
山西省运城市番茄黄化曲叶病毒的分子鉴定
2018-11-08 06:05郝浩永滕红梅许瑞祥
江苏农业科学 2018年19期
郝浩永, 滕红梅, 刘 缙, 许瑞祥, 王 新
(1.运城学院理科实验中心,山西运城 044000; 2.运城学院生命科学系,山西运城 044000)
番茄黄化曲叶病毒(tomato yellow leaf curl virus,简称TYLCV)是双生病毒科菜豆金色花叶病毒属(Begomovirus)病毒,主要依靠烟粉虱进行传播。大多数菜豆金色花叶病毒属病毒的基因组由双组分组成,即DNA-A、DNA-B,大小为2.5~2.8 kbp,少数病毒为单组分,仅含有DNA-A[1-2]。目前国内发现的TYLCV大多为DNA-A,其基因组大小约为 2.8 kbp,有些单组分病毒还伴随有卫星DNA[1]。TYLCV于1964年在以色列被发现并正式命名,随后在短短几十年时间内,迅速在世界各地蔓延,给番茄等农作物生产造成了巨大的损失[3]。2002年番茄黄化曲叶病毒病传入我国上海[4],之后在我国20多个省、市相继发生,给番茄生产带来了严重的威胁[5-7]。2010年,全国番茄种植面积超过20万hm2,经济损失达数十亿元[8]。
有报道称,2010年山西省运城市局部番茄田已出现了番茄黄化曲叶病毒病的危害[9-10]。2016年,笔者在运城学院新校区及周边多处番茄种植地发现疑似感染番茄黄化曲叶病毒的植株,主要表现为叶片边缘黄化、卷曲、植株矮化等症状。进一步调查发现,番茄黄化曲叶病毒病在运城市的多个番茄种植区都有发生,而且危害较严重,尤其是在夏季和秋季。由于运城市天气干旱炎热的时期较长,适合该病的传播媒介——烟粉虱的生长和繁殖,因此对当地的番茄生产造成了极大的经济损失。近年来,山西省运城市大规模烟粉虱暴发活动频繁出现,2017年9月上旬运城市郊区甚至城区均出现大量烟粉虱。本研究的目的在于通过采集感病番茄植株样品,对引起该病的TYLCV的DNA-A组分进行检测和鉴定,以明确其DNA-A组分的基因组结构,了解发病原因和症状表现,以期为当地防治番茄黄化曲叶病毒病提供一定依据。
1 材料与方法
1.1 感病样品采集
疑似感染番茄黄化曲叶病毒的番茄病株于2016年8月采自山西省运城学院新校区,病株表现矮化、叶片黄化且严重上卷、落花等TYLCV侵染的典型症状,采集新鲜叶片直接用于总DNA的提取,或保存于-70 ℃备用。以健康植株作为对照。
1.2 番茄叶片总DNA的提取
采用十六烷基三甲基溴化铵法提取番茄叶片的总DNA,于-20 ℃保存备用[11]。
1.3 引物设计与合成
参照GenBank中已发表的TYLCV的序列,通过同源性比对分析,并参考文献[12],根据开放阅读框(open reading frame,简称ORF)AC1和AV2保守区域,设计2对引物(P1F/P1R、P2F/P2R)。引物P1F/P1R用于扩增约2 450 bp的DNA片段,引物P2F/P2R用于扩增约550 bp的DNA片段。2个片段的重叠部分用于拼接,以获得病毒DNA-A的全长序列。2对引物的序列如表1所示。引物由生工生物工程(上海)股份有限公司合成。利用简并引物PCRc1/PBLv2040扩增可能存在的双生病毒DNA-B组分[13],利用通用引物β01/β02、UNA101/UNA102分别扩增可能存在的双生病毒伴随卫星DNAβ与DNA1全长序列[14-15]。
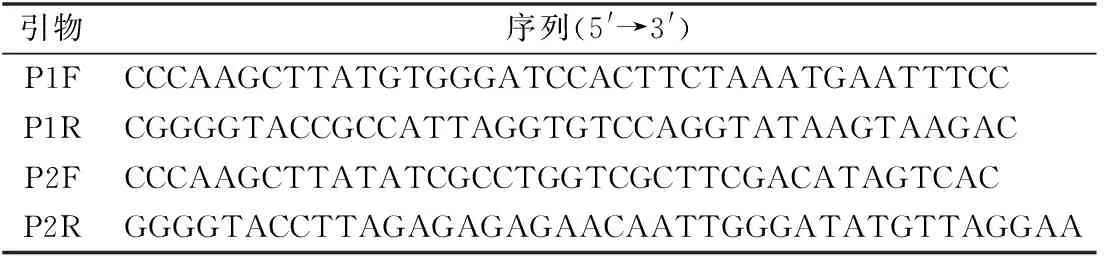
表1 引物序列信息
1.4 目的片段的扩增和序列测定
以提取的番茄植株叶片的总DNA作为模板进行PCR扩增,引物P1F/P1R和P2F/P2R所用的PCR反应体系相同,反应条件不同。PCR反应体系:1 μL DNA模板,2 μLTaqPlus DNA聚合酶(2.5 U/μL),5 μL 10×TaqPlus Buffer,引物(100 μmol/L)各1 μL,10 μL dNTP Mixture(2.5 μmol/L),加入ddH2O补足至50 μL。引物P1F/P1R的反应条件:94 ℃ 5 min;94 ℃ 30 s,50 ℃ 30 s,7 2 ℃ 2 min,15个循环;94 ℃ 30 s,58 ℃ 30 s,72 ℃ 2 min,25个循环;72 ℃ 10 min。引物P2F/P2R的反应条件:94 ℃ 5 min;94 ℃ 30 s,50 ℃ 30 s,72 ℃ 30s,35个循环;72 ℃ 5 min。
DNA-B、DNAβ、DNA1的PCR检测参照文献[13-15]完成。PCR扩增完成后送至生工生物工程(上海)股份有限公司进行测序。
1.5 序列拼接和分析
测序结果使用DNA Star Lasergene 7.1软件中的Seqman程序进行拼接,用MegAlign程序进行序列相似性比较。用NCBI中的ORF finder程序查找ORF,采用软件MEGA 6.0中的Clustal W方法进行多序列比对和邻接(neighbor-joining)法构建系统进化树,并进行1 000次Bootstrap检验。
2 结果与分析
2.1 PCR检测结果
以病株样品和健康番茄样品提取的DNA为模板,利用引物P1F/P1R在病株样品中扩增出大小约为2 450 bp的条带,在健康植株中未扩增出(图1);利用引物P2F/P2R进行PCR扩增,从病株中扩增出大小约为550 bp的条带,在健康植株中未扩增出(图2),与相关文献报道[12]一致,确认本研究所采病株感染了番茄黄化曲叶病毒。DNA-B、DNAβ、DNA1在样品中均未检测到,推测感染的番茄黄化曲叶病毒可能是单组分病毒,且无卫星DNA伴随。


2.2 病毒DNA-A的全长序列分析及其基因组结构特征
将长短2条目的条带的测序结果用软件DNA Star Lasergene 7.1进行拼接,最终得到1个长2 781 nt的单链环状DNA,即DNA-A组分,将其命名为SXYC-X4,GenBank序列号为KY499720。SXYC-X4具有菜豆金色花叶病毒属病毒基因组的典型特征:有6个ORF,其中AV1、AV2位于病毒链上,AC1、AC2、AC3、AC4位于其互补链上;AV1(308~1 084 nt)编码病毒的外壳蛋白(coat protein,简称CP),AV2(148~498 nt)编码RNA沉默抑制子和运动蛋白,AC1(1 542~2 615 nt)编码复制相关蛋白(replication-associated protein,简称Rep),AC2(1 226~1 633 nt)编码转录激活蛋白(transcriptional activator protein,简称TrAP),AC3(1 081~1 485 nt)编码复制增强蛋白(replication enhancer protein,简称Ren),AC4(2 171~2 464 nt)编码复制或转录调控因子;在AC1和AV2之间有1个313 nt的基因间隔区(intergenic region,简称IR),为非编码区,该区含有保守的九核苷酸序列TAATATT/AC(2 775~2 nt)的茎环结构、TATA box(2 675~2 678 nt)[16]及正向重复序列ATCGGTGT(2 653~2 660 nt,2 661~2 668 nt)。
2.3 病毒DNA-A与其他TYLCV的 DNA-A的同源性分析
将SXYC-X4的全长序列通过Blast进行比对分析,结果显示,其与山东省潍坊市的病毒分离物TYLCV-SDWF-L7(KC999850)、河南省焦作市的病毒分离物TYLCV-HNJZ-19(KX034543)的同源性较高,均为99.5%。在AV1、AV2、AC1、AC2、AC3、AC4核苷酸序列同源性方面,SXYC-X4与TYLCV-SDWF-L7相比较同源性分别为99.2%、99.7%、99.7%、99.0%、99.5%、99.3%,SXYC-X4与TYLCV-HNJZ-19相比较同源性分别为99.1%、99.7%、99.5%、99.3%、99.5%、99.3%。从GenBank上选取25个具有代表性的双生病毒序列进行比对,由表2可知,SXYC-X4与这25种双生病毒的DNA-A全序列同源性为76.8%~99.5%,IR区为61.5%~99.7%。推导AV1、AV2、AC1、AC2、AC3、AC4编码的蛋白质氨基酸序列的同源性分别为77.3%~99.2%、72.2%~100.0%、69.5%~99.4%、62.7%~98.5%、70.1%~99.3%、32.9%~97.9%,其中AV1氨基酸序列的变异最小,AC4氨基酸序列的变异最大。
2.4 病毒DNA-A与其他双生病毒的系统进化分析
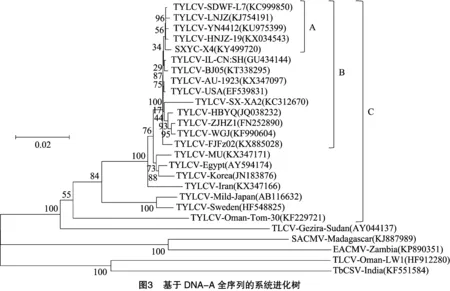
为进一步明确SXYC-X4的系统进化关系及分类地位,将SXYC-X4与表2中25个双生病毒的核苷酸序列构建系统进化树。由图3可知,SXYC-X4与国内的TYLCV-SDWF-L7、TYLCV-HNJZ-19、TYLCV-YN4412、TYLCV-LNJZ等的亲缘关系较近,形成1个独立的分支A,再与澳大利亚的TYLCV-AU-1923、美国的TYLCV-USA及国内其他分离物形成1个较大的分支B,与所有番茄黄化曲叶病毒则形成1个更大的分支C;而与番茄曲叶病毒、南非木薯花叶病毒、东非木薯花叶病毒、烟草曲茎病毒等的亲缘关系相对较远。据报道,通常双生病毒IR区的变异最大[1,7,12,17],而本研究中AC4氨基酸序列相似性变异大于IR区(表1),SXYC-X4与TbCSV-India的同源性高于属TYLCV的TYLCV-Oman-Tom-30、TYLCV-Sweden、TYLCV-Mild-Japan,然后分别基于IR区和AC4氨基酸序列作系统进化分析。由图4可知,基于IR区,SXYC-X4与TYLCV-AU-1923、TYLCV-USA、TYLCV-LNJZ等形成1个分支A,再与除TYLCV-Mild-Japan、TYLCV-Oman-Tom-30、TYLCV-Sweden之外的TYLCV形成1个较大的分支B。由图5可知,基于AC4氨基酸序列,SXYC-X4与大部分的TYLCV形成1个分支A,所有TYLCV与TbCSV-India形成1个更大的分支B。
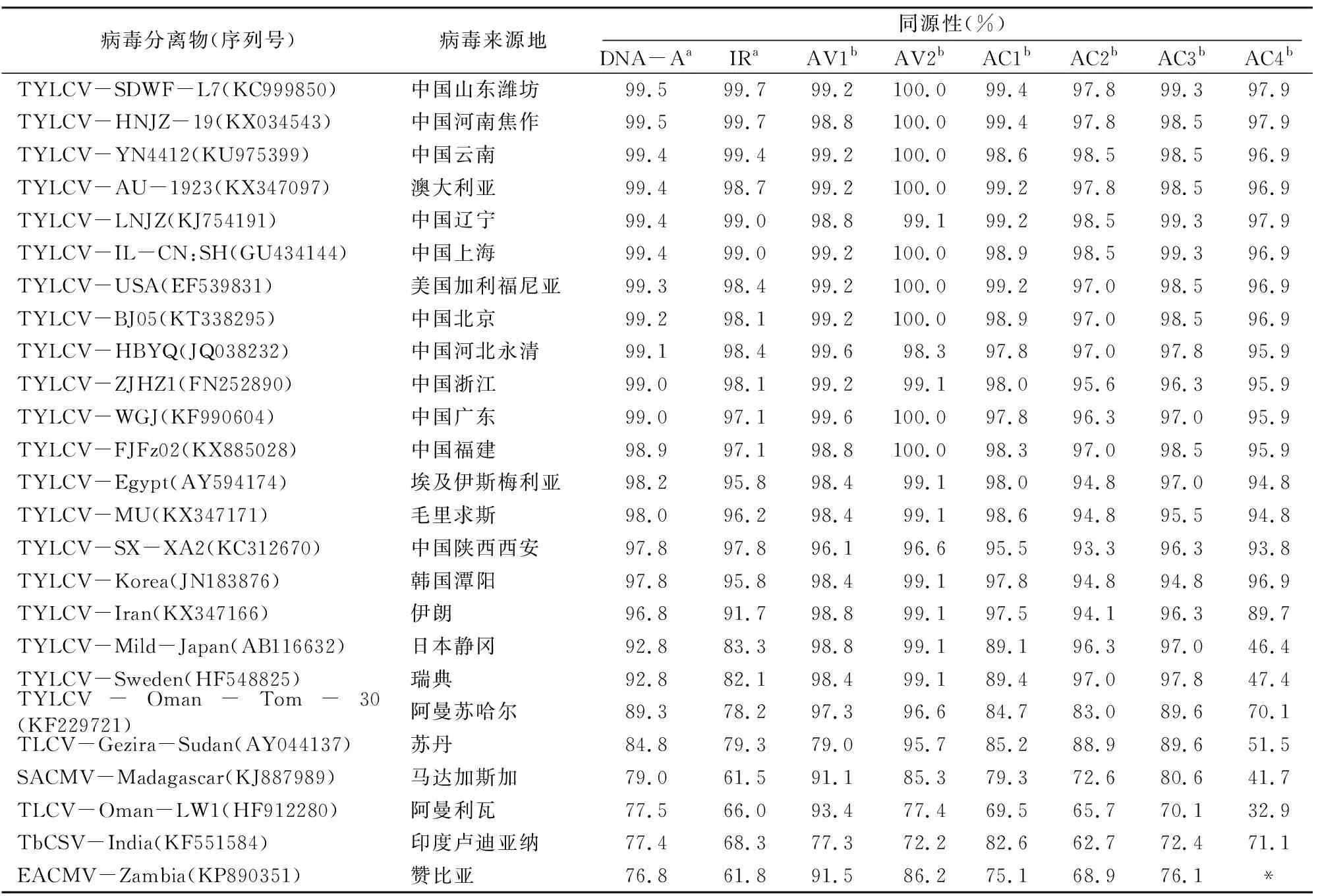
表2 SXYC-X4与25种菜豆金色花叶病毒属病毒DNA-A及ORF的序列比对结果
注:TLCV代表烟草曲叶病毒;SACMV代表南非木薯花叶病毒;TbCSV代表烟草曲茎病毒;EACMV代表东非木薯花叶病毒;a代表核苷酸序列;b代表氨基酸序列;*代表无可比对区域。
3 讨论与结论
通过对山西省运城市病毒分离物SXYC-X4基因组全序列进行分析,结果表明其具有双生病毒科菜豆金色花叶病毒属单组分病毒基因组的典型特征,与TYLCV-SDWF-L7、TYLCV-HNJZ-19的同源性较高,均为99.5%。与SXYC-X4全基因组核苷酸序列同源性大于89%的病毒均为TYLCV,而与同为双生病毒的番茄曲叶病毒、南非木薯花叶病毒、东非木薯花叶病毒、烟草曲茎病毒等的同源性均小于89%。根据双生病毒科病毒分类原则[17-20],SXYC-X4应属于TYLCV的一个株系。通过构建DNA-A全序列系统进化树发现,SXYC-X4与属TYLCV-Isreal株系[21]山东、河南、云南、辽宁等地区的分离物聚在一支,亲缘关系最近(图3),因此笔者认为,SXYC-X4属TYLCV-Isreal株系,这也是目前分布最广的一个株系[22]。根据目前已报道的资料[14-15,23-28]来看,单组分双生病毒通常伴有卫星DNA,但本研究未检测到,SXYC-X4是否伴有卫星DNA仍须进一步研究确认。在图3分支B中,仅有TYLCV-AU-1923、TYLCV-USA为国外分离物,且分离的时间较早(分别为2006年、2007年),其余皆为国内分离物,分离时间较晚(2007—2016年),推测可能由于种质资源交流及烟粉虱扩散致使TYLCV在山西省运城地区蔓延。在双生病毒的各ORF中,最保守的是AV1基因,通常同一地区双生病毒的AV1同源性相当高[29-31]。本研究中,SXYC-X4与来自世界各地25种双生病毒的各ORF编码的蛋白中,AV1编码的CP氨基酸序列的变异最小(77.3%~99.2%),这进一步证实这种推测。

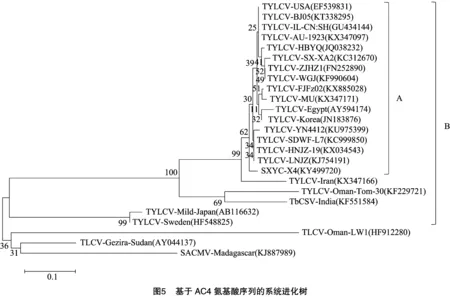
TbCSV也可感染番茄[32],TYLCV与TbCSV可复合侵染烟草、番茄等作物,不同双生病毒之间的重组经常发生[33-34]。RNA沉默是指多种真核生物体内发现的一种基于RNA水平的高度保守的特异性降解机制。RNA沉默的一种重要效应是防御病毒侵染。崔晓峰等发现,中国番茄黄化曲叶病毒Y10分离物在侵染本氏烟等寄主植物时,并未诱导明显的症状,而烟草曲茎病毒Y35分离物单独侵染则诱导曲叶等症状,病毒Y35的AC4蛋白是一个比Y10的AC4蛋白更强的RNA沉默抑制子,证明菜豆金色花叶病毒属病毒的AC4蛋白与病毒致病性相关[35]。何自福等研究发现,来自同一地区的ToLCGDV-[G3]和ToLCGDV-[G2]同属菜豆金色花叶病毒属的成员,亲缘关系最近,全基因组同源率为87.9%[29]。本研究发现,AC4氨基酸序列的变异最大(32.9%~97.9%),TYLCV与TbCSV-India的进化关系在DNA-A全序列水平较远,在IR区水平稍近,在AC4氨基酸序列水平最近,由此推测,自然界中TYLCV与TbCSV在AC4序列处可能已经发生了广泛的重组,TYLCV有可能会获得更强的致病性,应提高警惕。
番茄黄化曲叶病可由多种双生病毒侵染引起,TYLCV是主要毒源,此病症可能还由其他病毒复合侵染引起[21,36-37]。本研究目前只检测了TYLCV,且发现SXYC-X4与TbCSV在AC4氨基酸序列上的同源性高于部分TYLCV,病株是否同时感染了其他病毒,仍须进一步检测。
本研究从分子水平上鉴定了山西省运城地区引起番茄黄化曲叶病的病原,明确了病毒分离物DNA-A全长序列,分析了其在双生病毒科中的分类地位及系统进化关系,为进一步利用全长序列构建侵染性克隆奠定了基础,对山西省运城市当地番茄黄化曲叶病毒病的防治和番茄抗病育种工作具有重要的参考价值。
猜你喜欢
中国蔬菜(2021年5期)2021-06-15
现代装饰(2018年3期)2018-05-22
中国蔬菜(2016年8期)2017-01-15
中国果菜(2016年9期)2016-03-01
火花(2016年7期)2016-02-27
中国糖料(2015年6期)2015-11-25
中国烟草学报(2015年3期)2015-01-25
中国艺术时空(2014年2期)2014-02-28
新疆农垦科技(2014年5期)2014-02-28
长江蔬菜(2013年21期)2013-04-02
