl体系:Buffer 2 μl,dNTPs 1 μl,前后引物各 1 μl,Ex Taq 酶 0.4 μl,ddH2O 13.6 μl,DNA 模板 1 μl。反应程序:95 ℃,5 min;95℃,40 s;57℃,30 s;72℃,50 s;41个循环;72 ℃,10 min。
PCR扩增产物采用琼脂糖凝胶电泳,核酸染料染色,紫外灯下观察拍照。

表1 目标基因引物及标记片段大小Table 1 Sequences of primers for target gene and expected fragment length
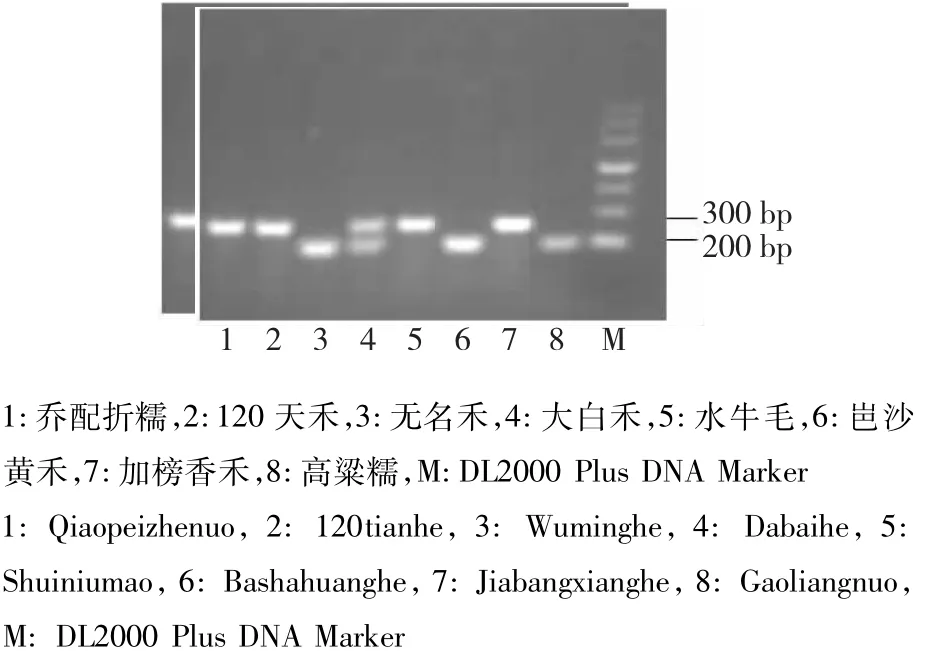
图1 部分品种Pi9基因分子标记检测Fig.1 Detection of Pi9 gene molecular markers in some varieties
1.2.4 叶稻瘟病田间抗性调查 2016-2017年材料种植于昆明市宜良县(海拔1600 m),属稻瘟病严重稻区。顺序排列,每个材料种植30株,感病对照为丽江新团黑谷,利用蒙古稻进行稻瘟病诱发。于7月上旬(封行期)进行叶稻瘟抗性调查,每个材料年度抗性级别有差异的以最高级别计。调查按《中华人民共和国国家标准GB/T15790-2009》品种抗性划分标准叶瘟抗性分级指标进行调查分级:0级:高抗(HR),无病;1级:抗(R),只有针尖大小的褐点或稍大褐点;2级:中抗(MR),圆形稍长的灰色小病斑,边缘褐色,直径1~2 mm;3级:中感1(MS1),典型纺锤型病斑,长1~2 cm,通常局限于两条主脉间,病斑面积2%以下;4级:中感2(MS2),典型病斑,病斑面积2.1% ~10%;5级:感(S1),典型病斑,病斑面积10.1% ~25%;6级:感(S2),典型病斑,病斑面积25.1% ~50%;7级:高感1(HS1),典型病斑,病斑面积50.1% ~75%;8级:高感2(HS2),叶片全枯死。
2 结果与分析
2.1 稻种资源稻瘟病抗性基因分子标记检测

图2 部分品种Pi2基因分子标记检测Fig.2 Detection of Pi2 gene molecular markers in some varieties

图3 部分品种Pikh基因分子标记检测Fig.3 Detection of Pikh gene molecular markers in some varieties
利用特异引物对82份贵州水稻材料和丽江新团黑谷、蒙古稻基因组DNA进行了扩增,每个基因均能扩增出预期大小的条带(图1~4),但有的品种不能扩增出相应的条带,很可能是该品种基因组引物结合位点发生了突变造成引物不匹配;Pi9有4个品种,Pikh和Pi5(图4中4号样)各有1个品种同时出现抗感条带,说明这些品种在此位点是杂合的。
由表2可以看出,通过长期的人工和自然选择,各稻瘟病抗性基因频率不一,Pi5(32.35%)、Pikh(30.86%)相对较高,而 Pi9(2.56%)和 Pi2(2.47%)频率较低。云南贵州地处同一高原,地方稻群体存在共同点,也存在差异。共同点是Pi9和Pi2在贵州禾和云南地方稻种频率均较低,而云南Pi5和Pikh频率分别为52.17%和65.22%,远高于贵州相应的基因频率[17]。
这4个基因可以组合成0(不携带所检测的任何基因)、Pikh、Pi5、Pi9、Pi5+Pikh、Pi2+Pi5、Pi9+Pi5等7种基因型,其比例分别是50%、21.95%、14.63%、1.22%、8.54%、2.44%、1.22%(表3),41个品种(基因型0)不携带所检测的任何1个基因,占50%,携带1个基因的品种(基因型Pikh、Pi5、Pi9)有 31 个(占 37.8%),2 个基因(Pi5+Pikh、Pi2+Pi5 和 Pi9+Pi5)的有 10 个(占 12.2%)。
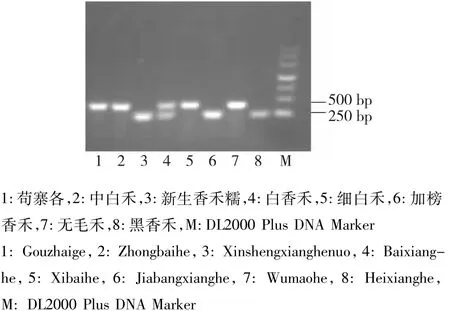
图4 部分品种Pi5基因分子标记检测Fig.4 Detection of Pi5 gene molecular markers in some varieties

表2 4个稻瘟病抗性基因频率Table 2 Frequencies of four rice blast resistance genes

表3 4个稻瘟病抗性基因组合的基因型及在各县的分布Table 3 Genotypes of four rice blast resistance genes and their distribution in each county
从表3还可以看出,从江、黎平、榕江、雷山4县材料是典型的贵州禾,出现 0、Pikh、Pi5、Pi9、Pi5+Pikh和Pi9+Pi5共6种基因型,而Pi2+Pi5基因型只出现在荔波县;从江县61.54%品种属于0型,0型为优势类型,黎平县的材料0型(46.43%)和Pikh(42.86%)同为优势类型,雷山县和榕江县由于供试品种少,选材存在一定的偶然性不能判断。荔波县虽与从江县接壤,供试品种少于从江县和黎平县,但基因型却达到了6种,且没有明显的优势类型,多于从江县(4种)和黎平县(3种),表明荔波县品种相对丰富。同时印证了“贵州禾”的SSR多样性程度较低的结果[18]。
2.2 稻种资源田间叶稻瘟抗性表现
在田间稻瘟病诱发试验中,诱发品种蒙古稻严重发病(8级),感病对照丽江新团黑谷田间表现感病(7级),表明大田发病充分。82个品种中,46个品种对叶瘟表现高抗(0~1级),占56.1%,其中黄鳝血(2013523309)、红米(2014522141)2个品种表现高抗(0级),表明贵州禾在云南宜良具有一定的抗性,这些品种分布在 0、Pikh、Pi5+Pikh、Pi5、Pi2+Pi5、Pi9+Pi5这6种基因型中(表4)。在品种数较多的0、Pikh、Pi5+Pikh和Pi5这4种基因型中,0型不携带所检测的任何基因,抗性变异最大,变幅1~6级,有18个(43.9%)表现感病(3~6级),平均抗性2.39级,而其他3种基因型变幅较小(1~3级),都有部分品种表现中感(3级),Pikh型有2个,Pi5+Pikh和Pi5型各有1个,3种基因型平均抗性分别为1.28、1.57 和1.17 级,抗性明显高于不携带所检测的任何基因的0型,表明田间抗性与所检测的Pi5、Pikh基因有一定的关联。Pikh和Pi5这2个基因在云南抗稻瘟病现代育种中起到了一定的作用,在现代育成品种中,70%品种携带Pikh,38.33%品种携带Pi5[17],携带Pikh和Pi5的贵州禾在云南宜良表现一定的抗病性,进一步证明这2个基因在云南水稻生产中的有效性。

表4 各基因型稻瘟病抗性比较Table 4 Comparison of rice blast resistance among 7 genotypes
在0型品种中,虽不携带所检测的任何一个抗性基因,但仍有15个品种在田间也表现出了高抗叶稻瘟,占36.59%,说明这些材料携带所检测基因之外的抗性基因;也有少部分携带1~2个抗性基因的品种表现感病,如高粱糯通过检测虽携带Pi9,但抗性为7级(高感)、占里糯禾携带Pi5+Pikh,抗性为3级(中感),说明品种稻瘟病抗性的表达较为复杂,分子标记鉴定有一定的局限性。
3 讨论
培育抗稻瘟病品种是防治稻瘟病最经济有效的途径之一,但稻瘟病抗性有非常强的小种专化性,且稻瘟病菌生理小种的多样性和易变性,极易导致抗性品种丧失抗性。抗稻瘟病育种和品种布局中,育种家必须要清楚亲本及后代品系携带的抗病基因种类和数量,指导品种的搭配和布局。然而传统的抗病基因分析方法,需要大量的生理小种接种鉴定、抗性遗传分析等工作,非常繁琐。在资源研究中,由于资源数量大,要逐一进行接种鉴定工作量十分巨大,携带的抗性基因不清很大程度上影响了资源的利用。随着水稻抗稻瘟病基因的克隆,抗性基因分子标记得到了快速的发展,大大提高抗病基因鉴定及育种选择的效率,已成为抗病育种的一种简单快捷的途径。抗稻瘟病基因分子标记可准确快速鉴定是否含有抗性基因,且具有操作简便、用时短的优点,可进行高通量的筛选,故近年来这一技术在国内水稻育种和资源鉴定中得到了广泛的应用[19-20]。
贵州地处云贵高原的东斜坡面,全境均属山区,属中国亚热带高原季风湿润气候。“天无三日晴”是对贵州多雨潮湿的生态条件最形象的描述,在这样复杂的生态条件影响下,稻瘟病菌的群体结构呈现多样性和复杂性。采用基因组重复序列PCR技术(rep-PCR)对贵州省的稻瘟病菌进行基因组指纹分析,黄培英等将200个供试菌株划分为19个遗传谱系[21],姜于兰等将154个供试菌株分为12个遗传谱系,6个组群21个小种[22]。在这样复杂的稻瘟病菌环境条件下,对57个主栽品种稻瘟病的田间抗性监测表明56个对稻瘟病表现出不同程度的感病性,多数品种不抗稻瘟病或田间抗性表现较差[23]。曾令祥对来自贵州省79个县(市)4 000余份地方品种接种鉴定表明抗稻瘟病种质在贵州9个行政区域均有分布[24]。
Pi9基因来源于小粒野生稻(Oryza minuta)[25],亚洲栽培稻(Oryza sativa)内的Pi9基因是种间基因交流的结果。从分析结果看“贵州禾”中Pi9频率较低,这与外来稻种基因交流少有关,仍保持其原始状态,值得进一步研究,建议贵州抗稻瘟病育种中应注意引入Pi9基因。
4 结论
贵州稻种资源以往的稻瘟病抗性研究均为田间和接种鉴定,本研究利用与 Pikh、Pi2、Pi9、Pi5稻瘟病抗性基因紧密连锁的分子标记对贵州禾携带的稻瘟病抗性基因进行分析,明确了各品种携带的基因种类和数量。Pikh、Pi5这2个基因在贵州禾中出现频率较高,得到了广泛保留和利用;田间抗性结果表明多数贵州禾对云南宜良的稻瘟病菌具有一定的抗性,携带1~2个抗性基因的品种一般均表现抗病,抗性与携带的基因有一定的关系。本研究结果也表明,部分不携带这4个基因的材料田间也表现高抗,表明这些材料还含有其他的抗性基因,所以在实践中可利用分子标记进行基因的初步筛选,再配以传统的接种鉴定和田间鉴定,才能准确地评价一个种质。
猜你喜欢
智慧农业导刊(2022年21期)2022-11-03
当代水产(2022年7期)2022-09-20
河北科技师范学院学报(2022年2期)2022-08-26
作物学报(2022年6期)2022-04-08
中国饲料(2021年17期)2021-11-02
空间科学学报(2021年1期)2021-05-22
农民致富之友(2020年15期)2020-05-25
农业知识(2019年7期)2019-04-26
中国森林病虫(2019年1期)2019-03-04
农民文摘(2019年7期)2019-01-14
