
植物性状研究的机遇与挑战: 从器官到群落
2018-11-14 12:37何念鹏刘聪聪张佳慧于贵瑞
生态学报 2018年19期
何念鹏,刘聪聪,张佳慧,徐 丽,于贵瑞
1 中国科学院地理科学与资源研究所 生态系统网络观测与模拟重点实验室, 北京 100101 2 中国科学院大学 资源与环境学院, 北京 100049
1 植物性状及其研究进展
1.1 植物性状及其主要研究方向
植物性状(Plant trait)或植物功能性状(Plant functional trait)是植物对外界环境长期响应与适应后所呈现出来的可量度的特征[1]。在实际研究过程中,人们更多关注与植物生长、繁殖、存活或与资源获得、利用及其利用效率密切相关的性状。植物是陆地生态系统初级生产力的主要参与者和贡献者,因此大多数研究均围绕植物性状如何适应环境、优化资源利用或优化生产力而开展[2]。在指标上,更多选择了能体现植物对环境适应并(或)影响植物生产力的特定属性,如叶片形态特征(叶片大小、厚度、比叶面积和光合速率等)、根属性(根大小、比根长和元素含量等)和种子大小与质量等[3-5]。
植物性状研究可追溯到原始农业时期,人们通过对比发现与农作物产量和品质密切相关的性状,再通过选育实现高产与优质。在生态学萌芽期开始,植物生态学家就开始了对自然群落的植物性状的研究,重点关注植物如何通过特定性状或多种性状协同来实现群落结构稳定与生产力优化[4]。20世纪80年代后,植物性状研究作为生物学、地学和环境科学交叉研究的纽带,成为研究热点。近年来,定量研究植物性状(涉及碳收支、养分利用和水分经济学)在不同尺度的空间变异规律、如何实现植物生产力优化是其中的重要研究内容,探讨植被群落或植物多样性对全球变化(气候变化、氮沉降和酸沉降等)的响应与适应是重要的发展方向[5- 7]。本文对植物性状的评述主要集中在陆地生态系统的自然群落性状研究。
1.2 植物性状研究的主要进展
1.2.1 种内和种间性状变异及其空间格局
植物性状种内与种间变异是当前最广泛的研究内容。在控制实验中,植物叶片形态(面积、厚度和比叶面积)、解剖结构(栅栏组织、海绵组织和表皮细胞)和气孔都会随着光照、温度和CO2浓度变化而做出适应性的变化[8- 11]。在自然状态下,羊草的叶片厚度、比叶面积、脯氨酸和K+/Na+随着温度和降水的变化而发生显著的变化[12];拟南芥的个体大小和相对生长速率也呈现一定的纬度变异[13];光泽柳的比叶面积随着土壤水分含量的增大而增大[14];3种栎属植物的叶片形态和化学组成与降水具有显著的相关性[15]。与常绿植物相比,落叶植物的比叶面积更大[16];阔叶植物比针叶植物具有更高的气孔密度[17];最大气孔导度呈现出乔木>灌木>草本,落叶>常绿,阔叶>针叶[18]。Cornwell和Ackerly[14]发现植物性状沿环境梯度呈现的规律主要来自于种间变异,而种内变异的影响较小。
已有研究围绕植物叶片性状(叶片大小、叶片厚度、比叶面积、氮含量(N)、磷含量(P)、光合速率和光响应饱和曲线)和根性状(根分级特征、根大小、根长度、比根长、N、P、根瘤特征和菌根真菌特征),探讨了植物性状在不同生境的变异规律、特定物种(属或科)根叶性状沿环境梯度的变化规律[19-21]。Wang等[20]利用847种植物的实际调查数据,探讨了叶面积大小、叶厚度、比叶面积和叶片干物质含量4个性状沿3700 km的中国东部森林样带(NSTEC)的变化规律。Wang等[17]采用了压痕法调查了NSTEC样带760种植物的气孔性状(气孔密度和气孔大小),首次探讨了区域尺度叶片气孔特征,并从物种-功能群-群落层次揭示了植物气孔性状沿纬度变异规律及其主要影响因素。Tian等[22]对NSTEC样带优势乔木的叶片解剖结构性状进行了调查分析(上表皮厚度、下表皮厚度、栅栏组织厚度和海绵组织厚度等),阐明了叶片解剖性状的纬度格局与影响因素。上述研究基本形成了叶片常规形态性状-气孔性状-解剖结构性状的由表及里的指标体系。
近年来,围绕叶片C、N、P含量的空间格局及其影响机制开展了较多研究。例如,Ordonez等[23]通过数据收集整合分析了全球尺度植物比叶面积、叶片N、P和N∶P的空间格局及其影响因素。He等[19]通过分析内蒙古、西藏和新疆3个地区213种草原植物C、N及其C∶N,发现3个地区间草原植物的C∶N是相对稳定的,气候通过改变植物组成来影响草原植物叶N含量。Chen等[24]调查了中国14个森林386种木本植物叶片N、P含量及其N∶P比,并从气候、土壤和植物生活型角度探讨了其影响因素。Han等[25-26]通过收集文献数据,探讨了中国区域植物叶片N∶P化学计量特征以及叶片多元素的生物地理格局及其影响因素。
根性状及其生物地理格局也是当前植物性状研究的热点之一,相关研究提高了人们对植物如何适应环境和养分利用策略的认识。Comas和Eissenstat[27]报道了25个共存物种的比根长特征。 Gordon和Jackson[28]针对植物细根元素含量(C、N和P)及C∶N∶P进行研究,揭示了植物根系在元素周转中的作用。郭大立等围绕中国森林植物根形态、根瘤和外生菌根等开展了系统的研究工作,较深入地揭示了植物对养分利用策略,并深入探讨了中国从南到北优势乔木细根形态的空间变异特征[29-32]。
在全球尺度上,种子质量随着纬度的升高而降低,植物功能群和植被类型的变化是造成种子质量纬度变化的重要原因[33];Moles等[34]发现植物最大高度会随着纬度升高而发生明显的变化。
1.2.2 植物多种性状协同与权衡
在自然环境中能长期生存并繁衍的植物,既需要具有适应环境变化的能力,还必须具备能获得其生长和繁殖所必需光、水和养分的能力,是通过植物多种性状协同与权衡来实现的。植物叶片和根系具有很多可测量的性状,每个性状都能不同程度的反映植物对环境的适应或者植物对资源的获取能力。例如,比叶面积和叶片N含量等多种性状都对植物光合速率具有较大的影响[12];同时,叶片气孔密度和解剖结构等间接影响植物光合速率和水分利用效率[18,35-36]。类似地,细根是植物获取养分的最重要器官;其中,根长度、比根长和根瘤特征与养分获取能力密切相关,同时也与植物蒸腾能力等间接相关。植物不仅通过特定器官内部的多种性状的协同(如比叶面积与N含量)[37];还通过叶-根之间16种元素的协同控制机制,来实现对环境的响应与适应[38]。Wang等[39]发现叶和根的形态特征趋异而元素含量趋同是植物叶-根协同的重要机制。虽然相关研究已经取得了长足进展,但由于植物性状的多样性及其所面临环境的复杂性,还有许多问题仍待解决。
Wright等[16,40]利用全球叶片性状数据库提出了著名的叶经济型谱(Leaf economic spectrum, LES);植物叶片性状沿着一条连续变化的功能性状组合谱有序排列, 其一端代表着比叶重小、含N量高、光合速率大、呼吸速率高、叶寿命短的“快速投资-收益型”策略;而另一端代表着寿命长、比叶重大、含N量低、光合速率和呼吸速率都偏低的“缓慢投资-收益型”策略。在此基础上,Diaz等[6]利用4.6万种成年植物高度、叶面积、叶N含量等6个指标,从物种水平验证与发展了叶经济型谱理论,并发现树高-种子质量维度与叶经济谱系维度相互独立。LES的提出与发展,极大地提高了人们对植物通过性状协同实现生产力优化机制的认识,尤其是为解释碳氮元素在植物内固定、利用和流动提供了重要的理论依据。然而,关于LES的形成动力仍存在较大的争议;一方面,Maire等[41]认为环境梯度引发植物资源权衡效应,植物通过自我调节来响应环境变化,因此LES主要受环境因子主导;另一方面,目前LES还多集中在资源获取的维度,然而自然群落中影响植物光合作用的因素是多维的,未来还应考虑水分维度和温度维度等[42]。
近期,根系经济学谱系(Root economic spectrum, RES)和木材经济学谱系(Wood economic spectrum, WES)也被提出。RES把根系的呼吸强度、比根长和N含量等归为获得性强的性状(Acquisitive traits),而把根系干物质含量、根系直径和木质素/N含量归为更保守的性状[43]。WES认为:较高的木质密度有利于抵挡害虫和病原体的侵害,更加具有生物力学上的稳定性,从而提高了物种的寿命;较低的木质密度和较高的水分含量意味着更少的投资成本,因此物种有较高的树高和直径生长速度[44]。
1.2.3 植物性状与功能的关系
自然群落中的植物性状研究不应局限在特定器官、特定物种或特定科属上,而应重点探讨多种植物如何通过特定性状或多种性状协同,来实现群落结构稳定与功能优化机制。目前,植物性状-功能研究大多是建立在器官水平,如叶片N含量与光合速率的关系、叶片比叶面积与光合速率关系、叶片叶绿素含量与光合速率的关系,这些个体或者物种水平得出的结论能否适用于到群落水平,还有待实测数据证实。近年来,如何将个体或者物种水平测定的性状与群落结构和功能相结合,或从性状角度解释群落结构构建机制,是当前植物性状研究的热点与难点[45]。Kunstler等[46]通过分析全球14万个样地、300万颗树的最大生长速率、密度、高度和比叶面积数据,发现在区域和全球尺度上述性状与竞争关系整体表现出较好的一致性,为预测森林物种间相互关系提供了美好的愿景;同时发现性状的不相似性(dissimilarity)在局部群落对生产力影响方面的作用较小,但这种不相似性导致的权衡(Trade-off)确是影响物种间竞争和促进物种共存的重要基础。Cadotte等[47]通过45个乔木物种15种性状的分析,发现在物种水平叶片6种功能性状与光合速率具有显著的相关性。功能性状的群落加权平均值一直被认为是获得群落水平性状的潜在的重要途径,丰富度指数的加权方式最为常见,但这种加权方式忽略了自然群落中物种个体间的差异,基于物种生物量异速生长方程的群落生物量加权方式可很大程度克服上述不足。Wang等[20]通过结合生物量异速生长方程的群落生物量加权方式,探讨了森林植物叶片大小、比叶面积等的纬度格局,发现气候因素和土壤N含量具有重要影响。此外,气孔性状的群落生物量加权平均值与生态系统水分利用效率显著正关性[18];叶片解剖结构的群落加权平均值与生态系统生产力和水分利用效率具有紧密的联系[36]。
何念鹏等借助中国东部南北样带9个典型森林生态系统全面系统的调查数据(包括群落结构数据、样地内1120多个物种叶-枝-干-根样品的16种元素含量、叶片常规性状-气孔性状-解剖结构性状、叶片热值-叶绿素-非结构性碳水化合物、1—5级细根形态性状-元素含量等),借助群落结构数据+生物量异速生长方程+比叶面积数据,实现了部分植物性状从器官-物种-功能群-群落的推导,并进一步建立了天然森林群落植物性状与功能的联系。一些研究结果揭示了天然森林群落中特定性状与功能的定量关系,具有一定的理论和实践意义[18,36]。然而,目前有关天然森林群落植物性状与功能联系的研究,仍集中在单一性状,未来更应从多性状协同角度探讨大尺度下森林群落性状与功能的关系。
1.2.4 植物性状如何表征陆地植被对全球变化的响应与适应
结合陆地生态系统植被的结构和功能,定量分析植物性状对全球变化(气候变化、氮沉降和酸沉降等)的响应与适应是生态学近年来的研究热点。为提高模型模拟精度,将部分重要的植物性状纳入模型正逐渐成为未来模型发展的重要方向[38,41]。然而,由于传统的植物性状研究大多在物种水平进行,缺乏群落尺度的数据,一定程度地制约着模型在大尺度上的发展和应用。
植物化学计量特征为科学家探讨全球变化的效应提供了重要途径。Elser等[48-49]通过整合分析不同湖泊和全球控制实验(N或P添加),揭示了海洋和陆地生态系统中的磷限制性效应;并在此基础上,定量评估未来N沉降增加情景下陆地生态系统净初级生产力的变化。Yan等[50]通过整合分析全球淡水中植物N和P累积及其生态效应,发现在大气沉降和人为富营养化情况下,淡水系统的P累积远高于N累积,从而对淡水生态系统生产力形成巨大的负效应,需要加强治理力度使这些淡水系统尽快恢复到初始状态。Zhu等[51]基于实测数据发现中国区域大气沉降N∶P远高于植被和土壤,并预测不平衡的N∶P输入将进一步增强陆地生态系统磷元素的限制效应。此外,叶片N∶P比常用于确定不同地区的限制性元素,当N∶P < 14是为氮限制,当N∶P > 16时为磷限制。近年来,国内外学者从个体或群落尺度都开展了很多相关研究[25,52-53]。同时,随着生态化学计量学的日益成熟,人们提出可用植物群落不同组分C∶N比值,结合N沉降通量来构建陆地生态系统固碳效应的理论框架[54];最近,Zhu等[55]基于中国区域氮沉降量以及NESCT样带9个典型森林生态系统不同植物组分C∶N比值,基于化学计量学基本理论评估了大气氮沉降量对中国森林生态系统的固碳效应。
2 植物性状研究面临的机遇与挑战
国内外学者已经在植物性状研究领域开展了大量的研究工作,并取得了丰硕的成果。仔细分析上述4个主要进展的数据源,会发现在这些研究依然以优势种为主体与重点,并未考虑天然森林群落结构与物种组成的复杂性。近年来,国内研究提出利用长期固定监测样地开展系统的性状调查工作(如中国生态系统研究网络(CERN)、中国国家生态系统观测研究网络(CNERN)、中国森林生态系统定位观测网络(CTERN)、中国森林生物多样性监测网络(CForBio)),这此平台为人们开展森林植物性状、性状-功能关系提供了非常理想的平台,但近期其成果能尺度上推到森林群落的还很少[6,16]。总之,个体水平所获得的性状-功能的关系,在自然群落中是否适用还有待证实;同时,植物多种性状协同(或权衡)是植物对环境响应与适应的重要途径,但该思想在实际研究中很少被贯彻,未来可从如下几个方面深入探索。
2.1 重视多种性状协同或趋异规律的研究
以叶片为例,前期科研人员非常重视叶片常规性状、元素含量、光合特性等研究[6,16,23]。但长期忽视叶片气孔属性(气孔大小、气孔密度和气孔比面积)和叶片解剖属性(上表皮厚度、下表皮厚度和栅栏组织厚度等)。未来的研究不仅要从各个性状的种内、种间、区域变异分析,还更应该深入探讨多种性状的协同规律,从植物对环境的响应和适应角度分析。
2.2 以植物群落构造为基础,开展系统性的植物性状调查
在群落调查时,要尽可能覆盖所有物种,并且每个物种尽可能涵盖叶、枝、茎、根和种子等。目前,在包括比较权威的世界植物属性数据库(TRY数据库),均是以收集公开发表数据为基础而建立的,每个群落一般仅搜集一个或几个物种的数据;由于未考虑群落结构,相关研究是将物种水平等同于群落水平[6,16,46]。此外,TRY数据库的植物属性数据,绝大多数集中在叶片,非常缺乏植物叶-枝-茎-根-种子的配套数据。
2.3 在群落尺度开展性状-功能关系研究,突破性状研究的难点
受测试技术与研究手段的限制,前期研究多是在个体水平探讨性状-功能的关系;在生态系统、区域或全球尺度上,一直存在性状数据与功能数据空间不匹配的问题[45]。例如,植物性状的测定大多在个体或者种群水平开展,而大尺度研究的功能更多在群落或生态系统水平进行,二者间存在空间尺度与量纲间的不匹配。因此,如何实现植物性状从个体或者物种-功能群-群落-生态系统水平的尺度拓展,建立性状-生态系统功能的定量联系,是揭示生态系统对外界环境和资源变化响应与适应机制的基础,也是改进和优化模型、提高其预测精度的重要条件。近期,国外学者正在尝试建立植物性状与生态系统功能之间的关系[18,36-37,56- 59]。总体来说,这方面的研究还非常少,且研究方法和主要结论仍需要进一步改进与证实。为了加快这一过程,我们认为需要发展新的概念体系、指标体系,真正实现植物性状与宏观生态学的联系;通过与高新技术的结合,既拓展植物性状研究的范畴,又使植物性状研究能更好地服务于区域甚至全球生态环境问题的解决。
3 群落性状或生态系统性状的定义与内涵
当前,生态学研究正在快速向宏观方向发展,关注的最基本尺度是群落或生态系统;另外,全球变化对陆地生态系统的影响评估也是以群落或生态系统结构与功能为重要对象。现阶段大多数的植物性状研究主要集中在植物器官或者个体水平,仅有少数研究将它们拓展到群落水平;在拓展过程中有3种方法:1)简单算数平均法;2)相对生物量或多度加权平均法;3)生物量加权平均法。方法1和方法2所获得的群落水平的植物性状(如叶片氮含量g/kg或%),虽然可以有效地探讨群落结构维持机制,由于无法实现对群落性状在量纲上向单位土地面积转换,使它很难与模型和遥感数据相匹配。而方法3可以获得基于单位土地面积的群落性状,在空间尺度匹配(或量纲匹配)的前提下实现个体水平测定的植物性状数据与生态模型和遥感观测相联系,更好地探讨区域尺度下自然生态系统结构和功能的关系及其对全球变化的响应与适应[36]。植物性状研究的壁垒和宏观生态学发展的现实需求,促使人们重新思考植物性状研究的尺度问题;需要将植物性状统一到群落尺度或单位土地面积尺度,才能与宏观生态学研究的主要观测手段紧密联系起来(通量观测、模型模拟和遥感观测),推动相关学科发展。
群落性状(Community trait)可定义为:在群落尺度能体现植物对环境适应性和生产力优化的性状(其定义也适用于动物或土壤微生物等)。其核心内涵:1)群落性状均被转化为以单位土地面积为基数的性状,如叶片大小、叶片生物量、叶片气孔密度、叶片N和P含量等;2)任何群落性状均是可测量或可推导,原则上均是采用群落生物量加权平均的方法进行推导的;3)群落性状应能从不同层面反映植物对环境的适应或生产力的优化,即具有明确的生态学意义。群落性状的提出和发展,一方面有利于将各种植物性状在单位土地面积上标准化,便于深入探讨不同性状间的内部关系、协同或趋异规律,进而更好地从性状角度揭示植物群落的构建与维持机制。另一方面,通过建立单位土地面积群落性状,可解决长期以来植物性状数据与宏观生态学主要测试数据(遥感、通量观测和模型等)空间尺度不匹配的问题,推动宏观生态学的发展、更好地服务于区域生态环境问题的解决。从长远看,只有充分利用各种高新技术的学科,才能获得更快的发展;因此,群落性状提出与发展将会成为相关领域新的生长点。为了更好地统筹自然生态系统中的植物、动物、微生物、土壤等要素,未来应进一步发展以单位面积为基础的生态系统性状(Ecosystem trait),生态系统性状将在相对统一的空间尺度和量纲上获得植物群落性状、动物群落性状、土壤微生物群落性状、土壤属性和气候要素等,为深入研究不同尺度生物-非生物性状间的关系奠定坚实基础,并以群落性状为核心连接宏观生态学的主要研究途径(遥感观测、通量观测和模型模拟),推动传统性状指标与宏观生态学的应用相结合。
4 群落性状指标体系初探
图1给出了一个典型森林生态系统的性状指标的基本框架图,并展示了生态系统性状、生态系统结构和生态系统功能的整体构架及其相互关系。在生态系统结构参数的协助下,可利用植物群落生物量加权法(非传统的直接算术平均法和相对生物量加权平均法),将器官水平测定的植物性状参数推导至群落水平,从而在同一尺度建立生态系统内植被-大气间的功能参数(总初级生产力、净初级生产力和蒸腾速率等)和土壤-大气界面的功能参数的定量关系,构建群落性状-功能间的桥梁。
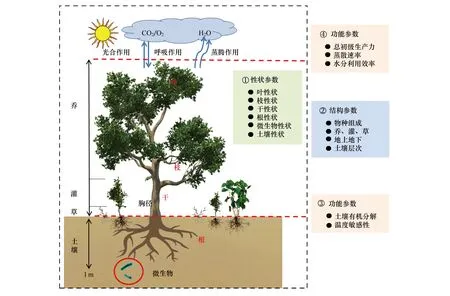
图1 森林群落性状与功能的基本框架图Fig.1 Logical picture of community traits and functioning in natural forest图中植物群落性状参数均是通过群落结构参数,利用生物量加权法将器官水平测定的性状推导到整个群落,并以单位土地面积为基础参数
按图1的框架,建立生态系统性状-结构-功能的基本指标体系、测试途径和推导方法(表1)。以叶片性状为例,从叶片形态性状(叶形状、叶大小和比叶面积等)、气孔形状(气孔大小、气孔密度、气孔比面积等)、解剖结构性状(上表皮厚度、栅栏组织厚度、下表皮厚度等)和多元素含量性状(C、N、P含量等)着手。在系统测试的基础上,再利用比叶面积+异速生长方程+群落结构数据推导获得各种基于单位土地面积的群落性状。目前建议的指标体系,会在未来发展过程中逐步增加与完善,例如,此次没有包括叶片光合速率,理由如下:1)叶片光合速率受温度和水分影响非常大,在短时间内都存在较大的差异;2)叶片光合速率在群落或生态系统水平常用总初级生产力、净初级生产力等指标来替代,并可通过通量观测系统测定。在实际操作过程中,个体或器官水平测定的光合速率难以推导到自然群落水平;虽然植物最大光合速率是一个相对稳定的指标,但自然界中由于物种冗余、相互竞争等因素,植物最大光合速率与群落净初级生产力的关系非常复杂,不同地点和不同生态系统类型间存在显著变异。
植物性状要实现从器官-群落的推导,理论上需要满足两个基本条件:1)调查应覆盖植物群落出现的所有物种;2)详细的植被群落结构调查数据。在实际操作过程中,针对每个群落需要开展详细的调查,这是通过文献搜集数据难以实现尺度上推的重要原因。网络化长期定监测样地(CERN、CNERN、CTERN和CForBio)为开展系统性植物性状测定提供了较为理想的平台,伴随着“群落性状”概念的提出和推导方法的逐步成熟,将有助于建立传统性状研究与宏观生态学的桥梁。何念鹏等利用热带-寒温带9个典型森林生态系统详细的调查数据,并尝试性地利用生物量加权法将叶片气孔从器官推导至群落水平,并发现群落水平的气孔性状呈现出明显的纬度格局,且与生态系统总初级生产力和水分利用效率显著相关[18,35-36],提供了天然森林群落性状与功能联系的直接证据。Zhang等[53]利用该方法获得了中国东部8个森林叶-枝-干-根和土壤的C、N、P含量的加权平均值,并发现其结果与前期收集数据(数据直接平均法)所获得结果差异显著。过去几年基于数据直接平均法或生物量相对加权法的研究较多,但真正基于群落生物量加权法的研究还仅在少数几个站点实现;尤其是在如何实现由点至面拓展、天然群落性状-功能定量评估、群落性状与快速发展的宏观生态学观测技术的联系等方面,还有待深入研究。
5 群落性状的应用前景与展望
(基于单位土地面积的)群落性状作为一个新概念,其指标体系和应用将会是一个逐渐深入和拓展的过程。随着调查数据的逐渐丰富,它在如下几方面具有较好的应用前景:1)群落性状将有利于更好地探讨自然群落的性状与功能间的定量。在传统研究中,大多数研究都是在物种水平测试植物性状并探讨性状与功能的关系,并将这种物种水平的定量关系直接应用于自然群落(物种水平=群落水平);在复杂的自然群落中,这种简单替换的定量关系的可信度还有待实测数据的进一步证实。2)群落性状将有利于揭示全球变化对生态系统结构和功能的影响。由于群落性状能与通量观测、近地面遥感和卫星遥感的观测数据在单位土地面积上匹配,使性状研究可更好地结合或服务于相关的新技术,即推动新技术的发展,又为性状研究提供新的生长点。当前,全球变化的生态效应多是在生态系统水平进行评估的;在利用N∶P比变化来探讨养分限制性或未来氮磷沉降不对称的生态效应时,如果能用群落水平的N∶P比(群落内不同物种长期适应与权衡的整体表现形式),其评估结果应该可以更准确。3)为生态模型提供更多和更准确的关键参数,提高模型模拟精度。以C∶N比为例,由于群落水平数据难以获得,大多数生态模型均使用个别物种数据或少数物种算数平均会值来替代,如能提供更精确的群落水平C∶N,应该可以提高相关模型的预测精度。4)群落性状的提出能促使更多植被性状纳入生态模型,为开发新一代模型奠定坚实基础[60]。目前,许多学者已经意识到纳入更多性状参数是提高预测精度的有效措施,随着群落性状数据的逐步增多,未来一定会有更多性状参数被纳入模型以提高模型精度。
如果未来能进一步发展生态系统性状,以单位土地面积为基础,建立植物群落性状、动物群落性状、土壤微生物群落性状、土壤属性和气候特征的联系,将有利于从新视角探讨生态系统内部生物和非生物间的有机联系。在实际调查过程中,许多学者在特定地点或多个地点,都开展了系统的群落结构与性状调查,但由于传统概念与个人理解的差异,这类数据或群落性状很少被公布并积累下来,这是性状研究的巨大损失。鉴于此,本文探讨了群落性状的定义及其潜在应用,希望能引起国内同行对性状研究概念与方法的思考,推动性状研究从物种水平向群落水平的跨越,甚至拓展到区域或全球水平。同时,希望新的概念体系能逐步将传统性状与高速发展的新技术(定量遥感、通量观测、生态模型)相结合,更好地服务于区域生态环境问题的解决,拓展性状研究的新生长点。
猜你喜欢
四川蚕业(2022年2期)2022-11-19
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
世界科学技术-中医药现代化(2020年2期)2020-07-25
河南科学(2020年3期)2020-06-02
科学大众(中学)(2019年3期)2019-05-17
汽车观察(2018年10期)2018-11-06
临床医药文献杂志(电子版)(2017年11期)2017-05-17
科技知识动漫(2017年1期)2017-02-06
草业科学(2015年9期)2015-12-11
