
玉米秸秆厌氧发酵过程中添加氮素对微生物群落和沼气产量的影响
2018-12-01 06:02赵一全马茹霞李家威梅自力王伟东
中国沼气 2018年5期
赵一全, 马茹霞, 李家威, 晏 磊, 罗 涛, 梅自力, 王伟东
(1.黑龙江八一农垦 大学生命科学技术学院, 黑龙江省寒区环境微生物与农业废弃物资源化利用重点实验室, 黑龙江 大庆 163319; 2.农业部沼气科学研究所, 农业部农村可再生能源开发利用重点实验室, 成都 610041)
我国作为农业大国秸秆资源量丰富,但利用率较低造成了资源浪费和环境污染,利用秸秆生产沼气是提高秸秆利用率和缓解环境污染的有效途径之一。沼气发酵是在多种微生物共同作用下完成的,温度[1],浓度[2],接种物,C/N[3-4],pH值[5]和搅拌[6]以及物料成分配比[7-8]等对厌氧发酵有重要影响。氮素是影响沼气发酵重要的营养元素之一,秸秆的碳氮比极高,这意味着以秸秆为原料的沼气发酵体系中氮素对于发酵过程的影响可能更大,优化发酵条件有利于微生物迅速繁殖,从而提高反应器性能和沼气产量。
根据秸秆的分子式计算秸秆完全液化能够提供厌氧发酵微生物所需的氮[9],但秸秆在实际发酵过程中很难全部被利用,且分解时间较长,微生物所需的氮含量不能在发酵各个阶段满足微生物的需求,尤其是在发酵初始阶段,影响了沼气工程的发酵效率和最终产量。余少杰等通过添加不同浓度的氯化铵来研究外加氮源对秸秆厌氧发酵的影响,研究结果[10]显示适量浓度的氮添加可以促进厌氧发酵体系中纤维素等物料的水解,而高浓度的氮素添加会抑制纤维素水解和甲烷生成。Piactek[11]等通过模型分析发现氮含量是影响水解和甲烷生成过程动力学的关键因素。Wagner[12]等研究了添加不同种类的氮源在厌氧发酵过程中对甲烷产量的影响,发现氮源中碳含量会影响甲烷的产率,在适宜的浓度范围内才能获得较高的甲烷产量。虽然原料中碳素转化为CH4和CO2是碳素间的循环,但在厌氧发酵中,微生物是将秸秆中的碳素转化为CH4的功能执行者。Wagner[13]等从多样性指数上分析了不同氮源对微生物多样性的影响,但未对功能菌株的丰度变化进行阐述。Alsouleman[14-15]等研究发现,高氨氮胁迫下厌氧反应体系发生适应性变化,主导菌群从拟杆菌转变为梭菌。随着宏基因组测序技术的快速发展,应用宏基因组技术来研究样本中直接参与碳和氮循环的功能微生物类群的总量和多样性是近年来环境微生物的研究热点之一,但对添加氮素后微生物群落多样性以及功能基因的变化以及如何影响发酵体系和甲烷产量的研究鲜有报道。因此,研究氮素添加对沼气发酵过程中微生物群落和关键功能基因的影响具有重要意义。
本研究解析玉米秸秆沼气发酵过程中添加氮素对料液中微生物多样性的影响,探究产甲烷关键功能基因丰度的变化,通过计算纤维素等物料的降解情况并结合产气量与产气周期,从微生物和功能基因的角度揭示氮素添加与秸秆沼气产量之间的关系,解析发酵过程中不同阶段细菌、古菌的多样性与功能特征,注释功能菌株的丰度的变化,进一步探究沼气发酵中关键因子的作用,为提高玉米秸秆沼气发酵产气效率提供数据支撑。
1 材料与方法
1.1 试验原料
试验用玉米秸秆取自黑龙江八一农垦大学实验实习基地,在10月份收获籽粒后,秸秆风干粉碎备用,接种污泥源于本实验室运行良好的厌氧发酵反应器,主要性质见表1,其中玉米风干秸秆的纤维素、半纤维素和木质素含量分别为30.52%,27.77%和10.99%。
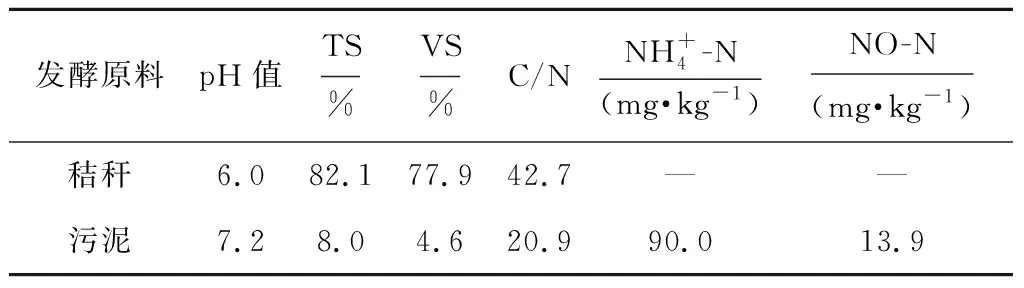
表1 发酵原料的主要性质
1.2 间歇式秸秆沼气发酵体系建立
试验装置选用25 L自制厌氧发酵反应器,上设有出气口和进料口,下设有出料口。出气口与15 L集气袋相连,用于收集气体并测定体积(见图1)。甲烷含量通过GA2000便携式沼气分析仪(Geotech Biogas Check)测定。发酵体积为20 L,接种污泥12 L,总TS设定为8%,启动时秸秆一次性进料,不足蒸馏水补齐。
根据前期单因素试验结果,选取添加有机氮尿素1 g·L-1作为处理,不加氮为对照,设置3个重复,控制发酵温度为35℃,反应时间60 d。随着发酵的进行,每10 d进行1次投料,物料添加量为总秸秆添加的1/6。
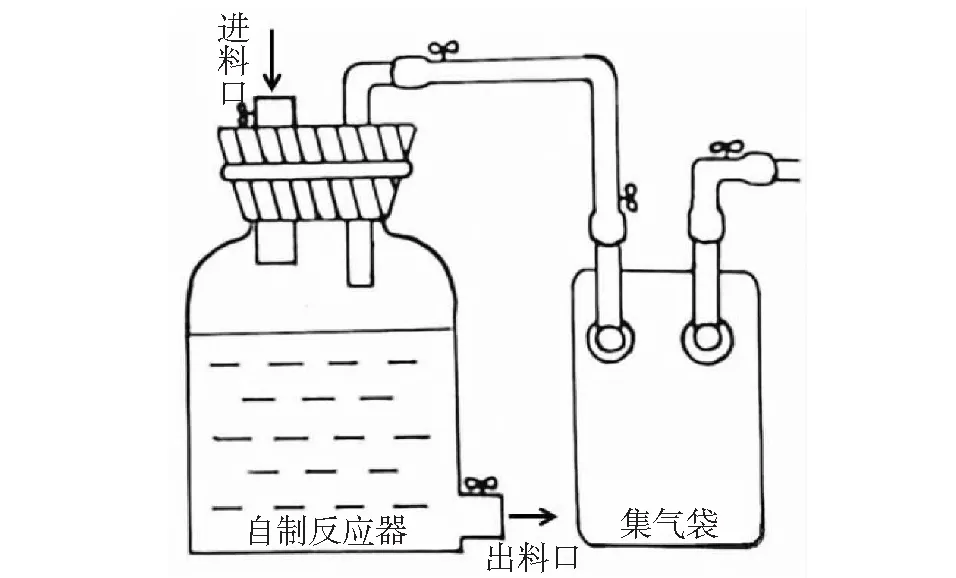
图1 发酵装置
1.3 试验方法
1.3.1 16SrRNA测序
在前期连续进料的沼气发酵试验中,对照和处理分别在发酵的第1,10,40天和60天进行取样,每个样品平行取3次,混合均匀后加入缓冲液(Extraction Buffer)-20℃保藏。利用改进的氯化苄法提取样品总DNA[16]。采用1%琼脂糖凝胶电泳检测,使用Nanodrop 2000c(Thermo)检测DNA的纯度和浓度。将合格的样品总DNA送至四川博贝特生物科技有限公司进行高通量测序。PCR扩增引物对应区域:细菌16S V4区引物515F(5′-GTGCCAGCMGCCGCGGTAA-3′)和909R(5′-CCCCGYCAATTCMTTTRAGT-3′)。古菌引物344F(5′-ACGGGGYGCAGCAGGCGCGA-3′)和806R(5′-GGACTACHVGGGTWTCTAAT-3′)。
1.3.2 统计分析
研究采用originpro 2017软件对试验数据进行处理和分析。
1.3.3 宏基因组学分析
在第一次产气高峰第10天进行取样,处理与对照样品平行取3次,混合均匀后加入缓冲液(Extraction Buffer),将样品送至上海派森诺公司进行DNA提取和宏基因组测序。
1.3.3.1 物种注释
将每个样本的Scaffolds/Scaftigs序列与NCBI-NT数据库中的细菌、古菌、真菌和病毒序列进行BLASTN相似性比对(E值设定为< 0.001)。由于每一条目标序列可能匹配多条参考序列,而这些匹配的参考序列又分属不同的分类单元,为使分析严谨可靠,同时又不丢失生物学意义,我们在MEGAN软件中采取“最低共同祖先(Lowest Common Ancestor,LCA)”算法[17],将参考序列分化为不同物种分枝前的最后一级共同分类,作为目标序列的物种分类注释信息。结合Scaffolds/Scaftigs序列在各样本中的丰度数据,获得各样本在所需分类等级上的相对丰度分布表。
1.3.3.2 功能注释
基因的功能注释是将非冗余蛋白序列集同常用的蛋白数据库进行比对,从而对各样本中的基因功能进行注释分析。KEGG注释是指通过将蛋白序列和KEGG代谢通路数据库进行比对,对宏基因组预测得到的基因根据代谢通路进行注释和分类。我们将上述非冗余蛋白序列集上传至KAAS(KEGG Auto maticAnnotation Server)进行功能注释,并对返回的注释结果进行汇总统计,获取各等级的注释结果及对应的丰度信息。
2 结果与分析
2.1 添加氮素对间歇式秸秆沼气发酵产气量的影响
在发酵的前28 d,处理的日产气量稳定且高于对照(见图2),在第10天达到第1个产气高峰,为17750 mL·d-1。处理和对照均在第25天达到产气高峰,分别为15077 mL·d-1和11707 mL·d-1。从第28天到第41天,对照恢复其产气潜力,日产气量高于处理,发挥产气优势。发酵前30 d进料对产气量的变化并不明显,从第40天开始,处理和对照在产气量上无显著性差异,产气趋势相似。产气量在每次进料后均上升,之后开始下降,到下一次进料产气量又出现了显著提升。测定发酵结束后纤维素和半纤维素降解率,处理达到33%和44%,显著高于对照的6%和28%。
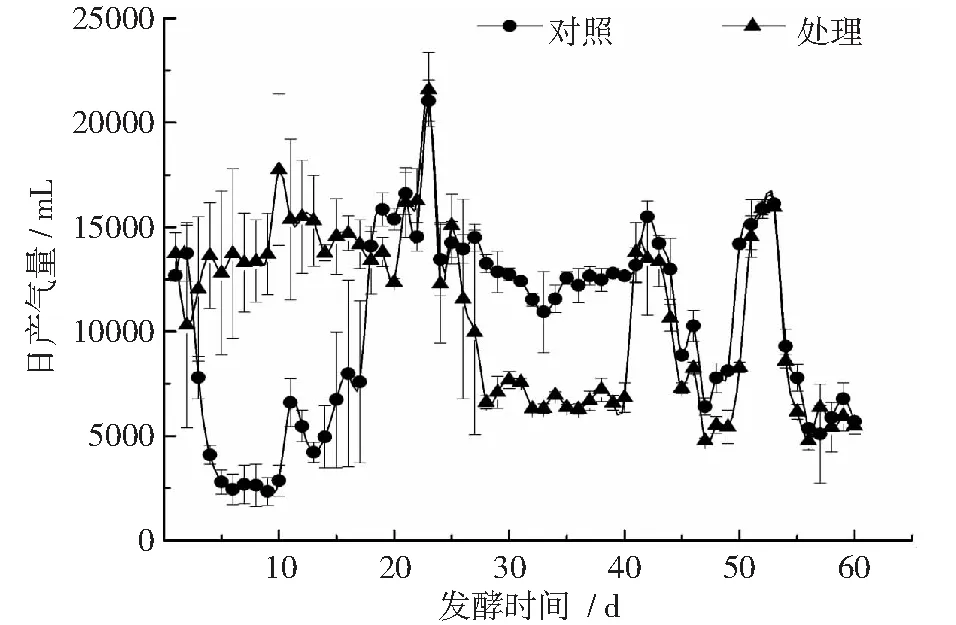
图2 加氮处理和对照的日产气量
日产甲烷量变化趋势与日产气量的变化趋势相似,笔者对发酵进行不同天数后的累积甲烷产量进行统计分析(见图3),发酵前10 d,20 d,30 d处理的甲烷累积产量分别是64943 mL,137048 mL,199459 mL,显著高于对照的14110 mL,50343 mL,123720 mL。发酵前40 d,处理的甲烷累积产量仍高于对照。发酵结束后,处理的甲烷总产量为308200 mL,仍然高于对照的284728 mL。处理在第10天达到第1个产甲烷高峰,为10880 mL·d-1。对照在第20天达到一个峰值,为6103 mL·d-1。处理和对照均在第23天达到日产甲烷产量的最高峰,分别为11388 mL·d-1和8164 mL·d-1。从第26天到第41天,对照日产甲烷量高于处理。从第41天到发酵结束,处理和对照的甲烷产量无显著差异。

图3 发酵不同时期甲烷累积产量
2.2 玉米秸秆沼气发酵过程中细菌和古菌组成多样性分析
2.2.1 16SrRNA测序结果
发酵料液中提取的7个样品总DNA条带清晰,杂质较少,无严重拖尾现象。Nanodrop检测出的DNA浓度在197.8~380.9 ng·L-1之间,OD260/280在1.90~2.07之间,均符合送样要求(浓度>10 ng·L-1;OD260/280在1.8~2.2之间)。对7个样品中的细菌和古菌的16SrRNA V4区序列进行高通量测序。对比Shannon指数可以看出(见表2),古菌的Shannon指数均低于细菌,说明细菌的微生物多样性要高于古菌的多样性。比较处理和对照中的细菌Alpha多样性指数,发酵第10天处理高于对照,随着发酵的进行,对照Shannon指数增加,说明其多样性有所增加,而处理的Shannon指数和Simpson指数均呈下降趋势,微生物多样性下降。从古菌的Alpha多样性指数变化规律可以看出,处理中古菌多样性的变化规律同处理中细菌变化一致。
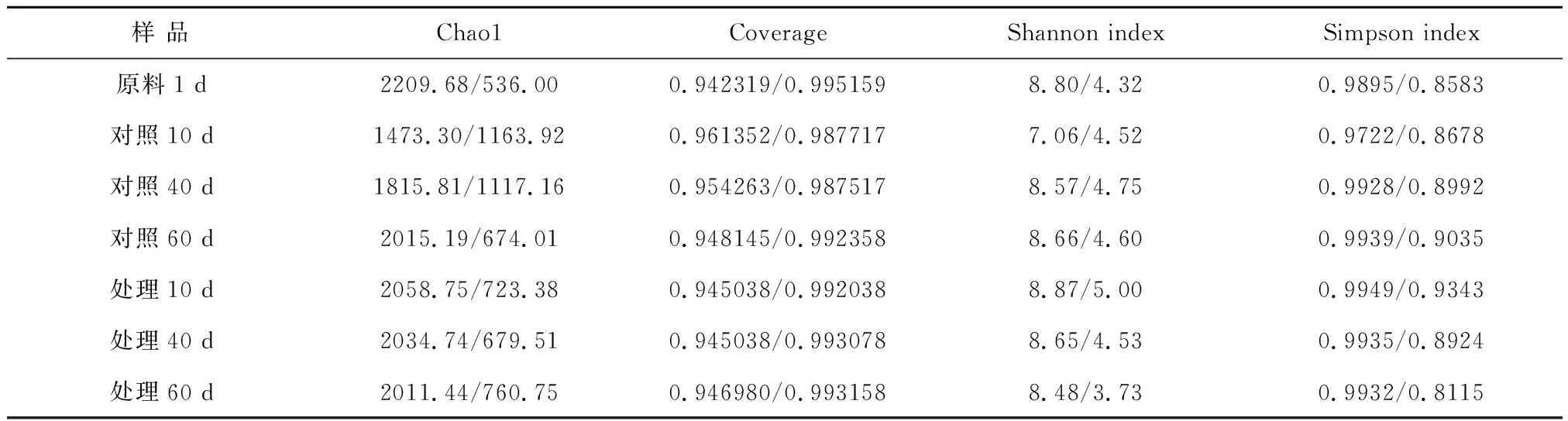
表2 细菌/古菌Alpha多样性指数
注:常见的Alpha多样性指数有Chao1丰富度指数,Shannon多样性指数,Simpson指数等。
2.2.2 发酵料液中细菌和古菌群落组成分析
将细菌的各菌群丰度占比1%以上的25类菌属作图(见图4),从属的分类水平进行分析,发酵原料中Clostridium占比最高。对照中,普氏菌属(Prevotella)、拟杆菌属(Bacteroides)、瘤胃球菌属(Ruminococcus)和Sphaerochaeta占比较高。处理中Ruminofilibacter,密螺旋体属(Treponema)和Sphaerochaeta占比较高。比较发酵各时期对照和处理均表现出不同的细菌菌群组成,二者优势菌群各不相同,菌群多样性存在差异。比较对照和处理发现,Sphaerochaeta菌属在发酵的各时期均存在。
古菌的各菌群丰度在属的分类水平上分析(见图5),在整个发酵过程中,甲烷鬃毛菌属(Methanosaeta)(36.7%~68.7%)一直占据最高比例。对照中占比较高的菌群包括Methanosaeta(45.8%~57.1%),Methanoculleus(5.6%~16.2%)。处理中占比较高的菌群包括Methanosaeta(36.7%~68.7%),Methanoculleus(7.2%~14.2%)和甲烷螺旋菌属(Methanospirillum)(6.6%~8.5%)。
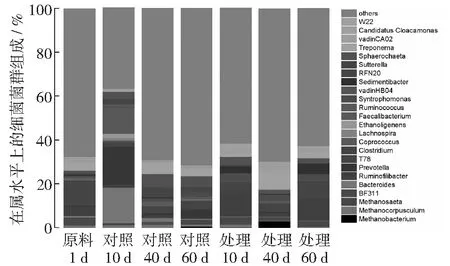
图4 发酵料液中细菌群落组成
在发酵第10天时,对照中Methanosaeta的丰度为57.1%,处理为36.7%,优势菌群Methanosaeta的数量随氮浓度的增加而减少。随着发酵的进行,处理中的Methanosaeta丰度逐渐增加,但对照中却在中期开始减少。说明Methanosaeta在氮素充足的环境中缺少竞争力,丰度下降。只有在环境中营养成分缺少的情况下Methanosaeta的竞争力才开始显现,成为环境中的优势菌属。Methanoculleus的丰度也受氮素的影响,其丰度随着氮浓度的升高而增加,Methanoculleus能在氮素充足的环境中快速成为优势菌属,随着发酵的持续和氮素的消耗,对照中Methanoculleus的丰度要高于处理。说明Methanoculleus可以充分利用秸秆分解后的营养物质成为环境中优势菌属,在环境中有较强的竞争力和适应能力。在发酵初期,氮素处理产气显著高于对照,可以看出在玉米秸秆厌氧发酵过程中,Methanoculleus对产气的贡献更大。随着发酵的进行,对照产气量持续提升,较处理产气率高,其中的甲烷杆菌属(Methanobacterium)丰度逐渐提升,对产气提升做出了较大贡献。
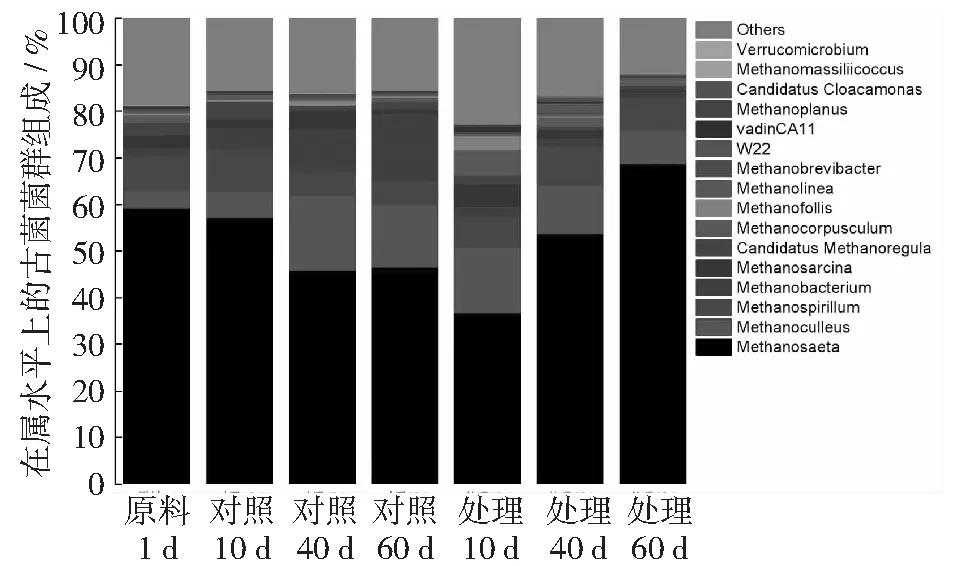
图5 发酵料液中古菌群落组成(属)
2.3 玉米秸秆沼气发酵体系宏基因组学研究
2.3.1 物种丰度差异分析
前期16SrRNA gene测序分析已经证明在发酵10 d处理和对照在丰度上存在差异,利用宏基因组数据可以进一步分析每个样本在各分类学水平的组成丰度分布,逐一比较每个分类单元在两个样本之间的丰度差异,并通过统计检验评价差异是否显著。处理与对照在门水平上有27个门具有显著差异,属水平上有475个属具有显著差异,在种水平上有777个种具有显著差异(见表3)。物种丰度的变化直接影响沼气发酵体系群落的功能性,尤其是功能菌种丰度的显著变化,通过以上数据可知,氮素添加能够显著影响发酵体系中微生物的丰度。

表3 丰度差异分析统计
注:表中第一列为两两比较的样本名;第二列至第四列分别对应两两比较中被认为具有显著差异的门、属和种的数量
2.3.2 功能基因差异分析
在功能基因水平上,我们分析了能量代谢中各功能基因的丰度(见图6),发现氮素处理中甲烷代谢(methane metabolism)的丰度高于处理,而氧化磷酸化等有关能量代谢的基因的丰度变化不明显。甲烷代谢基因是产甲烷的关键基因,氮素添加提升了甲烷代谢基因的丰度。

图6 能量代谢中各功能基因的丰度柱状图
3 讨论
Wagner[11]等研究了多种氮源添加对产气高峰期的影响,与本研究的氮素添加后产气高峰提前且一段时间产气量显著提升的研究结果一致,但没有对多周期发酵进行测定。我们在研究氮素添加的半连续发酵结果显示,处理在中后期产气能力低于对照。氮素添加虽然促进了纤维素的水解,为古菌的代谢提供了营养底物。但随发酵进行,处理中其它微生物所需的元素和营养底物被大量消耗,后期虽然氮素含量和对照持平,但根据元素守恒定律,其它必需元素含量会低于对照。而对照初期由于氮素含量和其他营养物质含量低,细菌代谢和纤维素水解受到限制,随着发酵的进行,纤维素水解产物等营养物质含量增加,细菌和古菌多样性逐渐提高,产气潜力开始显现。但整个发酵过程中处理的纤维素等物料的水解率显著高于対照,提供了更多的营养物质,处理的总甲烷产量仍高于対照。
在发酵起始阶段Clostridium丰度占比较高,这与Weimer[18]等发现Clostridium能够将纤维素和半纤维素等大分子水解成葡萄糖的结果一致。甲烷八叠球菌属(Methanosarcina)等产甲烷古菌的丰度提升可能和产酸菌群一样受氮素含量和底物浓度影响。产气高峰期Methanosarcina丰度增加,对产甲烷具有较大贡献,这与Bharathi[19]等的研究结论一致。Methanosaeta在处理和对照中丰度一直占据优势,Carr[20]等发现Methanosaeta有较强的产甲烷能力,结论与本研究结果相似。发酵体系中还注释到Sphaerochaeta,Abt B[21]等曾在白蚁的肠道中分离出该均属。Ritalaht[22]等曾在红杉河底层沉积物中分离出Sphaerochaeta.globosa,发现其能利用多种底物进行发酵产甲酸盐和乙酸盐。但在沼气发酵体系中,鲜有关于Sphaerochaeta的报道,其在玉米秸秆降解产甲烷环境中可能具有重要作用。
张彩杰[23]等在研究了C/N对细菌群落的影响时发现,碳氮比降低,细菌多样性提升,优势菌群丰度增高,这与本研究发现细菌的变化规律相同。本研究还分析了在产甲烷阶段具有重要作用的古菌群落多样性,发现古菌的群落多样性变化和细菌相似。对于食氢产甲烷菌和食乙酸产甲烷菌来说,丰富的营养底物可以被利用进行代谢,此时,产甲烷功能基因的丰度要比环境中微生物的多样性更为重要。Lin[24]等发现功能基因的多样性提升不利于代谢途径的有序性,并且可能带来更多的功能冗余,在探究有关能量代谢的基因丰度变化时发现,甲烷代谢相关基因丰度的增加和沼气产量呈正相关,与Qiang[25]等研究温度对沼气发酵体系影响的结果一致。虽然本研究没有探究功能基因的多样性与产气量的关系,但处理中的甲烷代谢功能基因表达显得更为集中。这说明氮素的添加和温度一样对甲烷代谢的功能基因具有调节作用。
本研究发现处理中古菌的多样性虽然提升了,但甲烷代谢的功能基因的丰度并没有降低,说明微生物多样性提升和功能基因的集中表达不存在矛盾。Shade[26]等认为微生物的多样性只是环境微生物中一个单纯的指标,我们同样认为多样性的高低不能直接说明一定的问题,复杂的代谢过程需要多种功能微生物共同参与,又或者环境中微生物多样性低,而重要的功能微生物丰度又占据绝对优势,或许这正是我们想要的,但自然环境往往不是这样的,这同样需要在具体的环境中分析。
4 结论
(1)以玉米秸秆为原料的厌氧沼气发酵过程中,利用16SrRNA gene和宏基因组测序技术分析发现,氮素添加能够提升厌氧发酵过程中细菌的多样性和丰度,细菌多样性的变化特别是具备纤维素降解功能菌株丰度的提升能够加速纤维素、半纤维素等物质的水解和代谢过程。氮素添加和纤维素等物质的加速分解有效的提升了反应器内古菌代谢所需的营养底物,提升了古菌的多样性和丰度,促进了产甲烷古菌的代谢和甲烷代谢基因丰度的提升。
(2)氮素添加通过提升微生物的丰度以及产甲烷功能基因的丰度,缩短了达到产气高峰所需的周期,并在一段时间内维持稳定的产气量和甲烷含量,最终在整个发酵周期内显著提升沼气产气量和TS甲烷产率。
猜你喜欢
环境卫生工程(2021年5期)2021-11-20
装备维修技术(2021年39期)2021-11-17
化工环保(2021年5期)2021-10-19
化学与生物工程(2021年8期)2021-08-26
兽医导刊(2021年15期)2021-01-10
环境卫生工程(2020年6期)2020-12-30
天津建设科技(2020年5期)2020-11-12
江苏理工学院学报(2020年2期)2020-10-23
小学科学(2020年2期)2020-06-15
小学科学(学生版)(2020年3期)2020-03-25
