
濒危植物景宁玉兰的萌条形态特征分析
2018-11-07 06:48:16杜有新吴伟建季志利王军峰江丽荣刘跃钧
生态科学 2018年5期
杜有新, 吴伟建, 季志利, 王军峰, 江丽荣, 刘跃钧, *
濒危植物景宁玉兰的萌条形态特征分析
杜有新1, 吴伟建2, 季志利3, 王军峰1, 江丽荣3, 刘跃钧1, *
1. 丽水市林业科学研究院, 浙江 丽水 323000 2. 松阳县林业局, 浙江 松阳 323400 3. 丽水莲都生态林场, 浙江 丽水 323000
为了解景宁玉兰萌条发育特点, 选择了灌丛(SH)、黄山松林(PTF)、落叶阔叶林(DBF)、杉木林(CLF)和林缘(FE)等5种坡向一致的生境类型, 通过样地调查, 比较了5种不同生境上景宁玉兰萌条大小、高度及枝系等性状。结果表明: (1)景宁玉兰在黄山松林下的分布密度最高, 但5种生境上的景宁玉兰每丛的萌条数量没有差异, 每丛最大萌条高度与每丛萌条数没有关系。(2)在落叶阔叶林下的萌枝基径显著小于其他生境(< 0.05), 黄山松林下一级枝粗度和长度显著小于其他生境(< 0.05); 灌丛中的总分枝率和逐步分枝率最高, 灌丛和林缘生境的逐步分枝率(SBR2:3)高于其他生境。(3)在所调查的景宁木兰居群中, 大于2根以上萌条的植株达82.5%, 说明萌条更新在种群繁衍中发挥着重要作用。本研究表明, 景宁玉兰萌生性特征主要受其内在生物学特性所控制, 而萌条形态则明显受到所处环境条件的影响。
景宁玉兰; 萌条形态; 分枝率
1 前言
景宁玉兰(P. L. Chiu et Z. H. Chen) 属浙江特有种[1-2], 为国家二级保护的濒危物种[3-4], 具有较高的科学价值和园林观赏价值[5]。景宁玉兰集中分布于浙江南部的浙江省丽水市的景宁县草鱼塘林场、松阳县牛头山地区、莲都区峰源乡和温州市乐清雁荡山, 其原生环境相似, 由于人类活动的干扰及自然环境变化导致的生境破碎化, 现代分布格局呈分散性特征[6], 主要生长于海拔900 m以上的落叶阔叶林、杉木林、黄山松林、林缘沟边和灌丛中[7]。
景宁玉兰为浅根性落叶灌木, 喜阴凉湿润、水分充足且土壤肥沃的生境, 对环境变化较为敏感[7-8]。在自然环境下, 景宁玉兰自然结实率不到1%[9], 但其萌生性强, 茎基部具有很强的萌芽能力[2], 实生苗定植后往往萌发数株萌蘖[10], 根颈萌条和残桩萌条[11]现象在野生环境下普遍存在, 这也是其对资源与环境变化的一种适应性响应[12]。之前对景宁玉兰生态生物学特性[2]、种群分布[6-7]、花粉活力[8-9]、无性繁殖[10]及属内亲缘关系[13]等方面进行了较多研究, 但未见有关不同生境下景宁玉兰萌条形态特征方面的报道。萌条形态可以反映萌生性树种的生长发育状况及其对环境的适应策略, 本文通过调查分析不同生境下景宁玉兰萌条的形态特征, 以期了解景宁玉兰萌条更新特征及其与生境的关系, 为科学保护合理保护其种质资源提供参考。
2 研究地与方法
2.1 研究地概况
本调查选取景宁玉兰代表性的3个分布点即丽水市松阳县大东坝镇灯塔村牛头山地带、丽水市莲都区峰源乡尤源村和葑垟村, 3个分布点相距20—60 km。分布点皆处于山体北坡, 包含了景宁玉兰自然生长环境的各种植被类型, 除了松阳牛头山海拔720 m处生长2丛外, 景宁玉兰主要分布于海拔950 m以上的山地。丽水地理位置为北纬27°25′—28°27′, 东经118°42′—120°26′之间, 属中亚热带季风气候, 温暖湿润, 四季分明, 年平均气温为16.9—18.5 ℃, 7月份平均气温为27.8—29.2 ℃, 极端最高温41 ℃, 1月份平均气温为5.0—7.8 ℃, 极端最低气温-13.1 ℃; 年平均相对湿度各地在75%—87%之间, 年均降水量1437—2069.5 mm[14]。景宁玉兰自然分布区内土壤为山地黄壤和黄棕壤, 土层厚度30—120 cm, pH值4.8—5.5[2]。
2.2 样地设置
于2016年2—3月, 根据景宁玉兰生长的实际环境, 把景宁玉兰生境分为5种植被类型, 分别为灌丛(SH)、黄山松()林(PTF)、落叶阔叶林(DBF)、杉木()林(CLF)及林缘沟边(FE)。
根据景宁玉兰分布状况, 在每种生境上分别设置1—3个典型样地, 每个样地面积10 m×20 m, 葑垟村共设置了5个样地, 尤源村共设置了4个样地。在松阳县牛头山地区, 景宁玉兰分布于海拔716—1300 m之间, 结合景宁玉兰居群规模, 在海拔720 m、1100 m和1250 m处分别设置1、2和3个样地, 其中海拔720 m处只发现相距不到5 m的3丛, 直到海拔1030 m后才再次出现景宁玉兰植株, 因此, 在720 m处只设置了1个样地, 牛头山上共设置6个样地(表1)。
2.3 样地调查
调查统计样地内景宁玉兰株(丛)数及其冠幅, 观测每丛所有萌条的基径和高度等信息, 记录伴生植物种类, 同时记录样地的经纬度、海拔、坡向、坡度等生态因子及干扰情况。在每个样地内选取2—3丛代表性成年植株, 在其冠层上部标记3根主枝, 用游标卡尺和直尺分别测量各级枝的直径和枝长。按Strahler法确定分枝序列[15], 冠层最外层的第一小枝为第一级, 两个以上第一级相遇即为第二级, 两个以上第二级相遇则为第三级, 依此类推。光照强度采用浙江托普仪器公司的农业环境检测仪(型号DJL-18)。
2.4 数据处理与分析

把松阳县牛头山的景宁玉兰居群分为3个海拔梯度, 即海拔700—1000 m、1000—1200 m和> 1200 m。通过单因素方差分析(ANOVA)分析, 采取LSD差异显著性检验, 比较了不同植被类型和不同海拔高度景宁玉兰萌条的形态特征。为便于统计和分析, 将基径最大的茎干视为母株, 其他茎干视为萌条。所有数据均在Excel 2003和SPSS 17.0软件上进行。
3 结果与分析
3.1 景宁玉兰萌条形态特征
景宁玉兰萌条形态特征及分枝率在不同生境之间存在一定差异(表2)。黄山松林下的景宁玉兰密度最高, 每公顷平均达732丛, 显著多于落叶阔叶林和杉木林(<0.05), 而且落叶阔叶林下景宁玉兰萌条基径显著小于其他林分(< 0.05)。在分枝特征方面, 黄山松和落叶阔叶林中景宁玉兰一级枝直径显著小于其他3种生境(< 0.05), 黄山松林下景宁玉兰一级枝长度即年生长量显著低于其他4种生境(< 0.05)。灌丛中的总分枝率显著高于杉木林和黄山松林(< 0.05), 灌丛和林缘生境上的景宁玉兰逐步分枝率皆处于较高水平, 也许光照条件不受限制, 枝系生长以拓展更大的生存空间。
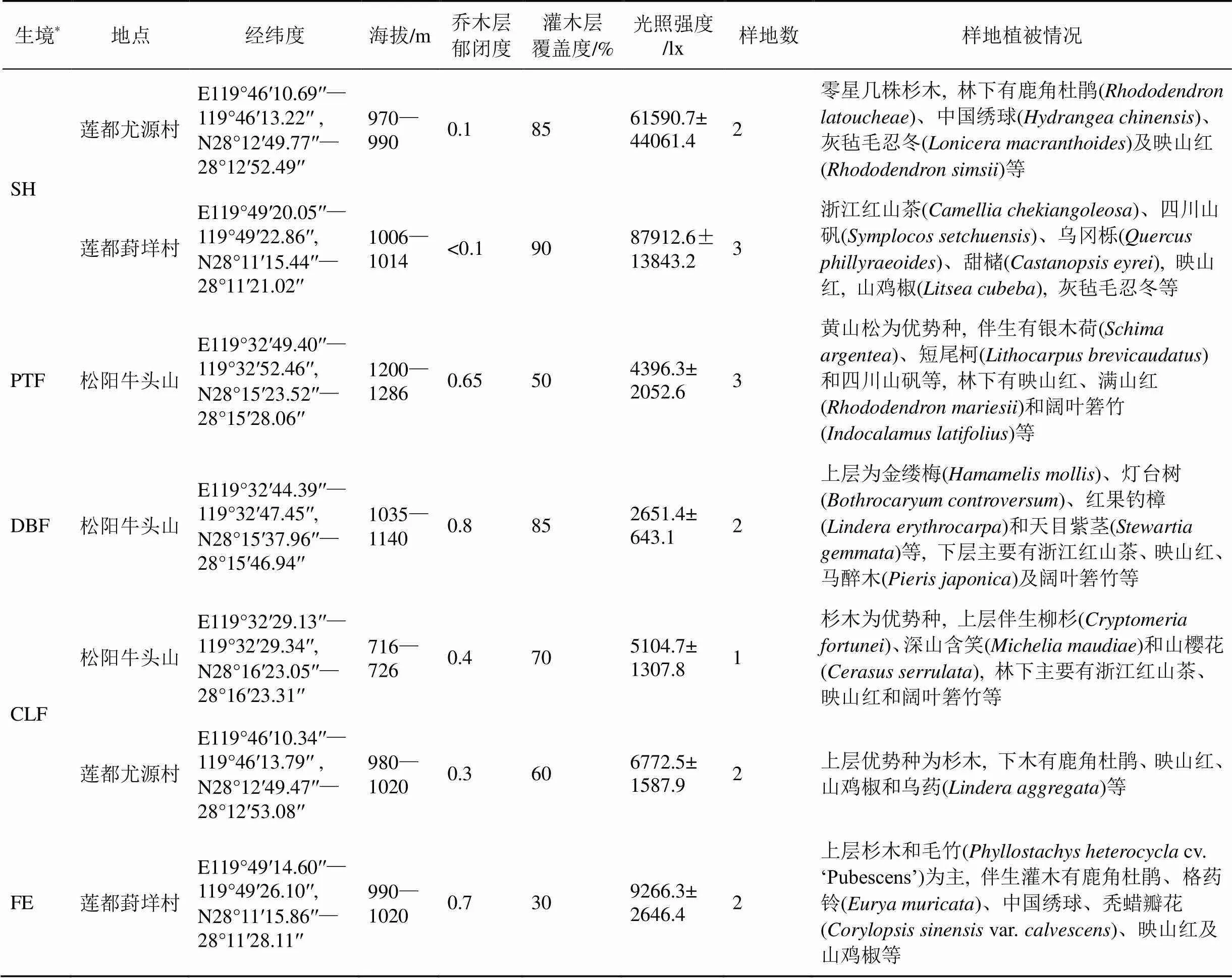
表1 景宁玉兰群落样地概况(均值±标准差)
注: * 此列中SH、PTF、DBF、 CLF 和FE分别代表灌丛、黄山松林、落叶阔叶林、杉木林和林缘生境。

表2 不同生境下景宁玉兰形态特征(均值±标准差)
注: 同列中不同小写字母者表示差异显著水平(< 0.05)。
3.2 萌条形态性状的相关性
通过萌条基径与高度之间相关分析表明, 萌条基径与高度具有显著正相关关系(< 0.01),每丛最大萌条基径与其高度之间具显著正相关关系(< 0.01)(图1 A-B); 最大萌条基径及其高度与每丛萌条数量没有相关关系(图1 C、D)。说明萌条大小及其高度密切相关, 但每丛最大萌条并不影响植株萌生的萌条数量。
3.3 萌条形态性状的变异性
植被类型和海拔高度对景宁玉兰萌条的基径、高度及一级枝长度均存在显著影响(< 0.05), 海拔对一级枝粗度和长度具有显著影响(表3), 说明这些性状与植株所处的生境条件密切相关。虽然不同生境之间的分枝率存在一定的差异(表2), 但总体上来说, 植被类型和海拔高度对每丛萌条数及分枝率的影响不显著。
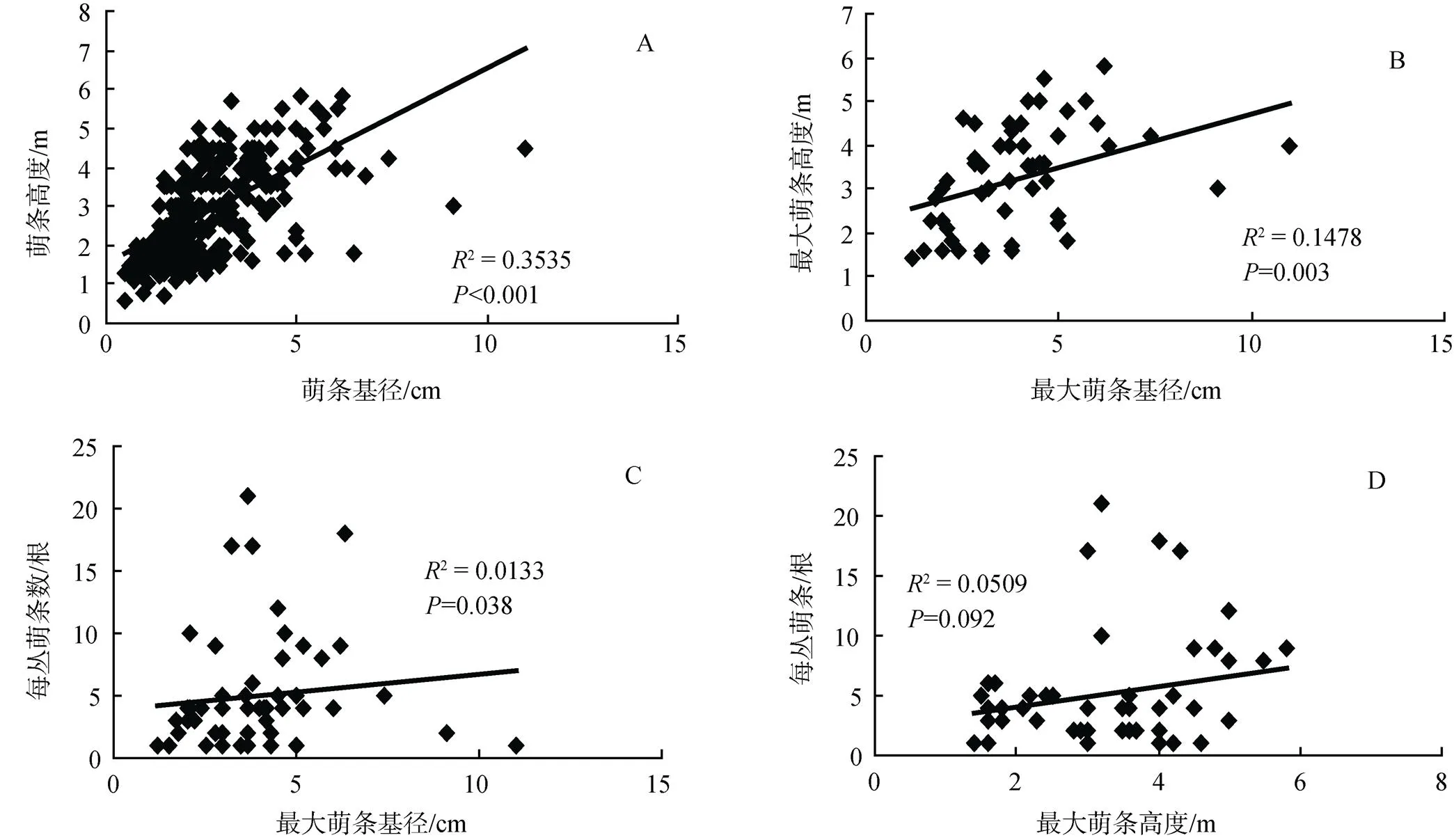
图1 萌条基径与高度(A)之间、最大萌条基径与其高度(B)以及最大萌条基径(C)和高度(D)与每丛萌条数之间相关性

表3 植被类型和海拔对景宁玉兰萌条形态特征的影响
注:*表示显著性差异(<0.05),**表示极显著性差异(<0.01)。
4 讨论
植物的萌生能力受内在生物学特性及微生境、资源水平和干扰状况等外在因素的综合影响[16]。景宁玉兰为萌条能力较强的密集型生长类型[10], 以根颈萌条类型为主[2]。萌条能力与母株的大小不存在相关关系, 并且萌条数量与母株高度也没有相关关系(图1), 这与萌条能力很强的亚热带乔木树种米心水青冈()不同, 米心水青冈的萌条能力与母株的大小以及萌条数量与母株高度皆呈正相关关系[17]。景宁玉兰植株萌条数量在不同生境之间也没有差异, 如在人类活动较为频繁的尤源村灌丛和没有干扰的牛头山黄山松林中, 景宁玉兰每丛的萌条数量没有差异, 但不同生境之间的萌条大小及高度存在差异(表2), 说明萌条能力很可能是因为植物的生物学特性造成的, 而萌条大小及高度差异很可能是由于萌条间以及个体间光照及土壤养分资源的竞争结果[18], 有待深入研究。
萌条形态特征能较好地反映植物对资源环境的适应状况[15]。景宁玉兰在不同生境上, 因为所处的微生境和面临的限制因素各不相同, 分枝构型在不同生境上的表现有较大差异。对林下植物来说, 光照是影响植物萌蘖和增粗的主要因素, 海拔是影响高生长的主要因素[19], 如黄山松和阔叶林下生境, 由于上层乔木树种遮蔽, 具有林下光照较少、种内种间竞争较激烈的特征, 景宁玉兰通过低分枝率(表2)以增加有效光合作用面积。在林缘和灌从中, 则由于具有遮蔽程度小但种间竞争较强, 景宁玉兰主要通过延伸一级枝长度和较高的分枝率, 以拓展上层生存空间, 获得更多的光照进行生长和繁殖[15]。
植物体是作为一个整体对外界环境变化做出反应, 一个性状的改变可能会引起其它性状相应的变化, 本研究只是分析比较了不同生境上景宁玉兰萌条的形态性状特征, 今后应重点关注微生境变化对景宁玉兰生态生理及生殖发育方面的影响, 以进一步了解景宁玉兰的生态适应性机制。
致谢:松阳县林业局景宁玉兰小区护林员雷国兴同志参加部分野外工作, 在此表示感谢!
[1] 裘宝林, 陈征海. 浙江木兰属新种[J]. 植物分类学报. 1989, 27 (1): 79–80.
[2] 斯金平, 徐端妙, 钟炳武, 等. 景宁木兰生物生态特性的初步研究[J]. 浙江林业科技, 2001, 21(4): 11–12, 24.
[3] 汪松, 解炎. 中国物种红色名录[M].北京: 科学出版社, 2004.
[4] 中华人民共和国国务院. 国家重点保护植物名录[J], 植物杂志, 1999, 5: 4–11.
[5] 刘兴剑, 汤诗杰, 孙起梦, 等. 华东植物区木兰科植物的地理分布[J]. 林业调查规划, 2008, 33(5): 40–43.
[6] 余泽智, 陈翔翔, 卢璐, 等. 景宁玉兰种群分布与群落结构研究[J]. 浙江林业科技, 2015, 35(3): 47–52.
[7] 周秀兰, 季必浩, 金民忠, 等. 景宁木兰原生地植物多样性及其保护现状评价[J]. 福建农业科技, 2015, 12: 66–67.
[8] 刘饶, 徐端妙, 潘智慧, 等. 景宁木兰濒危原因初探[J]. 河北农业科学, 2009, 13(6): 31–32.
[9] 卢璐, 余泽智, 刘雪燕, 等. 景宁木兰花粉萌发与贮藏特性研究[J]. 植物研究, 2014, 34(2): 182–187.
[10] 余泽智, 卢璐, 刘雪燕, 等. 景宁木兰穴盘扦插繁殖技术研究[J]. 种子, 2013, 32(4): 118–119.
[11] 张松, 唐亚, 王静, 等. 凹叶木兰萌枝更新及其在物种保存中的意义[J]. 西北植物学报, 2010, 30(4): 669–775.
[12] AUBINI. MESSIER C. KNEESHAW D. Population structure and growth acclimation of mountain maple along a successional gradient in the southern boreal forest[J]. Acoscience, 2005, 12 (4): 540–548.
[13] 蒋燕锋, 刘饶, 斯金平. 景宁木兰与木兰属其它植物之间亲缘关系与遗传基础研究[J]. 浙江林业科技, 2010, 30(2): 22–25.
[14] 张建新.浙江丽水木本植物区系的研究[J]. 武汉植物学研究, 2005, 23(5): 455–460.
[15] 林勇明, 洪滔, 吴承祯. 桂花植冠的枝系构型分析[J]. 热带亚热带植物学报, 2007, 15(4): 301–306.
[16] 朱万泽, 王金锡, 罗成荣, 等. 森林萌生更新研究进展[J]. 林业科学, 2007, 43(9): 74–82.
[17] 赵睿, 周学峰, 徐娜娜, 等. 米心水青冈种群萌条更新与高度生长[J]. 生态学报, 2009,29(7): 3665–3669.
[18] 肖宜安, 何平, 胡文海, 等. 濒危植物长柄双花木自然种群生殖构件的时空动态[J]. 应用生态学报, 2005, 16(7): 1200–1204.
[19] 肖宜安, 何平, 邓洪平, 等. 井冈山长柄双花木种群形态分化的数量分析[J]. 武汉植物学研究, 2002, 20(5): 365–370.
Analysis on morphological characters of sprouts of the endangered plant
DU Youxin1, WU Weijian2, JI Zhili3, WANG Junfeng1, JIANG Lirong3, LIU Yuejun1,*
1. Lishui Academy of Forestry, Lishui 323000, China 2. Songyang Forestry Bureau, Songyang 333400, China 3. Liandu Ecology Forest Farm, Lishui 323000, China
To understand the properties of sprouts ofspecies, 5 habitat types on the same aspect were selected, including shrubberies (SH),forest (PTF), deciduous broad-leaved forest (DBF),forest (CLF) and forest edges (FE). The basal diameter, height, and branching patterns of sproutsofwere investigated in the experimental plots at 5 different habitats. The results are as follows. (1) Plant density ofwas the highest inforest among 5 habitats. There were no differences in sprout number per cluster across 5 habitats. There was no relationship between height of the largest sprout and sprout number per cluster across 5 habitats. (2) Except for sprouts number, the morphological parameters (basal diameter, length, overall bifurcation ratio (OBR), stepwise bifurcation ratio (SBR1-2and SBR2-3), and length of first-order branch) varied with the different habitats. The basal diameter of sprouts ofgrowing in deciduous broad-leaved forests was the smallest among 5 habitats. Both basal diameter and length of the first branches ofgrowing inforest were the smallest among 5 habitats. The overall and stepwise bifurcation ratios in shrubberies and the stepwise bifurcation ratios (SBR2-3) at forest edges were the highest among the 5 habitats. (3) In all sites, 82.5% of all individuals were composed of more than 2 sprouts, indicating that sprouting played an important role in regeneration of thepopulation in Lishui. The research indicated that sprouting characters ofwere mainly controlled by the inherent biological properties. On the other hand, the morphological characters were markedly influenced by the environment conditions.
; sprouting morphology; bifurcation ratio
10.14108/j.cnki.1008-8873.2018.05.020
Q948.1
A
1008-8873(2018)05-152-06
2017-06-01;
2017-07-13
浙江省林业厅生物多样性保护项目(33112410150002); 丽水市重点研发项目(20150404)
杜有新(1965—), 男, 江西宜春人, 博士, 研究员, 主要从事植被生态学及植物保育研究, E-mail:yxdu765@sohu.com
通信作者:刘跃钧, 男, 硕士, 教授级高工, 主要从事中药材研究, E-mail:714117012@qq.com
杜有新, 吴伟建, 季志利, 等. 濒危植物景宁玉兰的萌条形态特征分析[J]. 生态科学, 2018, 37(5): 152-156.
DU Youxin, WU Weijian, JI Zhili, et al. Analysis on morphological characters of sprouts of the endangered plant[J]. Ecological Science, 2018, 37(5): 152-156.
猜你喜欢
北京大学学报(自然科学版)(2022年1期)2022-02-21 05:14:40
绿色科技(2021年13期)2021-07-29 09:13:28
四川林业科技(2019年5期)2019-07-07 07:14:30
安徽农学通报(2019年1期)2019-02-14 02:25:08
湖北林业科技(2019年6期)2019-01-06 02:14:29
黄山学院学报(2017年5期)2017-11-03 02:41:28
湖北林业科技(2017年2期)2017-05-25 18:44:32
山西教育·幼教(2017年11期)2017-04-13 23:40:30
浙江农业科学(2016年11期)2016-05-04 04:16:44
浙江林业(2015年10期)2015-12-11 16:37:31
