
烤烟CYP82E10基因EMS突变体筛选及其尼古丁转化率分析
2018-11-10 02:01李文正李梅云吴兴富王丙武高玉龙隋学艺赵璐宋中邦
中国烟草学报 2018年5期
李文正,李梅云,吴兴富,王丙武,高玉龙,隋学艺,赵璐,宋中邦
云南省烟草农业科学研究院/烟草行业烟草生物技术育种重点实验室/国家烟草基因工程研究中心,昆明 650021
生物碱是烟属植物特征性次级代谢物,栽培烟草中生物碱主要为尼古丁(nicotine)、去甲基尼古丁(nornicotine)、假木贼碱(anabasine)和新烟草碱(anatabine)。尼古丁是首要的生物碱,占生物碱总含量的90%~95%,去甲基尼古丁由尼古丁经过去甲基反应生成(该过程通常被称为尼古丁转化),含量通常低于总生物碱的5%。去甲基尼古丁在烟叶烘烤、调制、贮存阶段能够被亚硝化生产NNN,后者在许多动物实验中被证明与许多类型癌症关联,因此被WHO认定为强致癌物[1-3]。此外,有研究表明去甲基尼古丁本身也能直接诱导吸烟者血浆中蛋白的异常糖基化,影响类固醇类药物的药效和毒性[4]。因此,抑制尼古丁转化进而降低去甲基尼古丁和NNN含量,是烟草减害的重要方向之一。
白肋烟去甲基尼古丁含量远高于烤烟,因此尼古丁转化研究主要集中于白肋烟。白肋烟中存在一个目前仍未知的不稳定基因座,导致去甲基尼古丁含量很低的白肋烟亲代(非转化株)也能产生去甲基尼古丁含量很高的子代(高转化株),高转化株中大部分尼古丁转化生成去甲基尼古丁,比率甚至高达98%[5-6]。在生产实际中,每年需要投入大量的时间和经费淘汰尼古丁转化率超过3%的单株,以控制白肋烟去甲基尼古丁含量[7]。尼古丁去甲基酶负责催化尼古丁转化成去甲基尼古丁的反应,迄今为止共有4个功能基因被克隆,分别为CYP82E4、CYP82E5、CYP82E10、CYP82E21[8-11]。CYP82E21基 因 虽 能编码具有活性的尼古丁去甲基酶,但仅在子房中表达,对叶片去甲基尼古丁含量影响不大,研究焦点主要集中在前3者[10]。用甲基磺酸乙酯(EMS)化学诱变策略获得的CYP82E4、CYP82E5、CYP82E10均发生突变的三突白肋烟材料,尼古丁转化率仅为0.55%,和RNAi转基因植株相当[9,12-13]。Li等人针对上述3个基因的突变序列设计了CAPS和dCAPS标记进行分子标记辅助选择,加快低NNN品种培育[14]。目前,美国已获得众多低NNN含量的白肋烟、深色烟、烤烟改良品种,卷烟工业研究表明低NNN烟叶能够特异性降低混合型卷烟填料和主流烟气的NNN含量[15]。
烤烟是中式卷烟的主要原料,降低其去甲基尼古丁含量进而降低NNN含量对中式卷烟减害有重要意义[16]。但迄今为止关于烤烟去甲基尼古丁合成的研究较少,烤烟与白肋烟的合成机制是否一致目前尚无文献报道。本研究借鉴白肋烟类似的策略,在烤烟品种云烟87的EMS突变体库中筛选CYP82E2家族基因的突变体,获得错义突变体M594和M271,生物碱数据分析表明两个突变体的尼古丁转化率与对照相比显著下降,这与白肋烟CYP82E10基因突变几乎不影响尼古丁转化率不同,表明烤烟与白肋烟的去甲基尼古丁合成机制不同。
1 材料与方法
1.1 EMS突变体库创制
普通烟草云烟87种子参考已发表文献的操作进行EMS处理[9],处理后种子作为M1代突变体种子进行田间种植,套袋自交收获M2代种子。大田种植约2200份M2代EMS突变体植株,采集叶片提取基因组,每8份DNA混样,构成8倍混合池,保存于96孔板,用于后续突变体筛选。
1.2 Tilling筛选条件
采用Primer 3软件针对CYP82E10(NCBI登录号:HM802352)基因第一个外显子设计特异引物进行Tilling筛选,正向引物为CYP82E10_Tilling_F:GTCAAATACCACCTCTTAATAGTAA,反 向 引 物 为CYP82E10_Tilling_R:AAAAGTCCCTATTGGTAGGAAGTGC,PCR产物大小为1335 bp。扩增体系总体积为10 μL(TOYOBO预混液),其中包含20 ng DNA模板,0.8 mmol/L dNTP,1 单位 KOD FX Taq polymerase,320 nmol/L引物, 5 μL 2×KOD FX buffer。扩增采用touchdown PCR,95 ℃预变性 2 min,98 ℃变性10 s,64 ℃退火30 s,68 ℃延伸1 min,退火温度每个循环依次降低1 ℃,共4个循环。随后98 ℃变性10 s,60 ℃退火30 s,68 ℃延伸1 min,共45个循环。最后68 ℃延伸5 min,4 ℃保持。
CEL1 酶切体系体积为 6 μL,包含 0.2 μL CEL1(1 unit), 1.2 μL 10× CEL1 buffer, 2 μL PCR 产 物 和2.6 μL 去 离 子 水。 采 用 AdvanCE FS96 (Advanced Analytical Technologies, USA)进行毛细管电泳。
1.3 突变位点Sanger测序验证及功能预测
采集突变株幼嫩叶片组织,用DNA提取试剂盒(QIAGEN)提取基因组DNA。以基因组DNA为模板,采用CYP82E10_Tilling_F和CYP82E10_Tilling_R引物进行扩增,PCR产物回收后直接进行测序(Takara)。通过在线软件PROVEAN Protein(http://provean.jcvi.org/seq_submit.php)对突变位点的功能损失进行预测,PROVEAN得分小于-2.5表明突变会影响蛋白功能,大于-2.5表明突变对蛋白功能影响不大。利用SWISS-MODEL(https://swissmodel.expasy.org/)进行蛋白同源建模,分析高级结构。
1.4 田间试验设计及生物碱含量测定
2016—2017年,在云南省烟草农业科学研究院研和试验基地(云南玉溪)田间分别种植M4和M5代突变体和对照材料,采用随机区组设计,每个材料设3个重复小区,每个小区栽烟约30株。种植密度、栽培调制技术及田间管理措施与当地优质烟叶生产相同。各小区现蕾打顶,一周后一次性采集整株叶片60 ℃烘干,粉碎过60目筛待测。
内标喹啉、去甲基尼古丁购自Sigma-Aldrich公司,尼古丁购自TRC公司,检测仪器为Bruker 450GC-300MS气相色谱-串联质谱仪(Bruker)。准确称取烟样0.5 g于50 mL离心管中,加入5 mL 10%的NaOH溶液,摇匀,浸泡15 min,加入20 mL含内标的萃取液,超声60 min,在离心机上5000 rpm/min离心5 min,取2 mL下层二氯甲烷清液过装有2 g无水硫酸钠的微孔过滤器后,用气相色谱-串联质谱仪分析[17]。
2 结果与分析
2.1 云烟87突变体库创制
利用EMS诱变建立烤烟品种云烟87的突变体库,大田种植约2200份M2代EMS突变体植株(图1A)。采集叶片提取基因组DNA,并将所有样品DNA浓度稀释至40 ng/ μL(图1B),最终建立了包含1842份M2代云烟87突变体的DNA库。

图1 EMS突变体库构建Fig.1 Development of EMS mutant population
2.2 CYP82E10基因突变体筛选
烟草CYP82E10基因由2个外显子和1个内含子组成(图2A),已被证明具有尼古丁去甲基酶活性。利用Tilling技术在突变体库中筛选该基因发生突变的单株,共获得11个突变株(图2B),其中突变单株M594发生L115F突变,M271单株发生P129L突变(表1)。PROVEAN Protein分析表明M594单株突变PROVEAN得分为-3.06,M271单株突变PROVEAN得分为-3.16,2个突变均可能影响蛋白功能(表1)。构建CYP82E10野生型及突变蛋白的三维结构模型,发现M594单株中L115F突变发生在第4个α螺旋,M271单株中P129L突变发生在第5个α螺旋,2个突变均对蛋白三维结构产生影响(图3A)。有预测表明CYP82E2家族蛋白中存在6个保守的底物识别结构域[9,18],L115F和P129L均位于第1个识别结构域,因此极有可能影响酶与底物的结合(图3B)。提取M3代突变单株叶片基因组DNA,PCR扩增后进行Sanger测序验证,M594和M271单株中均发现野生型、杂合突变、纯合突变三种基因型(图4)。将纯合突变单株套袋自交获得M4代种子,用于后续表型分析。

图2 CYP82E10基因突变体筛选Fig.2 Identification of CYP82E10 mutants

表1 云烟87突变体库中CYP82E10基因突变体统计Tab.1 Information of CYP82E10 mutants screened from EMS population of N.tabacum var.Yunyan87

图3 突变蛋白结构及功能预测Fig.3 Structural and functional prediction of mutated protein
2.3 突变体材料尼古丁转化率分析
田间种植突变材料和云烟87,随机挑取10株进行Sanger测序,结果表明M271和M594中CYP82E10基因突变均已纯合,CYP82E4和CYP82E5基因未发生突变。烟株成熟后打顶,一周后采集整株叶片烘干分析尼古丁和去甲基尼古丁含量,并计算尼古丁转化率(图5)。M4代植株中,M594材料尼古丁转化率是云烟87的45%,M271材料尼古丁转化率是云烟87的38%。M5代植株中,M594材料尼古丁转化率是云烟87的34%,M271材料尼古丁转化率是云烟87的45%。这表明烤烟中CYP82E10蛋白的L115F,P129L突变能够使其尼古丁去甲基酶活性减弱,造成烟叶中尼古丁转化率显著下降。M4、M5两代突变体材料的尼古丁转化率均比云烟87显著降低,表明该性状能够稳定遗传。

图4 突变体CYP82E10基因测序峰图Fig.4 Sequence polymorphisms between mutant and wild type alleles in CYP82E10
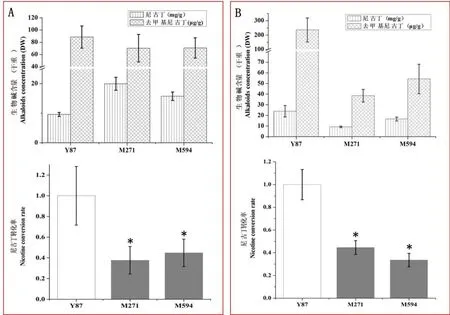
图5 突变体材料生物碱含量及尼古丁转化率Fig.5 Alkaloids concentration and nicotine conversion rate in mutant lines
3 讨论与结论
烟草中CYP82E2家族基因编码尼古丁去甲基酶,催化尼古丁转化生成去甲基尼古丁,是烟叶中致癌物NNN的前体物质。国际上该领域的研究主要集中在美国北卡州立大学、肯塔基大学等机构,主要研究材料是白肋烟。我国中式卷烟主要原料是烤烟,但目前尚未培育出低尼古丁转化率烤烟材料和品种,烤烟尼古丁转化机制也不清晰。本研究利用EMS处理获得云烟87突变体库,通过Tilling技术筛选获得了CYP82E10基因的两个错义突变体M594和M271,并进行尼古丁转化率分析。
CYP82E4在白肋烟中是主效基因,其EMS终止突变株尼古丁转化率为2.2%,远低于对照材料(>60%)[9,12]。CYP82E5、CYP82E10基 因 单 突 及双突材料的尼古丁转化率与对照无显著差异,但三突材料尼古丁转化率为0.55%,比CYP82E4单突更低,和RNAi转基因植株相当[9,13]。本研究中烤烟CYP82E10基因的两个错义突变体M594和M271的尼古丁转化率在连续2年的田间试验中均比对照显著降低,这表明烤烟和白肋烟的尼古丁转化机制不完全一致。烤烟CYP82E4基因不表达,主要由CYP82E5和CYP82E10编码的蛋白参与尼古丁的转化,而CYP82E10突变体尼古丁转化率显著下降,表明CYP82E5无法互补CYP82E10的功能。其次,本研究CYP82E10基因并非发生无义突变但造成其催化活性下降或丧失,说明CYP82E10蛋白115位的亮氨酸和129位的脯氨酸对其立体构象非常关键,突变可能造成底物尼古丁无法被结合。
NNN被世界卫生组织认定为强致癌物,是传统卷烟的主要危害成分之一,降低其烟叶含量是烟草减害的重要目标。此外,近年来随着加热不燃烧卷烟迅猛发展,由燃烧引起的有害成分大幅降低,烟叶内源NNN,NNK等TSNA物质的危害将更加突出。本研究获得的低尼古丁转化率材料中去甲基尼古丁含量显著下降,目前正作为育种材料进行自交和回交,培育低NNN烤烟新品种,为卷烟工业提供低危害烟叶原料。
猜你喜欢
亚热带农业研究(2022年1期)2022-08-08
农业科技通讯(2021年1期)2021-03-06
大众健康(2020年11期)2020-11-13
农业技术与装备(2020年10期)2020-11-03
云南农业(2020年5期)2020-06-12
中国农业科技导报(2020年3期)2020-03-15
中国食品学报(2019年12期)2019-01-13
诗歌月刊(2014年3期)2015-04-14
传奇故事(破茧成蝶)(2015年8期)2015-02-28
祝您健康(2000年5期)2000-12-29
