
碱性土上GF677与毛桃植株生理指标及叶片组织结构对比研究
2018-11-30 06:46邱东昀钟小江宋海岩杨文渊江国良
西南农业学报 2018年10期
陈 栋,邱东昀,钟小江,祝 进,宋海岩,李 靖,杨文渊,江国良*
(1.四川省农业科学院园艺研究所/农业部西南地区园艺作物生物学与种质创制重点实验室,四川 成都 610066;2.乐山市市中区脆红李技术协会,四川 乐山 614000;3.宜宾县农业局果树站,四川 宜宾 644600;4.四川省园艺推广总站,四川 成都 610016)
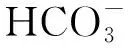
1 材料与方法
1.1 供试材料与试验地点
供试材料为抗性桃砧木GF677(扦插苗),以普通毛桃(实生苗)为对照。
试验地位于四川省简阳市周家乡南冲堰村四川省农业科学院园艺研究所晚熟桃示范基地(N30°30′50.97″,E104°26′35.21″),属亚热带湿润气候,年平均气温17 ℃,年降雨量900 mm。试验地块上茬作物为桃树,已种植20余年,园区土壤为侏罗系和白垩系紫色砂页岩发育而成的石灰性紫色土[8],土壤背景值:pH值8.44,有机质6.53 g/kg,全氮0.60 g/kg,全磷0.631 g/kg,全钾20.03 g/kg,阳离子交换量19.2 cmol(+)/kg,全铁63.3 g/kg,有效锌0.54 mg/kg,有效铁19.4 mg/kg,有效锰5.5 mg/kg,交换性钙81.0 cmol 1/2Ca2+/kg,交换性镁1.6 cmol 1/2 Mg2+/kg。2014年采取聚土起垄方式改土重建,株行距为1 m×4.5 m,栽植时GF677和毛桃均为1年生苗,本试验测定时间为2016年5-10月。
1.2 试验方法
选取生长势一致的GF677与毛桃实生苗,单株为1重复,每个处理设置5个重复。自5月5日起每隔1月左右从每株树上选取树冠各方位、无病虫害新梢中部成熟功能叶10片,拍照后放入液氮带回实验室进行生理生态指标测定及叶绿素合成前体含量测定,另取鲜叶2片放入FAA固定液中用于制作石蜡切片,2片放入2.5 %戊二醛中用于观察叶绿体显微结构。
(1)抗氧化酶系统:过氧化物酶(POD)活性采用愈创木酚法、超氧化物歧化酶(SOD)活性采用NBT还原比色法、过氧化氢酶(CAT)活性采用过氧化氢比色法。
(2)MDA含量:采用硫代巴比妥酸法。
(3)相对电导率:使用德国WTW 公司Cond315i电导率仪。
(4)不同时期枝条及根系可溶性蛋白含量测定:采用考马斯亮蓝G250显色法,参照李合生[9]。
(5)不同时期枝条及根系可溶性糖及可溶性淀粉含量测定:
可溶性糖采用蒽酮-浓硫酸比色法测定,可溶性淀粉采用高氯酸浸提,蒽酮-浓硫酸比色法,参照李合生[9]。
(6)不同时期根系高铁还原酶活性测定参照陈丽茜[10],方法略有改动。反应液组成:0.5 mM CaSO4,0.1 mM Mes (4-morpho-lineethanesulfonie acid),0.1 mM BPDS(4,7-dipheny1-1,10-phenanhroline-disufonic acid)和100 μM Fe-EDTA,用1M NaOH将pH调到5.5。用蒸馏水清洗处理后的根2~3次,置于0.5 mM CaSO4溶液中5 min。取鲜根置于10~20 mL试管中,加入20 mL反应液,在25 ℃黑暗条件下反应1 h,并每隔15 min振荡反应液一次。在535 nm波长下测量反应液吸光度,以不加根样的反应液为空白对照。将根取出用吸水纸吸干水分,称取对应根样的重量。最后,根据Fe2+浓度标准曲线,得到反应液中Fe2+的量,并由此计算出高铁还原酶的活性(以μmol·g-1FW·h-1计)。
(7)新鲜叶片制成石蜡切片,步骤如下:叶片组织固定于4 %多聚甲醛,常规脱水包埋切片,脱蜡至水份干后用番红固绿染液A染色2 h,之后用自来水冲洗;梯度酒精脱色各1~2 min;用番红固绿染液B染色1~2 min;无水乙醇脱水3~5 min;最后用二甲苯透明,中性树胶封片。使用图像测量软件Digimizer4.3.4观测叶片上、下表皮、栅栏组织和海绵组织。
2 结果与分析
2.1 叶片抗氧化酶系统比较
POD是以过氧化氢为电子受体催化底物氧化的酶,主要存在于细胞的过氧化物酶体中,以铁卟啉为辅基,可催化过氧化氢,氧化酚类和胺类化合物,具有消除过氧化氢和酚类、胺类毒性的双重作用。
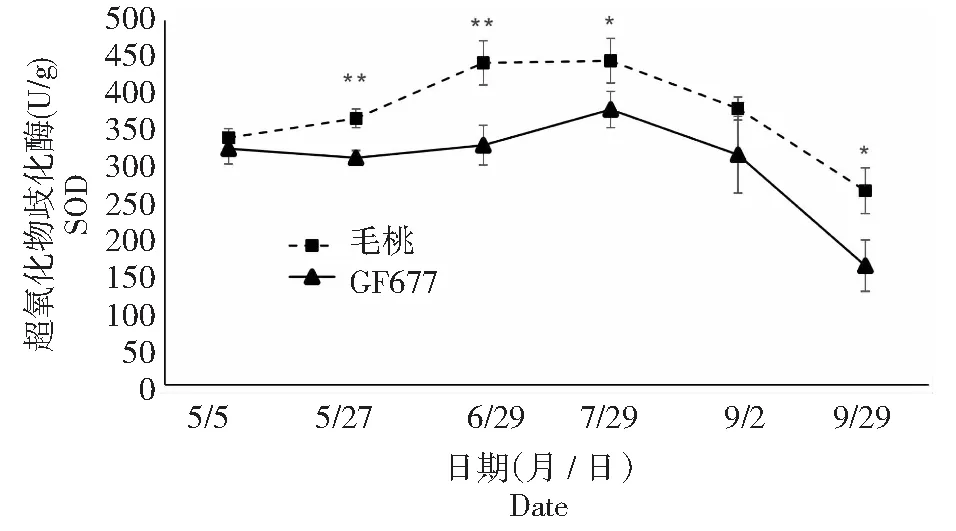
图中**表示该时期同一指标在P<1 %水平上差异显著,*表示该时期同一指标在P<5 %水平上差异显著,下同** in the picture indicates in P<1 % significant level in the same period and * in the picture indicates in P<5 % significant level in the same period,the same as below图1 碱性土上GF677和毛桃不同时期叶片SOD酶活性变化Fig.1 SOD activities of leaves of GF677 and wild peach on alkaline soil during different periods
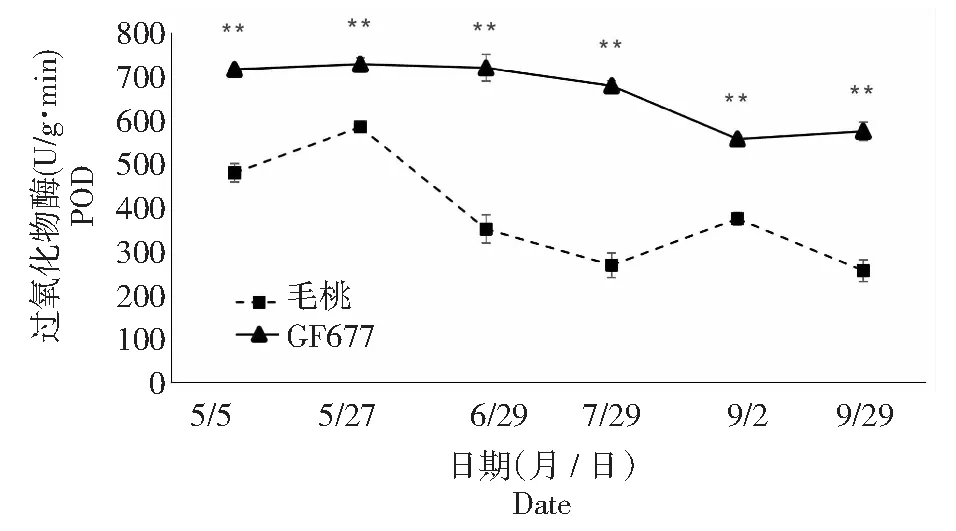
图2 碱性土上GF677和毛桃不同时期叶片POD酶活性变化Fig.2 POD activities of leaves of GF677 and wild peach on alkaline soil during different periods
由图2可知,碱性土上不同时期GF677叶片POD酶活性呈现先下降后略微上升的趋势,在5月5日至9月29日6个时期均极显著高于毛桃,甚至达到毛桃POD酶活性的2倍。
CAT是一种酶类清除剂,是以铁卟啉为辅基的结合酶。它可促使H2O2分解为分子氧和水,清除体内的过氧化氢,从而使细胞免于遭受H2O2的毒害,是生物防御体系的关键酶之一。由图3可知,GF677叶片因受到碱胁迫而导致CAT酶活性短期内略微升高,但随着碱胁迫时间的增加叶片CAT酶活性降低,最低值(120.19 U·g-1·min-1)出现在7月29日,随后由于GF677叶片适应碱胁迫后CAT活性开始上升。而毛桃叶片CAT酶活性受碱胁迫后呈现先下降后上升的趋势,最低值(72.57 U·g-1·min-1)出现在7月29日。GF677CAT酶活性在6月29日至9月29日4个时期均显著或极显著高于毛桃。
3种抗氧化酶(尤其是含有铁卟啉为辅基的POD和CAT)不同时期的变化,表明毛桃在受到碱胁迫后铁素吸收受阻,造成铁卟啉为辅基的POD和CAT酶活性显著或极显著低于GF677。GF677叶片在受到碱胁迫时体内抗氧化酶系统能够提高活性,减轻代谢产物的毒害,GF677表现出更强的抗性。
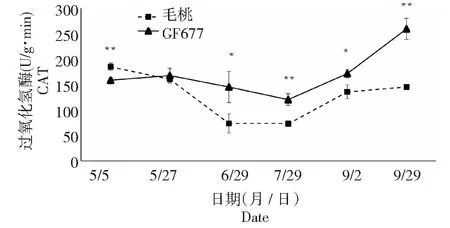
图3 碱性土上GF677和毛桃不同时期叶片CAT酶活性变化Fig.3 CAT activities of leaves of GF677 and wild peach on alkaline soil during different periods
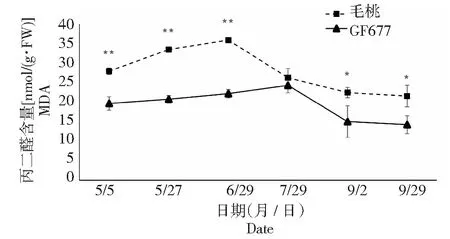
图4 碱性土上GF677和毛桃不同时期叶片丙二醛含量变化Fig.4 Malondialdehyde content of leaves of GF677 and wild peach on alkaline soil during different periods
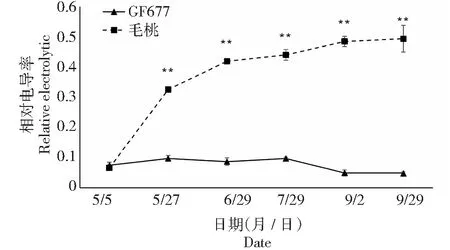
图5 碱性土上GF677和毛桃不同时期叶片相对电导率变化Fig.5 Relative electrolytic leakage of leaves of GF677 and wild peach on alkaline soil during different periods
2.2 叶片MDA含量比较
MDA是膜脂过氧化最重要的产物之一,它的产生还能加剧膜的损伤因此在植物衰老生理和抗性生理研究中MDA含量是一个常用指标,可通过MDA了解膜脂过氧化的程度,以间接测定膜系统受损程度以及植物的抗逆性。由图4可知,碱性土上不同时期GF677和毛桃叶片MDA含量均呈现先上升后下降的趋势,单毛桃叶片MDA含量在5月5日至6月29日以及9月2日至9月29日多个时期显著或极显著高于GF677。表明毛桃受碱胁迫后叶片膜系统受损程度大于GF677。
2.3 相对电导率比较
细胞膜透性的大小可间接的用组织的相对电导率衡量。组织相对电导率越高,说明细胞膜完整性遭到破坏的程度就越大。由图5可知,碱性土上不同时期GF677叶片相对电导率变化呈现不明显,而毛桃叶片电导率呈现先迅速上升后趋于平缓的趋势,最高值(0.49)出现在9月29日,随胁迫程度增加自5月27日起毛桃叶片相对电导率均极显著高于GF677叶片。再次表明毛桃在受到碱胁迫后叶肉细胞的细胞膜受到破坏,GF677表现出更强的抗性。
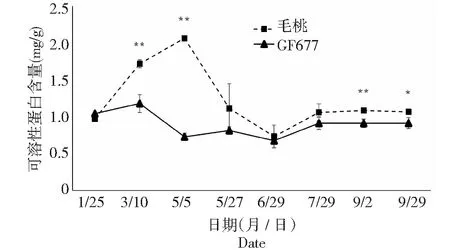
图6 碱性上GF677和毛桃不同时期枝条可溶性蛋白含量变化Fig.6 Soluble protein content of branches of GF677 and wild peach on alkaline soil during different periods
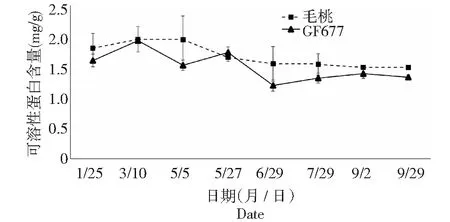
图7 碱性土上GF677和毛桃不同时期根系可溶性蛋白含量变化Fig.7 Soluble protein content of roots of GF677 and wild peach on alkaline soil during different periods
2.4 不同时期枝条及根系可溶性蛋白含量变化
可溶性蛋白是重要的渗透调节物质和营养物质,对细胞的生命物质及生物膜起到保护作用,因此经常用作筛选抗性的指标之一。由图6可知,碱性土上不同时期GF677枝条可溶性蛋白含量变化不大,而毛桃枝条可溶性蛋白含量呈现先上升后下降然后趋于平缓的趋势,最高峰(2.06 mg/g)出现在5月5日,其中在3月10日、5月5日、9月2日和9月29日多个时期毛桃枝条可溶性蛋白含量显著或极显著高于GF677。
由图7可知,碱性土上GF677与毛桃根系可溶性蛋白含量变化不大。表明通过枝条与根系可溶性蛋白含量难以鉴别桃树抗黄化能力。
2.5 不同时期枝条及根系可溶性糖及可溶性淀粉含量变化
可溶性糖是干旱胁迫诱导的小分子溶质之一,其种类主要包括葡萄糖、海藻糖、蔗糖等。这些可溶性糖类参与渗透调节,并可能在维持植物蛋白质稳定方面起到重要作用。由图8~11可知,碱性土上不同时期GF677与毛桃枝条可溶性糖变化趋势相近,但仍有不同:1月25日第1次测定时GF677枝条可溶性糖含量极显著高于毛桃,5月5日GF677枝条可溶性糖含量显著高于毛桃。碱性土上不同时期GF677和毛桃枝条可溶性淀粉变化趋势相近,但在5月5日、5月27日和7月29日至9月29日期间GF677枝条可溶性淀粉含量均极显著高于毛桃。
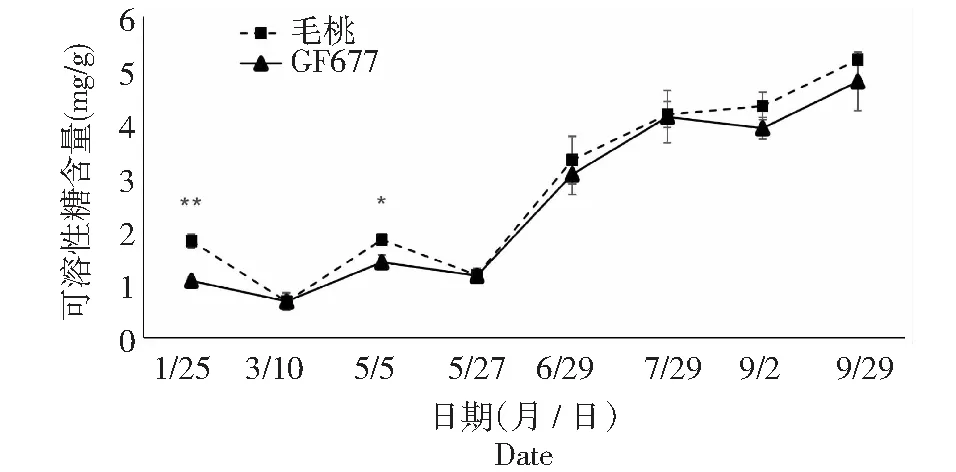
图8 碱性土上GF677和毛桃不同时期枝条可溶性糖含量变化Fig.8 Soluble sugar content of branches of GF677 and wild peach on alkaline soil during different periods
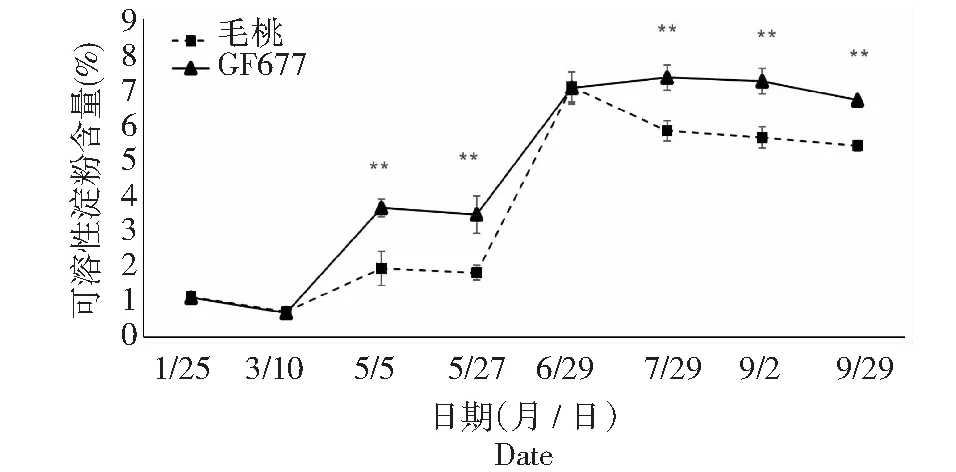
图9 碱性土上GF677和毛桃不同时期枝条可溶性淀粉含量变化Fig.9 Soluble starch content of branchs of GF677 and wild peach on alkaline soil during different periods
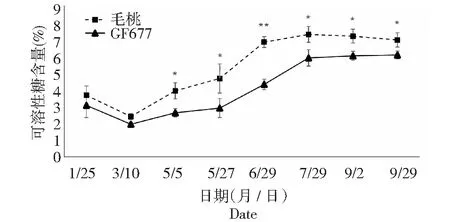
图10 碱性土上GF677和毛桃不同时期根系可溶性糖含量变化Fig.10 Soluble sugar content of roots of GF677 and wild peach on alkaline soil during different periods
碱性土上不同时期GF677和毛桃根系可溶性糖含量均呈现先下降后上升最后趋于平缓的趋势,且毛桃根系可溶性糖含量一直高于GF677,5月5日至9月29日期间毛桃根系可溶性糖含量均显著或极显著高于GF677。而碱性土上不同时期GF677和毛桃根系可溶性淀粉变化趋势相近,仅在9月2日GF677根系可溶性淀粉含量显著高于毛桃。
结合图8~11表明GF677植株体内更多的营养物质是以可溶性淀粉形式储存于枝条中,而毛桃植株体内主要以可溶性糖形式储存于根系中。
2.6 不同时期根系高铁还原酶活性变化
由图12可知,碱性土上不同时期GF677根系高铁还原酶(FCR)活性呈现先上升后下降的趋势,最高峰(101.90 μmol·g-1FW·h-1)出现在9月2日,毛桃根系FCR活性呈现先上升后下降然后又上升最后再下降的趋势,最高峰(80.35 μmol·g-1FW·h-1)出现在5月5日,在5月5日毛桃根系FCR活性极显著高于GF677,随后下降并于6月29日之后的4个时期均显著或极显著低于GF677。
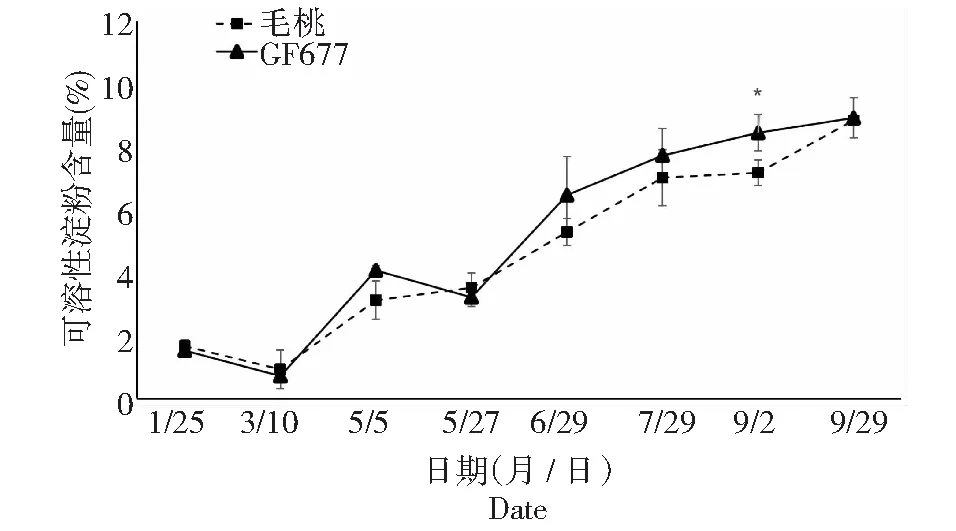
图11 碱性土上GF677和毛桃不同时期根系可溶性淀粉含量变化Fig.11 Soluble starch content of roots of GF677 and wild peach on alkaline soil during different periods
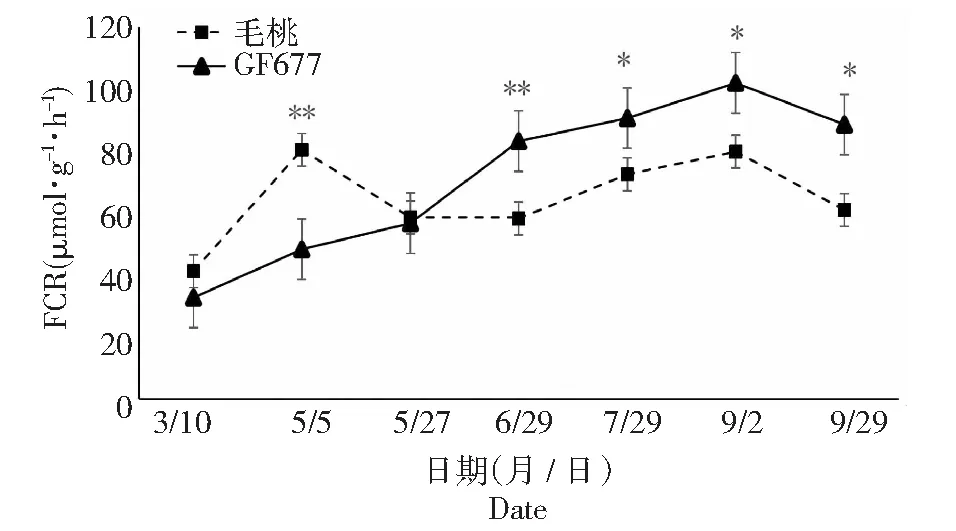
图12 碱性土上GF677和毛桃不同时期根系FCR活性变化Fig.12 FCR activities of roots of GF677 and wild peach on alkaline soil during different periods
2.7 叶片石蜡切片观察
植物在出现黄化病症时往往叶片会出现叶肉细胞空腔、解体,叶片厚度变薄,叶片卷曲等症状[11-12]。由图13-a可知,在碱性土上不同时期GF677叶片叶肉细胞充实,排列紧密,栅栏组织与海绵组织分布清晰(图13-a,c,e,g,i,k,m),没有出现空腔或解体的现象。而毛桃在碱性土上叶片出现空腔,栅栏组织与海绵组织均出现缩小的现象(图13-b,d,f,h,j,l,n),尤其是海绵组织,除7月29日外(图13-j),均出现较大程度的空腔,表明毛桃在碱性土上叶片肉细胞受到极大的损伤,尤其是海绵组织退化明显。
3 讨 论
植物在受到胁迫时,体内会产生超氧阴离子自由基、过氧化氢和酚类、胺类抗氧化酶系以及丙二醛等代谢产物对植物细胞产生毒害[13-14]。对碱性土上不同时期GF677和毛桃叶片抗氧化酶系统的几种酶类活性进行测定后发现,GF677叶片两种含有铁卟啉为辅基的酶类:POD、CAT酶活性在全部或部分生长期内均高于毛桃(尤其是POD酶活性在全生长期均极显著高于毛桃),体内丙二醛含量和相对电导率均低于毛桃,而SOD酶活性略低于毛桃,表明GF677在碱性土上叶片体内抗氧化酶活性升高并及时清理各种有毒物质,而毛桃体内铁有效性降低,导致含有铁卟啉为辅基的酶类活性低于GF677,毛桃清除体内活性氧主要依靠SOD,因而GF677表现出更强的抗逆性。刘赟[15]等人研究硫酸亚铁铵对香樟叶片黄化的影响时也发现,黄化香樟叶片POD活性明显下降,而施用硫酸亚铁铵后能显著提高黄香樟叶片的POD活性。陈栋等人[7]的研究发现黄化桃叶CAT活性与SOD活性变化趋势相反,而肖家欣等人[16]的研究表明黄化叶片CAT活性甚至高于正常叶,由此可见不同黄化植株CAT酶活性变化较为复杂,仍有待进一步研究。
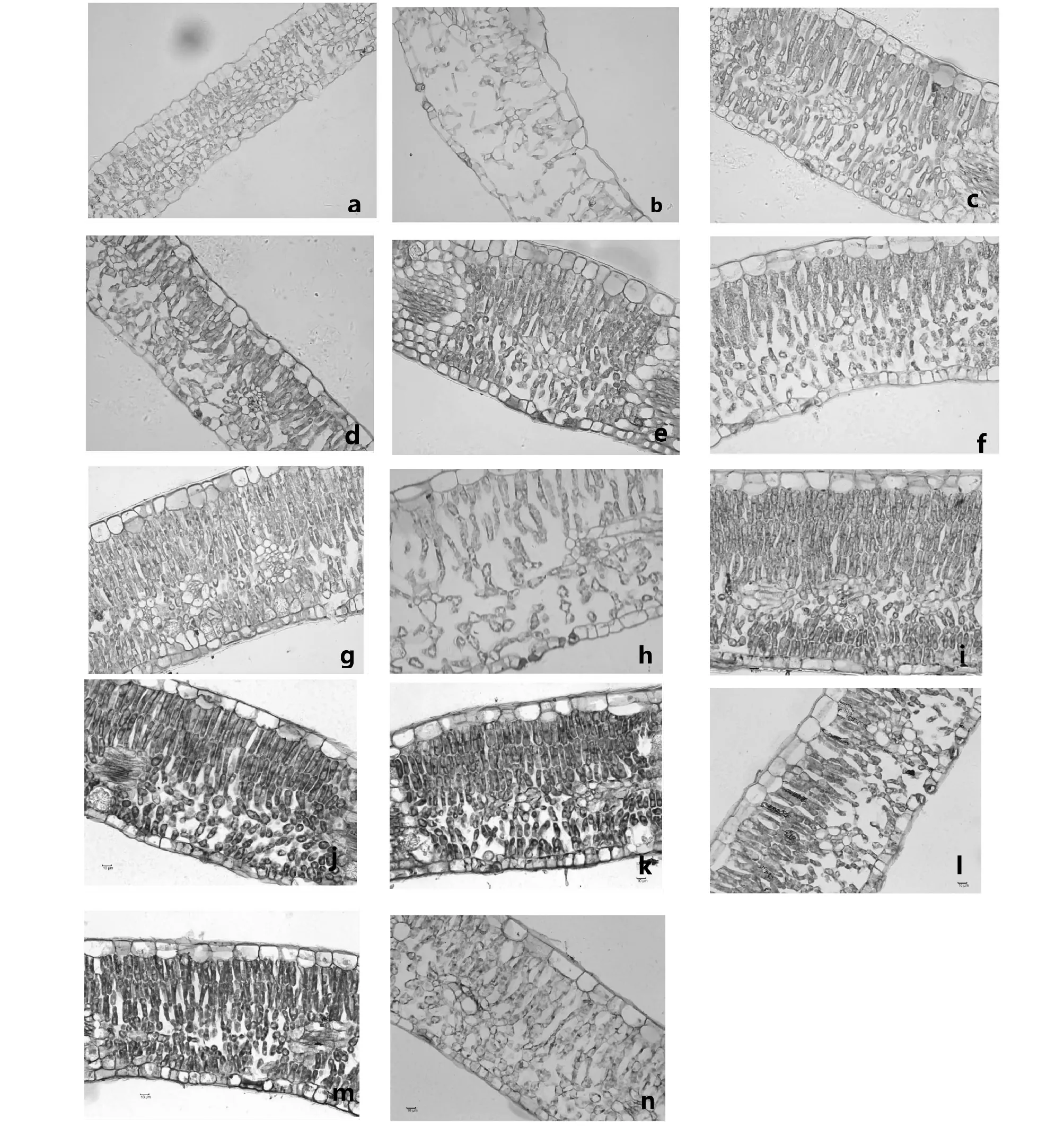
a:“GF677”3月10日;b:毛桃3月10日;c:“GF677”4月28日;d:毛桃4月28日;e:“GF677”5月27日;f:毛桃5月27日;g:“GF677”6月29日;h:毛桃6月29日;i:“GF677”7月29日;j:毛桃7月29日;k:“GF677”9月2日;l:毛桃9月2日;m:“GF677”9月29日;n:毛桃9月29日(以上图片均为400倍镜下观察所得)a:‘GF677’on March 10th;b:wild peach on March 10th ;c:‘GF677’on April 28th ;d:wild peach on April 28th ;e:‘GF677’on May 27th ;f:wild peach on May 27th ;g:‘GF677’on June 29th;h:wild peach on June 29th ;i:‘GF677’on July 29th ;j:wild peach on July 29th ;k:‘GF677’on September 2nd;l:wild peach on September 2nd;m:‘GF677’on September 29th ;n:wild peach on September 29th (All the pictures above were magnified 400 times in the microscopic observation)图13 碱性土上GF677和毛桃不同时期叶片石蜡切片观察Fig.13 Paraffin sections of leaves of GF677 and wild peach on alkaline soil during different periods
MDA和相对电导率也是反映植物逆境胁迫程度的重要指标。通常植物在遭受胁迫后,细胞脂膜过氧化产物MDA会升高,细胞受损并会导致相对电导率的升高[17-18]。本试验中毛桃叶片MDA含量在多个时期显著或极显著高于GF677,且叶片相对电导率在多个时期极显著高于GF677,表明毛桃在碱性土上受到了较强的碱胁迫,叶片细胞受损严重。
可溶性蛋白是重要的渗透调节物质和营养物质,可溶性蛋白增加和积累能提高细胞的保水能力,对细胞的生命物质及生物膜起到保护作用,因此经常用作筛选抗性的指标之一[19]。李音音[20]在研究黄化甜瓜生物学特性时甚至发现叶片黄化的甜瓜植株9388-1其可溶性蛋白含量高于正常甜瓜品种白莎蜜1号。然而本试验中GF677与毛桃在受到碱胁迫后枝条与根系可溶性蛋白含量变化幅度不大或趋势相近,难以鉴别桃树抗黄化能力。
可溶性糖是干旱胁迫诱导的小分子溶质之一,其种类主要包括葡萄糖、海藻糖、蔗糖等。这些可溶性糖类参与渗透调节,并可能在维持植物蛋白质稳定方面起到重要作用[21]。而可溶性淀粉作为植物体内重要的营养物质,可以反映植物的养分充足情况[22]。结合图8~11表明GF677植株体内更多的营养物质是以可溶性淀粉形式储存于枝条中,而毛桃植株体内主要以可溶性糖形式储存于根系中。
高铁还原酶又称被为三价铁螯合还原酶(ferric-chelate reductases,FROs)家族属于flavocyto chromes超家族中的1个亚类[23],是双子叶植物和非禾本科单子叶植物应答铁胁迫的关键酶之一[24],能够将细胞质膜上的Fe3+-螯合物还原成2价并通过Fe2+转运蛋白跨膜转运到细胞质中。在缺铁条件下,植物FRO基因表达是正常生长条件下表达量的4~5倍,以高效利用土壤中的铁[25]。陈微微[26]在研究拟南芥缺铁细胞信号转导调控因子时发现,缺铁条件下,双子叶植物拟南芥通过诱导根系FCR活性,促进根际Fe3+的还原来适应低铁环境。叶义全[27]在研究蔗糖和一氧化氮对植物缺铁响应的调控作用及其机制时也发现缺铁会导致拟南芥根系FCR活性升高。本试验在碱胁迫下,毛桃根系FCR活性会在短时间内急剧上升又迅速下降,而GF677根系FCR活性会持续上升,表明缺铁会诱导GF677与毛桃根系FCR活性的上升,但GF677能够在更长的时间内适应碱胁迫,保持较高的FCR活性,从而保证对铁素的还原与吸收。
植物在出现黄化病症时往往叶片会出现叶肉细胞空腔、解体,叶片厚度变薄,叶片卷曲等症状。在碱性土上不同时期GF677叶片叶肉细胞充实,排列紧密,栅栏组织与海绵组织分布清晰。而毛桃在碱性土上叶片出现空腔,栅栏组织与海绵组织均出现缩小的现象,尤其是海绵组织,除7月29日外(图13-j),均出现较大程度的空腔,这与黎秀丽等人[28]对库尔勒香梨黄化病叶结构的观察所得结果一致。
4 结 论
综上所述,GF677叶片在受到碱胁迫时体内含铁卟啉为辅基的酶类POD、CAT酶活性更高,保证了体内活性氧的及时清除,而毛桃叶片MDA与相对电导率在受到碱胁迫后明显升高,叶肉细胞受损严重。GF677植株体内更多的营养物质是以可溶性淀粉形式储存于枝条中,而毛桃植株体内主要以可溶性糖形式储存于根系中。碱胁迫同时诱导了毛桃与GF677根系高铁还原酶活性的上升,但毛桃根系FCR活性会在短时间内急剧上升又迅速下降,而GF677根系FCR活性会持续上升,表明GF677能够在更长的时间内适应碱胁迫,保持较高的FCR活性,从而保证对铁素的还原与吸收。GF677作为引入我国的抗性砧木资源,在四川盆地碱性土上表现出极强的抗碱抗黄化能力,在植株抗氧化系统、植株养分状况和根系铁还原能力上均强于毛桃,在生产上具有较好的推广前景。本试验为今后评价桃砧木资源抗碱抗黄化能力提供了可靠的依据。
猜你喜欢
星星·散文诗(2022年13期)2022-12-21
星星·诗歌原创(2022年5期)2022-06-04
中国果业信息(2021年10期)2021-12-07
昆明医科大学学报(2021年8期)2021-08-13
今日农业(2020年22期)2020-12-14
商品与质量(2019年36期)2019-04-15
热带林业(2019年4期)2019-03-05
天然产物研究与开发(2019年1期)2019-03-01
中国果业信息(2017年4期)2017-05-09
中国果业信息(2017年10期)2017-01-13
