
稻田3种捕食性天敌对灰飞虱种群的调控作用
2018-11-20 02:57:56邢亚楠代克涛车喜庆
江苏农业科学 2018年20期
邢亚楠, 代克涛, 车喜庆
(1.辽宁省盐碱地利用研究所,辽宁盘锦 124010; 2.中化现代农业有限公司,北京 10010)
水稻作为辽宁省第二大粮食作物,年播种面积、产量分别达到56万hm2、420万t。盘锦市作为辽宁省水稻的主产区,其飞虱的发生已严重影响到滨海稻区水稻的生产[1]。辽宁省盐碱地利用研究所通过多年来对飞虱的预测预报发现,盘锦稻区主要以灰飞虱、白背飞虱、褐飞虱等3种飞虱混合发生,造成不同程度的危害。相比于白背飞虱、褐飞虱,这两者属于迁飞性,而灰飞虱以若虫状态在盘锦稻区越冬,其基数大、发生重[2-3]。一方面,灰飞虱以成虫、若虫状态群集在稻丛下部,直接刺进水稻茎秆组织内吸食汁液。另一方面,灰飞虱还能传播条纹叶枯病病毒等,给水稻造成很大的危害,产量损失一般在5%左右,发生严重的年份产量损失超过10%[4-5]。
目前对灰飞虱的防治主要采用化学防治法,鉴于大量广谱性杀虫剂的使用,不仅使灰飞虱产生了抗药性,也使得田间天敌数量减少,严重破坏了生态平衡,引起一系列食品安全和环境污染等重要问题[6-8]。为顺应农业协调可持续发展的主题要求,生物防治应运而生,保护和利用自然天敌昆虫资源作为生物防治的一种重要手段有着重要作用。目前,关于天敌利用等问题的研究多局限在室内捕食功能测定,而对于在田间自然环境中,多种天敌与害虫复合系统的报道较少。
多种天敌对灰飞虱捕食关系是相当复杂的,不是单种天敌捕食量的简单叠加。二次正交旋转组合设计能较好地描述多因子共存作用时捕食性天敌的捕食作用[9]。孙文鹏等曾运用二次旋转组合设计研究了大豆田4种捕食性天敌与大豆蚜共存系统的捕食关系,并应用于大豆蚜的田间防控[10]。常瑾曾运用二次正交旋转组合设计研究了稻田生态系统中3种蜘蛛对白背飞虱的捕食效应[11]。
通过笔者所在研究室多年来对滨海稻区节肢类捕食性天敌进行的普查发现,横纹金珠、草间钻头蛛、粽管巢蛛在田间所占比例分别为37.5%、11.6%、22.9%,为稻田优秀的捕食者[12-13]。因此,本研究以3种优势蜘蛛为天敌,以灰飞虱为靶标害虫,明确其相互作用关系,以期为灰飞虱的田间生态调控提供理论依据。
1 材料与方法
1.1 供试材料与调查方法
采集田间灰飞虱,在水稻(盐丰47,由辽宁省盐碱地利用研究所提供)植株上饲养,连续转移3~5次,确保灰飞虱不带病毒后再大量繁殖(室外笼罩饲养),挑选健康灰飞虱作为试虫[14]。
横纹金珠、草间钻头蛛、粽管巢蛛均采集于水稻田,室内配对产卵,先产的卵于冰箱4 ℃条件下保存,卵量备足后一起孵化,保持发育一致,取健康蜘蛛用于试验。
于2014年7月20日和2015年7月20日连续2年在大洼新建农场试验田进行试验,每隔2 m罩网,网罩规格为 1 m×1 m×2 m,网孔为100目,保证每网内2穴水稻。试验设2次重复,共144网。人工清除网内天敌昆虫及灰飞虱后,按设计要求(表1)接入定量的天敌和灰飞虱。接入灰飞虱时,先惊扰灰飞虱种群,待灰飞虱活动后用毛笔轻挑至目标水稻植株上,类似自然分布状态,确定成活7 d后调查网内水稻植株上灰飞虱的数量。

表1 天敌组合编码值与其密度的关系
1.2 试验设计与统计分析
采用4因子2次正交旋转组合设计法,得到回归方程:y=b0+∑bixi+∑bijxixj+∑biixi2;其中,bi、bij、bii分别为一次项、交互项、二次项系数;xi为参试物种;y为共存系统害虫被捕食量的估计值。对方程进行方差分析,分析捕食性天敌粽管巢蛛(x1)、草间钻头蛛(x2)、横纹金珠(x3)与灰飞虱(x4)之间的相互关系以及与捕食性天敌之间的相互关系。应用DPS 7.05统计分析软件进行数据分析。因子水平和编码值见表1、表2,每个试验中心点重复2次。
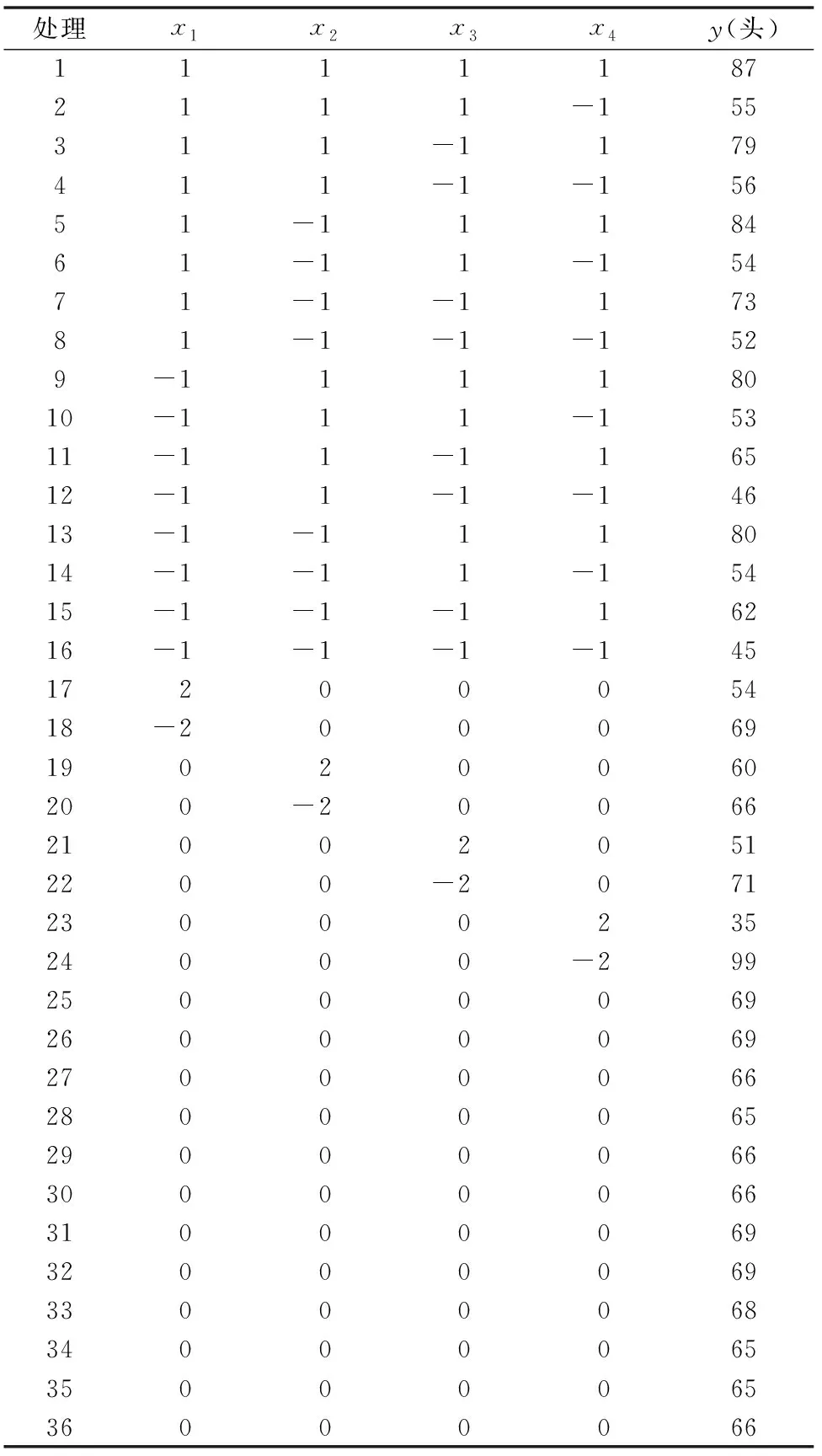
表2 4因子2次正交旋转组合设计试验方案及结果
2 结果与分析
2.1 天敌与灰飞虱共存数学关系模型的建立
根据表2的结果,采用DPS系统建立以为粽管巢蛛(x1)、草间钻头蛛(x2)、横纹金珠(x3)、灰飞虱(x4)为决策变量,7 d后以灰飞虱的捕食量为目标函数(y)的四元二次回归方程:y=66.916 67+3.541 67x1+1.208 33x2+4.541 67x3+13.458 33x4-1.197 92x12-0.822 92x22-1.322 92x32+0.177 08x42+0.687 50x1x2-1.812 50x1x3+1.062 50x1x4-0.687 50x2x3+0.437 50x2x4+2.187 50x3x4。
在α=0.1水平上剔除不显著项,重做统计得新回归方程:y=66.916 67+3.541 67x1+1.208 33x2+4.541 67x3+13.458 33x4-1.197 92x12-0.822 92x22-1.322 92x32-1.812 50x1x3+1.062 50x1x4+2.187 50x3x4。
对各回归方程中的系数进行显著性检验,由表3可知,一次项粽管巢蛛(x1)、草间钻头蛛(x2)、横纹金珠(x3)与灰飞虱(x4)对7 d后灰飞虱的捕食量有显著的影响。方程整体回归达到显著水平[F2=68.637 73>F0.01(df=14,21),P=0.001<0.01],说明本试验设计与结果良好,回归方程能反映粽管巢蛛、草间钻头蛛、横纹金珠之间的相互关系,具有一定的参考价值。
二次项x1x4、x3x4、x1x3在0.01水平下显著,其他项则可以忽略。在回归方程中二次项与交互项系数为正,表明天敌自身或者相互间对灰飞虱捕食量增加有促进作用;系数为负,表明天敌自身或者相互间对其捕食灰飞虱有负影响。
2.2 各因子对捕食效应的影响
当固定其他因子为0水平时,粽管巢蛛、草间钻头蛛、横纹金珠、灰飞虱捕食量的单因子分析效应如图1所示。
由图1斜率可以看出,x4(灰飞虱)>x3(横纹金珠)>x1(粽管巢蛛)>x2(草间钻头蛛),说明灰背飞虱自身密度x4与其被捕食量y的关系,呈一条逐渐上升的曲线,且其上升幅度较稳定,对其自身的捕食量y值影响最大。捕食性天敌中以横纹金珠的影响最大,其次为粽管巢蛛,草间钻头蛛的影响最小。
随着天敌数量由-2增加到1水平时,灰飞虱被捕食的数量y值逐渐增加,增幅较大。当天敌数量由1水平增加到2水平时,灰飞虱被捕食的数量y值基本不再增加,说明随着天敌密度的增加,个体间的干扰、竞争作用也逐渐增强,使得捕食能力不断降低。
2.3 天敌的交互作用
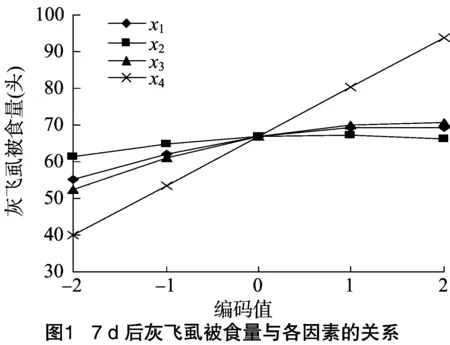
对回归方程中二次相关系数进行显著性检验,x1x3、x1x4、x3x4在0.01水平下显著,其他项可以忽略,其中x1x4、x3x4的二次项系数为正值,即存在显著正交互作用,x1x3的二次项系数为负值,表明二者之间存在显著的种间制约作用。
由图2可知,当x1(粽管巢蛛)的编码值由-2水平到1水平时,随着x3(横纹金珠)种群密度逐渐增加,灰飞虱被捕食量(y)也逐步增长;当x1(粽管巢蛛)的编码值由1水平增至2水平,x3(横纹金珠)的编码值由-2水平到0水平时,灰飞虱被捕食量增幅逐渐减小。当x3(横纹金珠)的编码值由1水平到2水平时,灰飞虱被捕食量却逐渐降低。这说明横纹金珠的种群密度增加到一定数量时,其种内干扰作用较强,与x1粽管巢蛛的种间竞争也较强,具有一定的负调控作用。
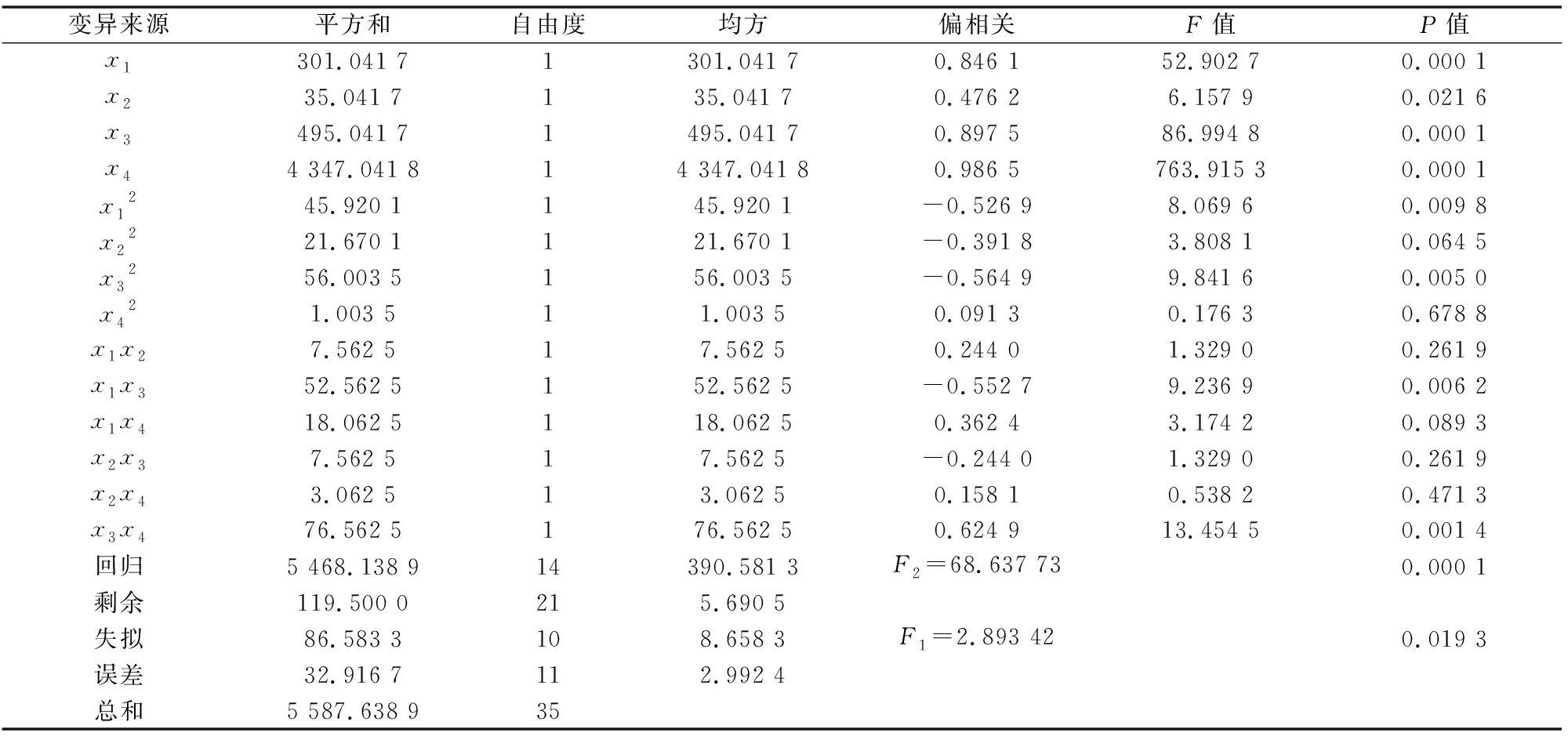
表3 天敌组合结果方差分析
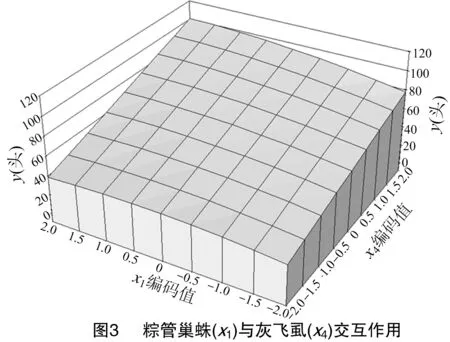
由图3可知,随着粽管巢蛛(x1)和灰飞虱(x4)种群数量的不断增加,灰飞虱被捕食量也逐步增加,可见粽管巢蛛对灰飞虱种群数量有重要的正调控作用。当x1=2、x4=2时,y值最大,为100。
由图4可知,随着横纹金珠(x3)和灰飞虱(x4)种群数量的不断增加,灰飞虱被捕食量也逐步增加,可见x3(横纹金珠)对灰飞虱种群数量也有显著的正调控作用。当x3=2、x4=2时,y最大,为106。

2.4 不同天敌对灰飞虱控制作用的优化
根据DPS软件中的数据进行优化分析,得到不同天敌对蚜虫控制作用的优化组合,灰飞虱数量为120头(x4=2)时,粽管巢蛛为6头(x1=1),草间钻头蛛为6头(x2=1),横纹金珠为8头(x3=2),此时飞虱被捕食量ymax为108,最高防效为90%。
3 讨论
罩笼试验是通过控制网孔大小,有效屏蔽不同大小的昆虫天敌,进而获得这些天敌对害虫的捕食作用,是害虫生物防治研究和实践的重要一环[15-17]。常瑾研究发现,稻田生态系统中3种蜘蛛对白背飞虱均具有较强的控制作用, 但随着蜘蛛密度不断增加,其种内及种间的竞争越来越激烈,因此飞虱被食量呈下降趋势[11]。这与本研究得到的结果相似,符合天敌与害虫共存的田间实际情况。但室内人工模拟环境,生境过于简单,环境因子过于稳定,与稻田间自然生态环境差别较大,往往不能对天敌控害作用提供最有说服力的证据[16]。而本试验是在田间进行的,为了解自然界的捕食作用提供了参考。
本研究在田间利用罩笼试验法,采用4因子2次正交旋转组合设计,进行害虫与多种天敌共存试验,得到了4因子对灰飞虱捕食作用的调控顺序依次为x4(灰飞虱)>x3(横纹金珠)>x1(粽管巢蛛)>x2(草间钻头蛛),均为正效应,最佳防效可高达90%。7月中下旬开始,气温适度的环境均有利于灰飞虱的繁殖,为防治的最佳时期,本试验结果可为灰飞虱生防实践提供依据。
猜你喜欢
工业设计(2021年2期)2021-03-24 11:07:07
数学大王·中高年级(2020年11期)2020-11-19 10:54:37
阅读(书香天地)(2020年2期)2020-04-07 03:34:02
Asian Herpetological Research(2019年2期)2019-06-26 03:03:36
中国中医基础医学杂志(2018年12期)2018-02-13 03:15:52
小学生(看图说画)(2017年4期)2017-11-01 06:07:02
发明与创新(2016年27期)2016-10-13 12:39:07
化工科技(2016年6期)2016-06-06 01:54:18
实用手外科杂志(2015年4期)2015-08-27 01:54:00
中国艺术时空(2013年4期)2013-03-11 19:49:23
