
冻融循环对五台山典型植被土壤微团聚体的影响
2018-12-05 08:52李杨波韩进斌郭勇强李变华申彦科
水土保持通报 2018年5期
李杨波, 刘 楠, 韩进斌, 郭勇强, 李变华, 申彦科
(1.太原理工大学 水利科学与工程学院, 山西 太原 030024; 2.山西省林业厅,山西省五台山树木园, 山西 忻州 035500; 3.中水北方勘测设计研究有限责任公司, 天津 300220)
中高纬度和高海拔地区随着温度的改变,晚秋到早春会出现土壤冻融现象,使其土壤团聚体组成改变[1-3]。高山林线附近区域是全球变暖的敏感指示体,又是生态交错带,在气候变化背景下,该区气候、植被覆盖情况、覆雪深度等都可能发生变化,进而影响该区土壤冻融循环频率和强度,导致土壤团聚体组成及分形维数等土壤结构指标改变[4-7],最终可能影响与之相关的生物地球化学过程及坡面稳定,特别是在秋冬、冬春两个季节交替时期。然而,季节性冻融对高山林线附近土壤结构的影响仍有待进一步研究。因此,本研究拟选择山西五台山林线附近典型植被草甸(CD)、华北落叶松(Larixprincipis-rupprechtii, HL)和云杉(Piceameyeri, YS),采用土柱野外原位培养法,探索冻融作用对其土壤的影响,以期为该区气候变化背景下的土壤生态研究提供基础数据。
1 材料与方法
1.1 研究区概况
五台山位于山西省东北部(38°27′—39°15′N,112°48′—113°55′E),山体横跨五台县、代县和繁峙县,属太行山系的北段,是太行山系支脉,位于黄土高原东部的森林草原交错地带,海河流域上游。其主峰北台(叶斗峰)海拔3 061.1 m,是华北最高峰。
五台山位于暖温带湿润半湿润气候区,山顶属于高寒气候。根据多年观测数据,中台气象站(海拔高度2 895.8 m)年均气温为-4.2 ℃,极端最低气温为-44.8 ℃,年均降雨量880.6 mm,是太行山气温最低、降水量最高,湿度和风力最大的地区[8-10]。五台山具有较完整的植被垂直带谱,植被带从山麓至山顶依次为:灌草丛及农田带—山地灌丛带—针、阔叶混交林带—针叶林带—亚高山灌丛带—亚高山、高山草甸带[11]。五台山植被对调节区域气候、涵养水源、水土保持、保持生物多样性等方面有着举足轻重的作用。
1.2 试验方案
(1) 样地选择。样地分草甸(CD)、华北落叶松(HL)和云杉(YS)3个典型植被样地,样地具体信息见表1。

表1 样地基本信息
注:CD为草甸样地;HL为华北落叶松样地;YS为云杉样地。下同。
(2) 采样点的布置。土柱:在每个群落内,分别在坡上、坡中、坡下设置一个20 m×20 m的固定样地,每个样地设置2个原位观测点,每个点设置7根土柱(PVC管,内径10 cm,长20 cm)。土柱垂直打入土壤中,尽量不破坏土壤的原状结构。
纽扣式温度记录仪(iButton DS1 992 L):在每个群落布置2个温度记录仪(埋在土壤10 cm深处),每小时自动监测土壤温度。根据温度记录仪记录的温度,得出图1。

图1 样地土壤温度记录
取样时间涵盖了整个变化周期,包括初冻期、深冻期、融化期,以及生长季。土柱7个采集时间点为:2015年9月26日,2015年10月30日,2015年12月20日,2016年3月15日,2016年5月20日,2016年7月26日,2016年9月26日。取样时间点表示一个时段的起始和结束,该时间点对应的微团聚体含量代表微团聚体含量经过一个时间段后的含量,如(20151030)表示生长季末期结束和初冻期起始,该时间点对应的微团聚体含量为经过生长季末期后最终含量。7个时间采集点将培养期分成6个时间段。详细情况见表2。

表2 研究取样时间每段时期的分类
1.3 样品检测
使用烘干法检测土壤含水量,用密度计法进行土壤微团聚体的测量。
1.4 分形维数的计算
根据杨培岭等[12]的分形维数公式:
D=3-lg(wi/w0)lg(di/dmax)
式中:D——土壤粒径结构分形维数;di——第i级粒径和第i+1粒径平均值(mm);dmax——最大粒级土粒平均直径(mm);wi——直径小于di的累积质量(g);w0——土壤样品总质量(g)。土壤样品分形维数计算方法为:首先求出土壤样品不同粒径(di)的1 g(di/dmax)和1 g(wi/wo)值,然后以前者为横坐标,后者为纵坐标做散点图,直线回归后求得斜率(K)最终用公式D=3-K求出土壤质量分形维数。
1.5 数据处理
采用Microsoft Office Excel 2013和SPSS 17.0统计分析软件进行数据分析处理。
2 结果与分析
2.1 季节性冻融对土壤微团聚体的影响
2.1.1 不同样地土壤微团聚体随时间变化特征 图2表明培养期内不同样地各粒径微团聚体含量变化。由图2得,在土壤温度变化剧烈的生长季末期、初冻期和融化期3个时期内,微团聚体含量发生显著变化,且不同样地微团聚体含量变化也不同。生长季末期,草旬样地0.25~0.05 mm,0.02~0.002,<0.002 mm微团聚体含量显著增加(p<0.05),2~0.25 mm,0.05~0.02 mm颗粒含量显著降低(p<0.05);华北落叶松样地2~0.25 mm颗粒含量显著增加(p<0.05),0.02~0.002 mm,<0.002 mm微团聚体含量显著降低(p<0.05);云杉样地2~0.25 mm,0.05~0.02 mm,<0.002 mm颗粒含量显著增加(p<0.05),0.25~0.05 mm,0.02~0.002 mm微团聚体含量显著降低(p<0.05)。可见在生长季末期,草甸样地较大粒径土壤颗粒有向更小粒径微团聚体转化的趋势,而华北落叶松和云杉样地正相反,虽然云杉样地0.05~0.02 mm,<0.002 mm微团聚体含量也显著增加,但其增长的绝对数量,远低于2~0.25 mm团聚体(图2)。

注:不同小写字母表示微团聚体含量在不同时段内差异性显著(p<0.05)。
图2样地微团聚体含量随时间变化
这与娄鑫等[13]的研究结果相似,温度变化剧烈,粒径较大的团聚体含量上升。但也有研究表明[14]土壤温度越稳定,越有利于土壤团聚体之间聚合,这可能是在其研究中,土壤温度(20 ℃左右)远高于冻融期,以及土壤含水量、有机碳等差异性所导致。而草甸样地微团聚体的变化,可能是由于在生长季末期植物出现凋零,其根系活性也随之下降,根对土壤颗粒的缠绕,特别是细根对土壤颗粒的缠绕减弱,致使较大粒径团聚体分散,土壤中粒径更小的微团聚体含量显著增加。随着土壤温度的下降,土壤动物、微生物活性的改变,也可能会造成草甸样地团聚体发生这种变化。对于云杉和华北落叶松样地,可能由于海拔低、枯落物较厚,再加上林木具有一定的郁闭度(表1),土壤外部环境可能较草甸样地稳定,所以这两个样地植物根系、土壤动物及土壤微生物的活性得以保持,且可能高于草甸样地,土壤颗粒间胶结物质,如生物分泌物等无明显减少,土壤颗粒间的作用力没有明显减弱,因此,华北落叶松和云杉两个样地土壤微团聚体可能会持续凝结,导致较大粒径团聚体含量增加(图2)。特别是对于海拔最低、枯落物层最厚的云杉样地(表1),其2~0.25 mm团聚体增加量明显高于华北落叶松样地(图2)。此外,土壤有机质含量及其随时间变化的差异性,也会造成研究区不同样地团聚体变化分异,彭新华等[15]和李鉴霖[16]认为土壤中有机质含量下降会使更小颗粒重新凝结,增大微团聚体粒径,但对于本研究区仍待进一步研究。
初冻期,草甸样地0.05~0.02 mm和华北落叶松样地0.25~0.05 mm微团聚体含量显著降低(p<0.05);草甸样地和云杉样地0.25~0.05 mm微团聚体含量均显著增加(p<0.05)。融化期,草甸样地和云杉样地0.02~0.002 mm微团聚体含量显著下降(p<0.05),草甸样地和华北落叶松样地0.05~0.02 mm微团聚体含量显著增加(p<0.05)。初冻期和融化期,季节性冻融对草甸样地土壤微团聚体的影响大于其余2个样地。可能是因为,草甸样地没有林木覆盖,且积雪较其余样地浅,基本没有枯落物覆盖,温度变幅较其余2个样地更剧烈,土壤水发生频繁相变,致使土壤微生物活性受到影响而降低。而对于华北落叶松和云杉样地,土壤表面枯落物和积雪作为保温层,使其土壤与外界环境隔绝,减少土壤温度变幅。同时,乔木层也作为保温层,一定程度上减少了下垫面热量向大气传输,致使这两个样地土壤生物所在环境也较为稳定,活性也相对较高。草甸样地土壤含水量与2~0.25 mm,0.05~0.02 mm和<0.002 mm微团聚体含量呈显著负相关关系(p<0.05),与0.25~0.05 mm微团聚含量呈显著正相关关系(p<0.05)。而对于华北落叶松和云杉样地,其土壤微团聚体含量与土壤含水量之间无显著相关性,这可能是因为其土壤温度较草甸样地稳定,另一方面华北落叶松和云杉样地的样地条件(表1),使得积雪之下一些苔藓和地衣等在寒冷时期能够继续存活,土壤中的生物活性可能高于草甸样地,最终也许是因为这些原因的干扰,导致这2个样地土壤含水量与其土壤微团聚体含量无显著相关。所以,在初冻期和融化期,草甸样地更多粒径团聚体含量变化显著。
深冻期样地土壤微团聚含量变化也相对稳定,只有草甸和云杉样地0.25~0.05 mm,云杉样地0.02~0.002 mm微团聚体含量显著下降(p<0.05)。深冻期初始阶段,云杉样地土壤温度在0 ℃上下波动,而在深冻期内草甸样地土壤温度虽然始终处于0 ℃以下,但也存在一定程度的波动(图1),一方面土壤中未冻水和冰晶可能因此出现相变,特别是云杉样地,其土壤温度波动阶段,可能使得一部分积雪融化进入土壤表层,增加了其土壤含水量,接下来土壤温度又进入0 ℃以下,这部分水的相变将会引起土壤颗粒的变化;另一方面土壤中的生物活性可能因温度变化而改变,因为在深冻期这个全年温度最低、环境最严酷的时期,土壤生物活性对温度变化的敏感性可能增强。因此,导致这两个样地土壤微团聚体含量显著变化(p<0.05,图2)。
生长季前期,草甸样地<0.002 mm微团聚体含量显著上升(p<0.05);华北落叶松样地0.02~0.002 mm微团聚体含量显著下降(p<0.05);云杉样地2~0.25 mm显著下降,0.25~0.05 mm,0.02~0.002 mm和<0.002 mm微团聚体含量显著上升(p<0.05)。生长季中期,草甸样地0.05~0.02 mm微团聚体含量显著下降(p<0.05),0.02~0.002 mm微团聚体含量显著上升(p<0.05);华北落叶松样地0.05~0.02 mm和<0.002 mm微团聚体含量显著下降(p<0.05);云杉样地0.05~0.02和<0.002 mm微团聚体含量显著下降(p<0.05),0.02~0.002 mm微团聚体含量显著上升(p<0.05)。生长季前、中期,土壤温度回升,土壤生物活性恢复。植物根系在土壤中穿插、网罗、固结、缠绕及其分泌物的胶黏作用,土壤动物和微生物的活动,使得土壤颗粒发生重新排列、组合[17-19]。同时,土壤有机质含量也随土壤生物活性变化而变化。这些都显著改变了各样地土壤微团聚体组成,也使该时期土壤微团聚体相比季节性冻融期间,出现了更加频繁而显著的变化。
2.1.2 不同样地微团聚体含量的差异性 各样地土壤微团聚体含量在培养期间差异性见表3。
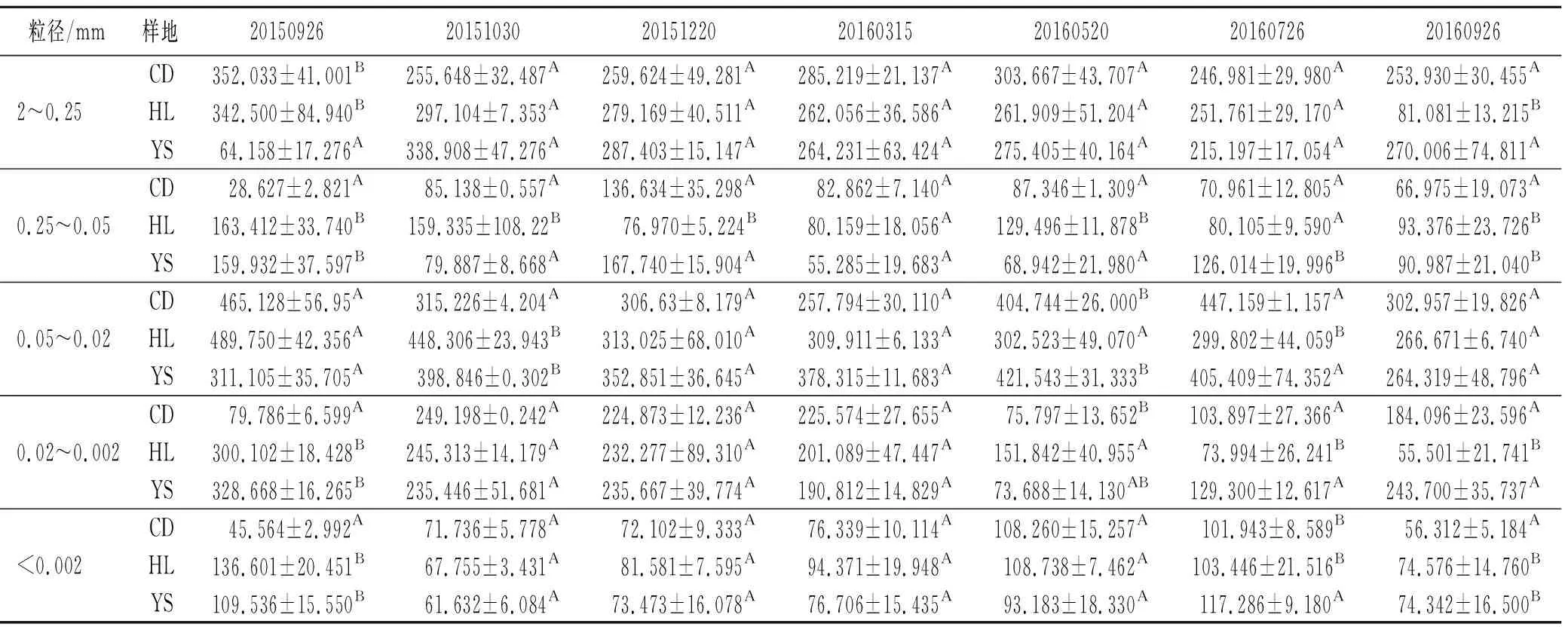
表3 研究区不同样地微团聚体含量比较
注:不同大写字母表示在同一时间点不同样地之间微团聚体含量差异显著(p<0.05)。
生长季,各级微团聚体含量在样地之间都有显著差异(p<0.05);初冻期和深冻期,除0.25~0.05 mm和0.05~0.02 mm微团聚体外(p<0.05),样地间土壤微团聚体含量无显著差异;融化期,仅有华北落叶松样地0.25~0.05 mm,0.05~0.02 mm和0.02~0.002 mm微团聚体含量与其余样地之间有显著差异(p<0.05)。可见,3个样地在生长季土壤微团聚体含量的差异明显多于季节性冻融期间,这可能是植被组成的不同,以及由此引起的土壤生物活性与有机质含量的差异,最终导致不同样地间土壤微团聚体含量和组成的显著差异。
2.2 冻融循环对土壤微团聚体分形维数的影响
由图3可知,在生长季末期,各样地土壤微团聚体质量分形维数减小,表明此时土壤的孔隙度增加,结构性变好,但是在初冻期、深冻期和融化期,土壤温度在0 ℃上下变化以及降低至0 ℃以下,土壤分形维数逐渐增加,表明在整个冻融期内样地土壤结性逐渐减弱。这与王恩姮等[20-21]的研究结果相似,然而,Lehrsch等[22-23]、王展等[24]却得出相反结果,土壤经历一次冻融循环后,土壤微团聚体分形维数降低,微团聚体稳定性和结构性变好。原因可能是本研究与王恩姮同样采用野外原位培养法,且试验样地植被相似;而Lehrsch、王展则采用室内冻融循环试验,样地为农田。再者本试验样地土壤含水量远高于王展试验样地土壤含水量,也可能是造成结果不一致的原因。

图3 研究区样地土壤分形维数变化趋势
生长季,植物根系的活性逐渐恢复,对土壤结构改善具有促进作用。而且土壤动物和微生物活性也得以恢复。首先,土壤动物活动频繁,土壤受到扰动发生破碎和移动[20];动物排泄物会增加,土壤有机物含量升高,从而使得土壤微团聚体稳定性增强,土壤结构性增强[25-26]。其次,微生物如细菌、真菌等自身的菌丝缠绕和它们自身释放的化学物质与土壤颗粒相结合[27],从而使得草甸和华北落叶松样地土壤微团聚体分形维数降低,土壤的结构性增强,然而在生长季初期,云杉样地却出现土壤微团聚体分形维数增加的趋势,祝忆伟等[28]也得出了相似的结论,其研究表明,样地土壤<0.002 mm和0.02~0.002 mm微团聚体含量与土壤分形维数呈正相关关系,而本研究云杉样地土壤分形维数同0.02~0.002 mm微团聚体也呈极显著正相关关系(p<0.01)。此外,由图2可知,生长季初期,云杉样地0.02~0.002 mm微团聚体含量显著增加(p<0.05),致使其土壤分形维数逐渐增加,也表明在此阶段,云杉样地土壤分形维数主要受0.02~0.002 mm微团聚体含量控制,与其余径级微团聚体含量无关。
总之,3个样地土壤微团聚体分形维数均具有周期性变化特征,即在经历了季节性冻融期的逐步增加之后,于生长季期间又逐渐降低,恢复至原有水平附近。从图3还可看出,云杉样地土壤微团聚体分形维数在季节性冻融期间总是小于其余两个样地,而华北落叶松样地土壤微团聚体分形维数虽然在深冻期超过了草甸样地,但是进入融化期又变得小于草甸样地,表明冻融作用对云杉样地土壤结构破坏性要小于草甸和华北落叶松样地,这主要与云杉样地优越的样地环境有关(表1)。进入生长季前期和中期,则是草甸样地的土壤微团聚体分形维数小于其余两个样地,可能是草甸样地在该时期大量的草本植物活性恢复,且高于其余2个样地,而乔木根系在表层土壤中有可能少于草本植物,因此根系对草甸表层土壤作用较强,优化了草甸土壤结构。在生长季末期则相反,草甸样地的土壤微团聚体分形维数大于其余两个样地,华北落叶松样地最小,可能因为郁闭度、枯落物厚度等原因,华北落叶松和云杉样地土壤温度得以较好的保持,不易随外界温度变化而剧烈变化,草甸样地则不然,因此,其草本植物根系活性下降较其余两个样地大,导致其土壤微团聚体分形维数较高,但这更能体现出草本植物根系对表层土壤结构的作用。然而,3个样地土壤微团聚体分形维数仅在生长季末期开始时存在显著差异(p<0.05),其余无显著差异。这与胡小兰[29]、甘凤玲等[30]的研究一致,可能是不同植被覆盖下的土壤中<0.002 mm和0.02~0.002 mm微团聚体含量差异显著(p<0.05)引起的(表3),且以上两者研究结果都表明土壤分形维数都与<0.002 mm和0.02~0.002 mm微团聚体含量呈显著的正相关关系(p<0.05),且0.02~0.002 mm与土壤分形维数的相关系数(0.962 6)大于<0.002 mm微团聚体(0.713 7)。
3个样地土壤含水量与土壤微团聚体分形维数之间存在显著负相关关系(p<0.05)。结合图3—4可知,研究区土壤微团聚体分形维数,即土壤结构性的周期性变化,与土壤含水量在季节性冻融循期的周期性变化密切相关。因为在季节性冻融期内,植物根系、土壤动物和微生物活性降低,对土壤颗粒的作用效果不显著,此时,含水量和温度可能是影响土壤分形维数的两个主要因素。而在生长季,土壤温度逐渐升高至合适水平,再加上恰当的土壤含水量,有利于植物、土壤动物和微生物活性恢复,因而它们对土壤颗粒作用效果较显著,土壤结构性明显增强,分形维数降低。

图4 研究区样地土壤含水量变化趋势
3 结 论
(1) 3个样地土壤微团聚体含量受到季节性冻融的影响发生改变,但比生长季土壤微团聚体的变化要少,且主要发生在初冻期和融化期。不过在深冻期,由于土壤温度波动和积雪保温作用,草甸样地和云杉样地土壤微团聚体含量也发生显著变化,但其变化仍然不及初冻期和融化期。
(2) 季节性冻融期间,样地植被不同造成的土壤微团聚体含量差异主要发生在融化期,而这一因素的影响在生长季期间表现的更加明显。
(3) 3个样地土壤微团聚体分形维数呈现一致变化趋势,都是在季节性冻融期间增加,进入生长季减少,体现出周期性变化规律和一定土壤结构恢复能力。
猜你喜欢
成都信息工程大学学报(2022年4期)2022-11-18
青海草业(2022年2期)2022-07-23
南方农业(2021年5期)2021-12-27
今日农业(2021年10期)2021-11-27
现代农村科技(2020年7期)2020-12-19
山东林业科技(2019年2期)2019-06-03
活力(2019年21期)2019-04-01
乡村科技(2019年14期)2019-02-21
现代营销(创富信息版)(2018年2期)2018-08-15
启迪与智慧·教育版(2018年12期)2018-02-26
