
普通小麦-大赖草易位系T6DL·7LrS的分子细胞遗传学鉴定
2018-12-07 01:04:42张雅莉王林生
生物工程学报 2018年11期
张雅莉,王林生
普通小麦-大赖草易位系T6DL·7LrS的分子细胞遗传学鉴定
张雅莉1,2,王林生1,2
1 河南科技大学 农学院,河南 洛阳 471023 2 洛阳市作物遗传改良与种质创新重点实验室,河南 洛阳 471023

张雅莉, 王林生. 普通小麦-大赖草易位系T6DL·7LrS的分子细胞遗传学鉴定. 生物工程学报, 2018, 34(11): 1823–1830.Zhang YL, Wang LS. Molecular and cytogenetic identification of Triticumaestivum-Leymus racemosus translocation lineT6DL·7LrS. Chin J Biotech, 2018, 34(11): 1823–1830.
大赖草高抗小麦赤霉病, 将大赖草抗性基因导入普通小麦, 对创新小麦赤霉病抗性种质有重要意义。为了获得普通小麦-大赖草抗赤霉病易位系,采用12 00 R60Co-γ射线处理小麦-大赖草二体附加系 DA7Lr花粉, 授予已去雄的普通小麦中国春,对其后代(M1) 种子根尖细胞有丝分裂中期染色体进行GISH分析,获得了1株具有1条普通小麦-大赖草易位染色体的植株,让其自交,对自交后代中具有2条易位染色体植株的花粉母细胞减数分裂中期I进行观察,发现2条易位染色体形成了稳定的棒状二价体,表明该植株为纯合体。利用顺序GISH-双色FISH分析,结合小麦D组专化探针Oligo-pAs1-2和B组专化探针Oligo-pSc119.2-2,进一步鉴定出该普通小麦-大赖草易位系为T6DL·7LrS,该易位系的育成也为小麦赤霉病遗传改良提供了新种质。
普通小麦,大赖草,易位系,分子细胞遗传学
大赖草(,2=4x=28,NsNsXmXm) 又称巨大冰麦草、大穗滨麦草,是禾本科赖草属草本植物,主要分布于北半球寒温带,在我国主要分布于新疆维吾尔自治区[1]、内蒙古自治区[2]的沙地和沙丘上。大赖草作为小麦的近缘野生物种,具有抗寒、抗旱、耐盐碱、抗小麦三锈(秆锈、条锈、叶锈)、赤霉病、黄矮病及根腐病等生物和非生物胁迫[3-5],是一种优异的小麦育种基因资源。
Mujeeb-Kazi等[6]成功获取普通小麦和大赖草的杂交种,并证明其高抗赤霉病。南京农业大学农业部作物细胞遗传重点实验室从1983年开始将大赖草的有益基因导入普通小麦的研究,获得了一批异附加系,并从中筛选出了具有较高赤霉病抗性的普通小麦–大赖草异附加系AddLr.2”、AddLr.7”和AddLr.14”[7]。Qi等[8]证明了Lr.2和Lr.14分别属于小麦的第7和第5部分同源群。为了减少大赖草整条染色体给普通小麦带来的冗余基因,遗传育种工作者采用了电离辐射、杀配子和Ph基因等方法创造易位系,以减少不利基因对普通小麦农艺性状造成的不良影响。刘文轩等[9]利用电离辐射大赖草单体异附加系Lr.7而得到了易位系T02和T08。王林生等[10]通过60Co-γ射线处理小麦-大赖草二体附加系DA5Lr雌配子,获得了普通小麦-大赖草染色体相互易位系T7DS·5LrL/T5LrS·7DL;崔承齐等[11]通过电离辐射得到了T7BS·7Lr#1S和T2AS·2AL-7Lr#1S易位系。
本研究在此基础上,通过电离辐射异附加系DA7Lr的花粉创造普通小麦-大赖草易位系,利用荧光原位杂交、分子标记技术和赤霉病抗性鉴定等技术,对辐射后代进行鉴定,筛选出抗赤霉病的普通小麦–大赖草易位系,为小麦抗赤霉病育种提供新的基因资源。
1 材料与方法
1.1 试验材料
利用60Co-γ射线(剂量1 200 Rad,剂量率100 Rad/min) 照射普通小麦–大赖草二体异附加系DA7Lr开花期麦穗,辐射花粉与普通小麦中国春杂交,让其后代种子自交,鉴定出纯合易位系。抗赤霉病对照品种苏麦3号和感赤霉病品种中国春由河南科技大学农学院小麦遗传育种研究室引进保存。
荧光原位杂交所用的两种小麦专化寡核苷酸重复序列的探针Oligo-pAs1-2 (Tamra-5′-CATTT CATCCACATAGCATGTGCAAGAAATTTGAGAG GGTTACGGCAAAAACTGGAT-3′)和Oligo-pSc119.2-2 (6-FAM-5′-TTCCACGATTGACGATTCCG GGG GTGCGTTTACGTGTCCGTCGTC-3′) 由上海英骏生物科技有限公司合成,荧光素Fluorescein- l2-dUTP和CY3-dUTP也由该公司提供。
1.2 染色体制片
1.2.1 根尖细胞有丝分裂中期
将小麦种子放入垫有湿滤纸的培养皿内发根(23 ℃),待根长至1.0−2.0 cm时,剪取1−2条种子根在冰水中处理20−24 h。用3份无水乙醇∶1份冰醋酸固定液固定,4 ℃冰箱中保存3 d后于45%醋酸中进行根尖压片,相差显微镜下染色体观察,放入−70 ℃冰箱或液氮中冷冻揭去盖玻片,脱水备用。
1.2.2 花粉母细胞减数分裂中期Ⅰ
挑取处于减数分裂中期Ⅰ的花药,用3份无水乙醇∶1份冰醋酸固定液固定,按根尖细胞有丝分裂中期制片方法压片。
1.3 染色体C-分带和荧光原位杂交
参照Gill等[12]的方法进行染色体C-分带。荧光原位杂交参照Mukai等[13]的方法稍加修改进行。参照Zhang 等[14]的方法顺次原位杂交,首先以大赖草基因组DNA为探针进行基因组原位杂交,通过SPOT CCD (Charge coupled device) 获取FISH图像,然后将信号洗脱,再以红色荧光标记Oligo-pSc119.2-2和绿色荧光标记Oligo-pAs1-2为探针进行双色荧光原位杂交,用DAPI (4ʹ,6-diamidino-2-phenylindole,DAPI) 染色,用SPOT CCD (Charge coupled device) 获取图像。图像结果参照Tang等[15]的标准图谱进行分析。
1.4 分子标记鉴定
植物DNA提取参照Sharp[16]的SDS法。采用10 μL PCR反应体系,模板DNA 20 ng、 1.5 mmol/L MgCl2、1× 缓冲液、200 mmol/L dNTPs、终浓度各为0.2 μmol/L的左右引物、0.5 UDNA聚合酶。PCR反应程序为:94 ℃预变性3 min;94 ℃变性30 s,50−60 ℃退火40 s,72 ℃延伸50 s,34个循环;72 ℃延伸10 min,4 ℃保存。扩增产物经8%非变性聚丙烯酰胺凝胶电泳,硝酸银染色后照相分析。
1.5 赤霉病抗病性鉴定
抗性鉴定在河南科技大学开元校区农场进行,试验按照随机区组设计。采用王裕中等[17]的单花滴注法接种禾谷镰刀菌(菌种由河南科技大学林学院植物病理系徐建强博士提供),接种后早晚两次喷雾,保证禾谷镰刀菌的发病湿度。2015−2017年连续3年均使用编号为LHLY-2高强毒菌株分生孢子悬浮液。每小区接种10穗,接种21 d后,调查接种穗的发病小穗数和总小穗数,计算病小穗率。病小穗率(%)=(病小穗数/总小数)×100。调查结果采用Spss软件进行统计 分析。
2 结果与分析
2.1 易位系T6DL·7LrS的分子细胞学鉴定
采用剂量1 200 Rad60Co-γ射线(剂量率 100 Rad/min) 照射普通小麦-大赖草二体异附加系DA7Lr开花期麦穗,将辐射附加系的花粉授予已去雄普通小麦中国春,对其后代种子进行荧光原位杂交(FISH) 分析,发现一株编号为NGH-02单株具有1条普通小麦-大赖草易位染色体,从自交后代种子中得到了1株含有2条易位染色体的单株,根尖细胞有丝分裂中期染色体制片,相差显微镜下观察并计数,2=44。
以绿色荧光标记大赖草基因组DNA并作为探针,对NGH-02纯合单株根尖染色体制片进行基因组原位杂交。结果显示,用荧光显微镜于450−490 nm激发光波长下可以观察到易位染色体的大赖草片段有弥散的绿色荧光杂交信号,且大赖草染色体端部有很强的杂交信号,易位的大赖草染色体片段为7Lr整个短臂即7LrS,易位的小麦染色体片段为某小麦染色体的一个臂(图1A)。因此可以判定该易位染色体是发生 在大赖草7LrS与普通小麦染色体之间的整臂 易位。
为了进一步确定易位染色体中小麦染色体片段的来源,将以大赖草基因组DNA为探针进行基因组原位杂交的信号洗脱干净,再以红色荧光标记的小麦B组专化探针Oligo-pSc119.2-2和绿色荧光标记的小麦D组专化探针Oligo-pAs1-2进行双色荧光原位杂交。原位杂交结果显示,在易位小麦染色体臂上有4个明显的绿色Oligo-pAs1-2点状杂交信号,没有发现红色的Oligo-pSc119.2-2的杂交信号(图1B)。因此可以判断易位的小麦染色体片段应为D组染色体。参照Tang[15]的原位杂交图谱,与这一荧光杂交信号特征相似的小麦染色体有3D、4D、5D、6D和7D。3D的短臂上有4个与易位染色体相似的杂交信号,但其短臂顶端同时具有Oligo-pSc119.2-2的杂交信号,可排除3D染色体;小麦4D和5D染色体长臂在着丝粒处还有明显的Oligo-pAs1-2 2个点状杂交信号,它们的长臂上有6个点状杂交信号,故二者也排除。小麦7D染色体长臂上虽然有4个点状Oligo-pAs1-2杂交信号,但位于长臂的近端处相距较近。而易位染色体的4点Oligo-pAs1-2杂交信号,其中2点位于长臂近着丝粒处,另两点位于长臂近端部(图1B和图2),因此在小麦D染色体中,只有6D染色体符合此特点。
对其花粉母细胞减数分裂中期Ⅰ染色体行为进行观察分析,发现3条易位染色体正常配对,形成了稳定的棒状二价体(图3),因此,该易位系命名为T6DL·7LrS。
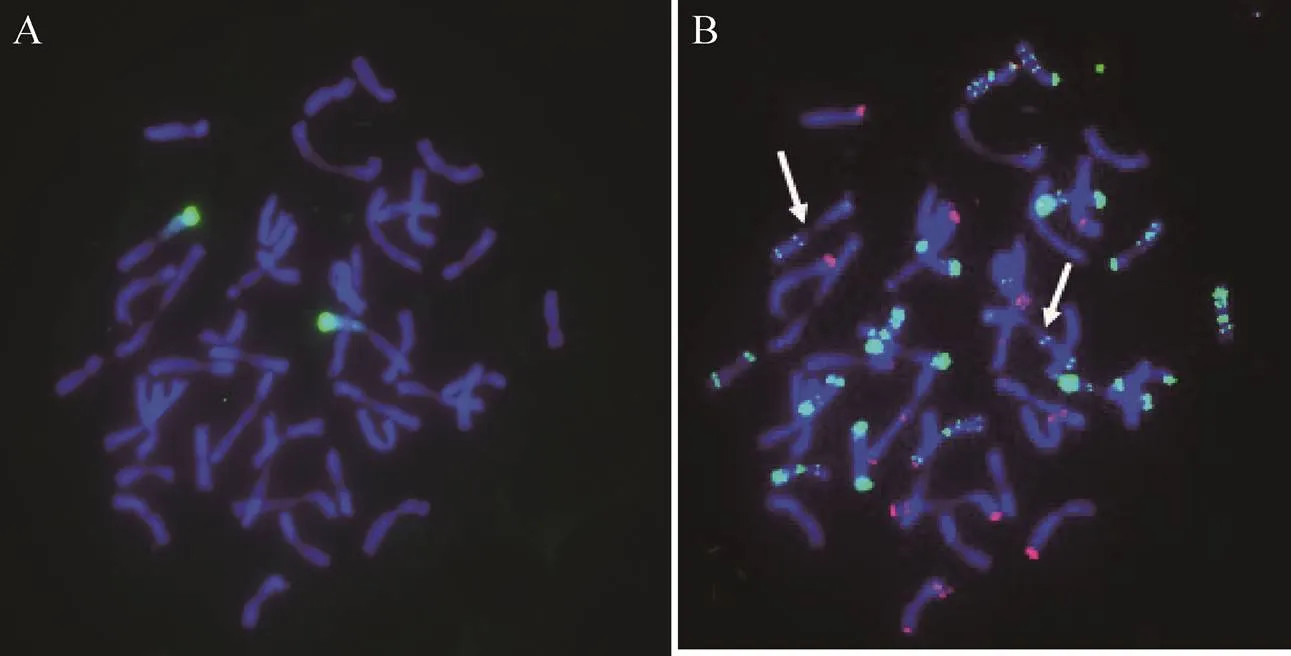
图1 易位系T6DL·7LrS荧光原位杂交(2n=44)

图2 易位染色体T6DL·7LrS荧光原位杂交
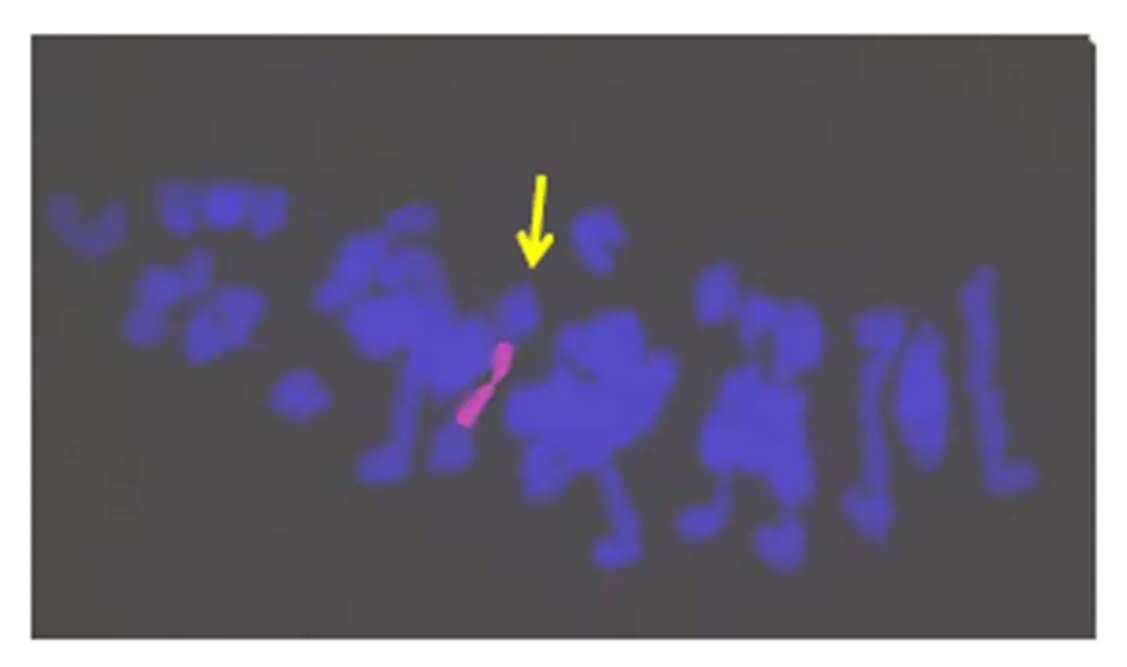
图3 易位系T6DL·7LrS花粉母细胞减数分裂中期Ⅰ染色体的荧光原位杂交
2.2 易位系的分子标记鉴定
根据小麦第7部分同源群不同区段的EST序列合成了81对EST-STS引物。经多态性分析,发现有3对引物在大赖草、DA7Lr和易位系T6DL·7LrS上有特异扩增,它们分别是BE591127、BQ168298和BE591737,均定位在第7同源群的7DS、7AS和7BS上,扩增出的3条特异条带,大小约1 000 bp、1 733 bp和398 bp (图4)。因 此,可以利用这3对特异引物追踪大赖草染色 体片段,鉴定含有该大赖草染色体片段的易位染色体。
2.3 易位系T6DL·7LrS的赤霉病抗性鉴定
经过连续3年的赤霉病接种鉴定,结果表明,易位系T6DL·7LrS的病小穗率显著低于感病亲本中国春和感病对照绵阳85-45,但稍高于抗病对照苏麦3号(表1和图5),表现出了较好的赤霉病抗性。
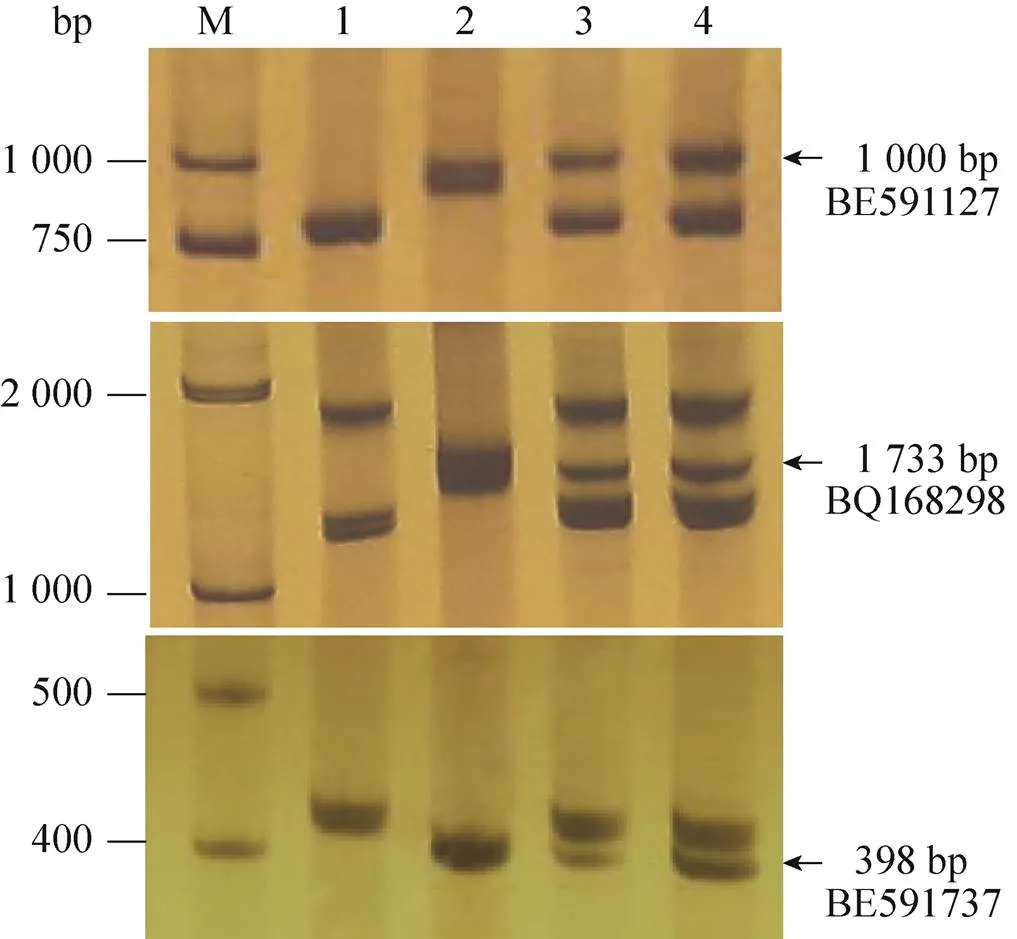
图4 三对引物的PCR扩增结果
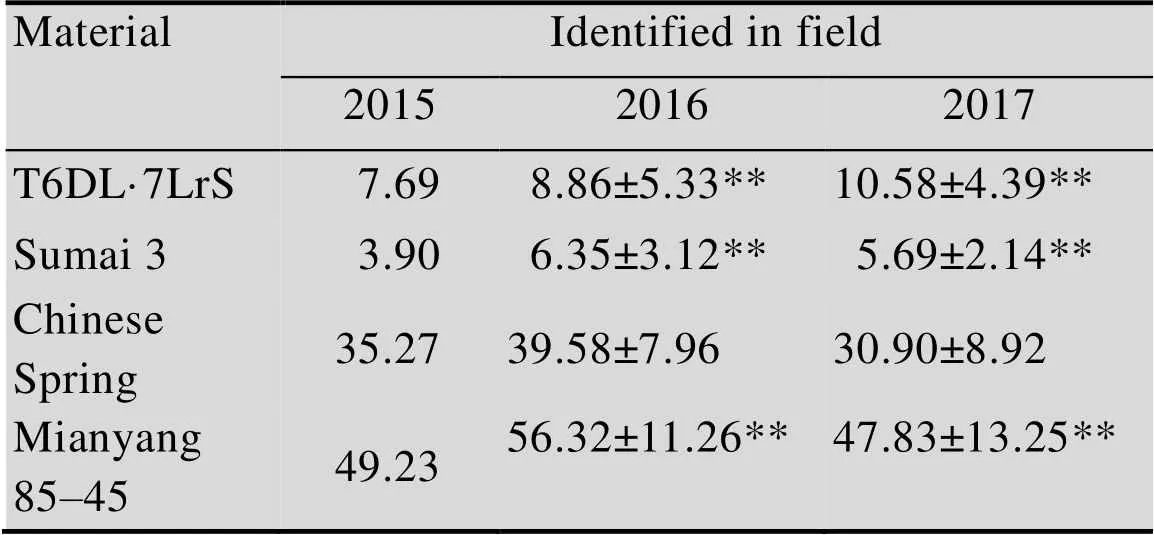
表1 T6DL·7LrS的赤霉病病小穗率
** indicate significant difference at<0.01 between the material and Chinese Spring.

图5 易位系T6DL·7LrS的赤霉病抗性鉴定
3 讨论
赤霉病是发生在温暖湿润麦区的一种灾难性病害,是一个世界性难题[18]。近几年不断往长江以北蔓延,并有明显扩展和加重趋势,仅河南省小麦赤霉病发生面积达100万公顷之多。究其原因是缺乏赤霉病抗性亲本,种质资源狭窄,仍大多局限于苏麦3号、Frontana等少数品种及其衍生系,很难有突破性进展。2017年国家农作物品种审定委员会审定通过的20个半冬性小麦品种及河南省审定通过的27个小麦品种全部高感赤霉病,致使我国小麦主产区存在巨大的安全隐患。因此,拓宽赤霉病育种遗传基础已成为育种工作者的当务之急。
大赖草作为普通小麦的近缘野生植物,高抗小麦赤霉病,早已引起研究者的高度重视。自20世纪80年代南京农业大学农业部作物细胞遗传重点实验室选育出高抗赤霉病的普通小麦大赖草异附加系以来,遗传育种工作者采用电离辐射、杀配子染色体等方法创造了一批普通小麦-大赖草易位系。刘文轩和杨宝军等[19-20]通过辐射Lr.2和Lr.7单体异附加系获得了易位系NAU618和NAU601。袁建华等[21]采用杀配子染色体诱导方法得到了3个易位系T1DS.Lr7L、T4AL.4AS-Lr7S和T1BL-Lr2S,这些易位系对赤霉病均表现较好的抗性,并在育种上进行了初步尝试。我们通过电离辐射小麦–大赖草二体附加系DA7Lr花粉创造出的普通小麦-大赖草易位系T6DL·7LrS,初步鉴定高抗小麦赤霉病,可以作为赤霉病抗性育种的中间材料。
尽管创造易位系的方法有多种,但电离辐射是最常用和最有效的方法之一。从Sears[22]采用电离辐射获得小麦-小伞山羊草易位系以来,已创造了许多具有外源有益基因的小麦新种质[23-28],为小麦育种发挥了重要作用。但由于电离辐射造成染色体断裂重接完全是随机的,产生的易位染色体补偿性往往较差,育种上直接利用还存在一定困难。本研究获得的易位系T6DL·7LrS,属于大赖草7Lr短臂与6D长臂的整臂易位,补偿性存在一定缺陷,同时也带入了大赖草的不利基因,造成结实性较差、植株较高、生育期延迟、千粒重较低等不良农艺性状,因此,可以在此易位系的基础上,创造携带大赖草有益基因的小片段 易位,尤其是中间插入易位已成为下一步研究的目标。
4 结论
本研究采用电离辐射的方法创造普通小麦大赖草易位系,并利用GISH-FISH双色荧光原位杂交技术,鉴定出了易位系T6DL·7LrS,并筛选出了3个在易位系T6DL·7LrS上有特异扩增的EST-STS多态性标记BE591127、BQ168298和BE591737,可有效追踪大赖草染色体片段。为赤霉病抗病育种提供了新思路。
[1] Dong YC, Su YZ, Zhong GY, et al. The exploration and preliminary study of(LAM.) tzvelev in Altai, Xinjiang Uighur Autonomous Region. Sci Agric Sin, 1985, 18(2): 54–56 (in Chinese).董玉琛, 孙雨珍, 钟干远, 等. 新疆阿勒泰地区大赖草的考察和初步研究. 中国农业科学, 1985, 18(2): 54–56.
[2] Yin GM, Zhao JH, Zhang YJ, et al. Adaption strategy aboutunder different grazing patterns. Anim Husband Feed Sci, 2013, 34(11): 33–36, 39 (in Chinese).殷国梅, 赵金花, 张英俊, 等. 大赖草对不同放牧方式的适应策略. 畜牧与饲料科学, 2013, 34(11): 33–36, 39.
[3] Dewey DR. The genomic system of classification as a guide to intergeneric hybridization with the perennial Triticeae//Gustafson JP, ed. Gene Manipulation in Plant Improvement. New York: Plenum Press, 1984, 209–279.
[4] McGuire PE, Dvôrák J. High salt-tolerance potential in wheatgrasses. Crop Sci, 1981, 21(5): 702–705.
[5] Mujeeb-Kazi A, Bekele GT, Mirand JL. Incorporation of alien genetic information frominto//Sakamoto S, ed. Proceedings of the 6th International Wheat Genet Symposium. Maruzen, Kyoto, Japan, 1983: 223–231.
[6] Mujeeb-Kazi A, Rodriguez R. An intergeneric hybrid ofL. X. J Hered, 1981, 72: 253–256.
[7] Chen PD, Wang ZT, Wang SL, et al. Transfer of useful germplasm fromLam. to common wheat. III. Development of addition lines with wheat scab resistance. Acta Genet Sin, 1995, 22(3): 206–210 (in Chinese).陈佩度, 王兆悌, 王苏玲, 等. 将大赖草种质转移给普通小麦的研究: III.抗赤霉病异附加系选育. 遗传学报, 1995, 22(3): 206–210.
[8] Qi LL, Wang SL, Chen PD, et al. Molecular cytogenetic analysis of Leymus racemosus chromosomes added to wheat. Theoret Appl Genet, 1997, 95(7): 1084–1091.
[9] Liu WX, Chen PD, Liu DJ. Development oftranslocation lines by irradiating adult plants at meiosis. Acta Bot Sin, 1999, 41(5): 463–467 (in Chinese). 刘文轩, 陈佩度, 刘大钧. 利用减数分裂期成株电离辐射选育小麦-大赖草易位系的研究. 植物学报, 1999, 41(5): 463–467.
[10] Wang LS, Chen PD, Wang XE. Molecular cytogenetic analysis ofreciprocal chromosomal translocation T7DS·5LrL/T5LrS·7DL. Chin Sci Bull, 2010, 55(11): 1026–1031.
[11] Cui CQ, Wang LS, Chen PD. Molecular and cytogenetic identification oftranslocation lines T7BS·7Lr#1S and T2AS·2AL-7Lr#1S. Acta Agron Sin, 2013, 39(2): 191–197 (in Chinese).崔承齐, 王林生, 陈佩度. 普通小麦–大赖草易位系T7BS•7Lr#1S和T2AS•2AL-7Lr#1S的分子细胞遗传学鉴定. 作物学报, 2013, 39(2): 191–197.
[12] Gill BS, Friebe B, Endo TR. Standard karyotype and nomenclature system for description of chromosome bands and structural aberrations in wheat (). Genome, 1991, 34(5): 830–839.
[13] Mukai Y, Nakahara Y, Yamamoto M. Simultaneous discrimination of the three genomes in hexaploid wheat by multicolor fluorescencehybridization using total genomic and highly repeated DNA probes. Genome, 1993, 36(3): 489–494
[14] Zhang P, Li W L, Friebe B, et al. Simultaneous painting of three genomes in hexaploid wheat by BACK-FISH. Genome, 2004, 47(5): 979–987.
[15] Tang ZX, Yang ZJ, Fu SL. Oligonucleotides replacing the roles of repetitive sequences pAs1, pSc119.2, pTa-535, pTa71, CCS1, and pAWRC.1 for FISH analysis. J Appl Genet, 2014, 55(3): 313–318.
[16] Sharp PJ, Chao S, Desai S, et al. The isolation characterization and application in the Triticeae of a set of wheat RFLP probes identifying each homoeologous chromosome arm. Theor Appl Genet, 1989, 78(3): 342–348.
[17] Wang YZ, Yang XN, Xiao QP. The improvement of identification technique of SCAB (Petch) resistance of wheat and the development of resistant sources. Sci Agric Sin, 1982, 15(5): 66–67 (in Chinese).王裕中, 杨新宁, 肖庆璞.小麦赤霉病抗性鉴定技术的改进及其抗源的开拓. 中国农业科学, 1982, 15(5): 66–67.
[18] Liu DJ. Breeding wheat for scab resistance-a worldwide hard nut to crack//Proceedings of International Conference on Wheat Genetics and Breeding-Perspectives of the 21st Century for Wheat Genetics and Breeding. Beijing: Agriculture Publisher, 2001: 4–7 (in Chinese).刘大钧. 小麦抗赤霉病育种——一个世界性难题//21世纪小麦遗传育种展望——小麦遗传育种国际学术讨论会文集. 北京: 农业出版社, 2001: 4–7.
[19] Liu WX, Chen PD, Liu DJ. Selection, breeding and identification of T01-atranslocation line. Acta Agron Sin, 2000, 26(3): 305–309 (in Chinese).刘文轩, 陈佩度, 刘大钧. 一个普通小麦_大赖草易位系T01的选育与鉴定. 作物学报, 2000, 26(3): 305–309.
[20] Yang BJ, Dou QW, Liu WX, et al. Development oftranslocation lines NAU601 and NAU618 and their test-cross analysis with double ditelosomic. Acta Genet Sin, 2002, 29(4): 350–354, 377 (in Chinese).杨宝军, 窦全文, 刘文轩, 等. 普通小麦-大赖草易位系NAU601和NAU618的选育及双端二体测交分析. 遗传学报, 2002, 29(4): 350–354, 377.
[21] Yuan JH, Chen PD, Liu DJ. Development oftranslocation lines using gametocidal chromosomes. Sci China Ser C: Life Sci, 2003, 46(5): 522–530.
[22] Sears ER. The transfer of leaf-rust resistance fromto wheat. Brookhaven Symp Biol, 1956, 9: 1–22.
[23] Sharma D, Knott DR. The transfer of leaf-rust resistance fromtoby irradiation. Genome, 1966, 8(1): 137–143.
[24] Sears ER, Gustafson JP. Use of radiation to transfer alien chromosome segments to wheat. Crop Sci, 1993, 33(5): 897–901.
[25] Friebe B, Jiang JM, Raupp WJ, et al. Characterization of wheat-alien translocations conferring resistance to diseases and pests: Current status. Euphytica, 1996, 91(1): 59–87.
[26] Luan Y, Wang XG, Liu WH, et al. Production and identification of wheat-6P translocation lines. Planta, 2010, 232(2): 501–510.
[27] Zhang J, Liu WP, Wu XH, et al. An intercalary translocation from6P chromosome into common wheat confers enhanced kernel number per spike. Planta, 2016, 244(4): 853–864.
[28] Song LQ, Lu YQ, Zhang JP, et al. Physical mapping ofchromosome 6P using deletion lines in common wheat background. Theor Appl Genet, 2016, 129(5): 1023–1034.
Molecular and cytogenetic identification of-translocation lineT6DL·7LrS
Yali Zhang1,2, and Linsheng Wang1,2
1 College of Agriculture, Henan University of Science and Technology,Luoyang 471023, Henan, China 2 Luoyang Key Laboratory of Crop Genetic Improvement and Germplasm Innovation,Luoyang 471023, Henan, China
had a high resistant capacity to wheat scab (Fusarum head blight). The transfer of scab resistant gene fromtois of great significance for broadening the germplasm of wheat resistance. To obtain-translocation line with scab resistance, we irradiated the pollen ofdisomic addition line DA7Lr by60Co-γ-rays 1 200 R (100 R/min) prior to pollinating to emasculationcv. Chinese Spring. One plant with one translocation chromosome was detected in the M1by GISH. The plant with one translocation chromosome was self-pollinated, and at meiotic metaphase I its progenies with two translocation chromosomes were analyzed for chromosome pairing behavior in their pollen mother cells (PMCs). One rod bivalent was observed at meiotic metaphase I, indicating that the plant with two translocation chromosomes was one translocation homozygote. Sequential GISH-FISH analysis, using Oligo-pAs1-2 and Oligo-pSc119.2-2 as probe, translocation line was confirmed as T6DL·7LrS. The translocation line had higher resistance to wheat scab and feasibility to be used as a new source in wheat breeding resistant to scab disease.
,, translocation line, molecular and cytogenetics
February 21, 2018;
April 23, 2018
National Natural Science Foundation of China (No. 31501301), Henan International Cooperation Program (No. 172102410052).
Linsheng Wang. Tel: +86-379-64282340; E-mail: 964965931@qq.com
10.13345/j.cjb.180066
国家自然科学基金 (No. 31501301),河南省国际合作项目 (No. 172102410052) 资助。
(本文责编 郝丽芳)
猜你喜欢
中国卫生标准管理(2022年21期)2023-01-03 02:36:34
华北农学报(2021年1期)2021-03-26 08:08:20
西南农业学报(2018年9期)2018-10-19 10:03:54
特产研究(2016年3期)2016-04-12 07:16:32
山东农业科学(2015年11期)2016-01-11 09:34:59
医学研究杂志(2015年2期)2015-06-10 06:45:00
湖北农业科学(2015年5期)2015-05-13 14:09:12
中国当代医药(2015年17期)2015-03-01 02:03:43
郑州大学学报(医学版)(2015年1期)2015-02-27 14:50:35
现代检验医学杂志(2014年4期)2014-02-02 02:44:41
