
外源NO对Cd胁迫下娃娃菜种子萌发及幼苗生长的保护效应
2018-11-07 06:47余沛东陈银萍闫志强柯昀琪杨波丁浚刚
生态科学 2018年5期
余沛东, 陈银萍, 闫志强, 柯昀琪, 杨波, 丁浚刚
外源NO对Cd胁迫下娃娃菜种子萌发及幼苗生长的保护效应
余沛东, 陈银萍*, 闫志强, 柯昀琪, 杨波, 丁浚刚
兰州交通大学环境与市政工程学院, 甘肃 兰州 730070
为探讨外源一氧化氮(NO)对镉(Cd)胁迫下娃娃菜()的保护效应, 采用0、50、100、200、500和1000 µmol·L–1的外源NO供体硝普钠(SNP)对100和200 mg·L–1CdCl2胁迫下的娃娃菜种子进行处理, 分析其对种子萌发、幼苗生长及生理指标的影响。结果表明: Cd胁迫抑制娃娃菜种子萌发和幼苗生长, 幼苗叶片叶绿素(Chl)和类胡萝卜素(Car)含量显著降低, 丙二醛(MDA)含量和相对电导率(REC)显著上升, 幼苗叶片膜脂过氧化加速。不同浓度的SNP处理, 不仅能提高Cd胁迫下娃娃菜种子的发芽率、发芽势、发芽指数和活力指数, 促进幼苗生长, 而且能抑制MDA含量上升, 降低叶片REC, 增加Chl和脯氨酸(Pro)含量, 增强抗氧化酶活性。外源NO可缓解Cd胁迫对娃娃菜种子萌发及幼苗生长的抑制作用, 缓解Cd胁迫引起的膜脂过氧化, 提高抗Cd能力, 其中以200 µmol·L–1SNP对Cd胁迫的缓解效果较好。
一氧化氮; Cd胁迫; 娃娃菜; 种子萌发; 幼苗生长
1 前言
近年来, 工农业生产使得重金属向环境中的排放增加, 给社会可持续发展带来了严重的影响[1], 比如, 土壤中过量的镉(Cd)除了对农作物生长发育造成严重的影响外, 还会通过食物链危害人类健康[2–3]。因此, 开展如何降低农作物Cd毒害方面的研究具有十分重要的意义。
我国东南沿海地区夏秋季(6—9月)气候炎热, 蔬菜生长不良, 供需矛盾突出, 而利用高原夏季凉爽, 日照充足, 昼夜温差大等气候特点发展起来的高原夏菜, 可以解决这一矛盾。娃娃菜()就是近年来国内学者研究较多的高原夏菜之一, 其具有养胃生津、除烦解渴、利尿通便、清热解毒等功能, 是补充营养、净化血液、疏通肠胃、预防疾病、促进新陈代谢的佳蔬[4]。调节植物生长的信号分子一氧化氮(NO)[5–6]参与了植物对重金属胁迫的响应, 如Cd[7–10]、Mn[11]和Cu[12]胁迫等。NO可以促进种子萌发, 通过增强种子抗氧化能力, 阻止丙二醛(MDA)含量的增加, 清除多余的活性氧(ROS), 从而减轻种子膜脂过氧化损伤。目前, 针对娃娃菜的研究主要集中在无公害和优质栽培技术[13]、病虫害的发生与综合防治[14]、品种的引进与选育[15]及氮素营养的调控机理[16]等方面, 而关于其对重金属特别是Cd的耐受能力和NO对重金属抑制的缓解能力尚未见研究。为此, 本文以娃娃菜为材料, 模拟土壤中Cd胁迫状况, 研究了NO在Cd胁迫下对娃娃菜种子萌发及幼苗生长的影响, 系统地分析不同浓度的NO供体硝普钠(sodium nitroprusside, SNP)对Cd胁迫下娃娃菜种子萌发数量、萌发速度、幼苗生长及生理特性的影响, 旨在为探索缓解Cd污染土壤的最佳SNP浓度, 为Cd污染植物的生态防御研究提供一定的科学依据。
2 材料与方法
2.1 材料和试剂
以娃娃菜品种“韩国2号”(甘肃省农科院提供)为供试材料。NO供体硝普钠(亚硝基铁氰化钠, sodium nitroprusside, SNP, 购自Sigma公司)现用现配, 通常, 0.15 mmol·L–1(mM)的SNP大约可以产生小于0.2 µmol·L–1(µM)的NO[17]。
2.2 实验设计
2.2.1 种子处理
培养时选取籽粒饱满、大小均匀且无病虫伤害的种子洗净种外包衣, 用0.1%的氯化汞消毒10 min, 然后再用蒸馏水冲洗 3—5次, 于水中吸胀12 h后, 吸干表面水分播于铺有两层滤纸的培养皿内进行常规发芽实验。
2.2.2 实验处理
以 Hoagland 营养液为基础培养液, 分别添加Cd2+(CdCl2)至最终浓度为0、100、200 mg·L–1, SNP 至最终浓度为 0、50、100、200、500、1000 µmol·L–1, 共 18 个处理, 每个处理设 6 个重复, 每个重复 50 粒种子。在 20 ℃黑暗中培养 24 h 后, 置于温度为 27 ℃, 光照时间 12 h·d–1的培养箱内培养, 每天定时更换处理液, 其中的3个重复每 24 h 观察发芽数, 以胚芽长超过 1 mm 为发芽标准, 发芽第 4 d 测定发芽势, 第 6 d 测定发芽率、胚芽长、胚根长, 胚根、胚芽干重和鲜重, 计算发芽指数和活力指数; 剩余的3个重复7 d 后将胚根芽长势一致的种子移植到口径为 15 cm 且装有珍珠岩的花盆中, 继续置于培养箱内培养, 每天用相应的处理液处理, 待幼苗三片真叶完全展平时测定叶片相对电导率(REC)、丙二醛(MDA)、脯氨酸(Pro)、叶绿素a(Chla)、叶绿素b(Chlb)、类胡萝卜素(Car)含量及超氧化物歧化酶(SOD)、过氧化氢酶(CAT)和过氧化物酶(POD)等抗氧化酶活性。
2.3 测定方法
发芽指数=∑(/), 其中为t天的发芽数,为发芽日数。
活力指数=发芽指数×胚芽长。
按照紫外分光光度法测定Chl和Car含量[18], 电导率仪法测定REC[18], 硫代巴比妥酸法测定MDA含量[19], 磺基水杨酸-酸性茚三酮比色法测定Pro含量[18], 氮蓝四唑(NBT)法测定SOD活性, 紫外分光光度法测定CAT活性, 愈创木酚法测定POD活性[20]。
2.4 数据分析
所有数据均取 3 次重复平均值, 采用SPSS 19.0统计分析软件进行数据分析及差异显著性检验。
3 结果与分析
3.1 外源NO对Cd胁迫下娃娃菜种子萌发的影响
100 mg·L–1和200 mg·L–1Cd 胁迫均抑制了娃娃菜种子的萌发, 使发芽率、发芽势、发芽指数和活力指数显著下降(<0.05), 100 mg·L–1Cd胁迫下, 分别较空白对照(0 μM SNP+ 0 mg·L–1Cd, CK)下降6.80%、2.06%、19.74%和24.13%, 200 mg·L–1Cd胁迫下, 分别下降10.68%、5.15%、26.29%和38.61%。不同浓度 SNP 处理均能不同程度缓解 Cd 胁迫对种子萌发的抑制作用, 其中, 200 μM SNP 处理使两种浓度Cd胁迫下种子的发芽指数和活力指数较 Cd 单独处理均显著升高(<0.05), 较100 mg·L–1Cd单独处理分别升高21.94%和16.60%, 较200 mg·L–1Cd单独处理分别升高25.85%和37.50%。但随SNP 浓度的升高, 缓解效果就越不明显(表1)。双因素方差分析表明, Cd、SNP以及Cd与SNP交互作用对发芽率和发芽势没有显著影响, 而对发芽指数和活力指数有显著的影响(表2)。
3.2 外源NO对Cd胁迫下娃娃菜幼苗生长的影响
两种浓度Cd胁迫均使胚芽长、胚芽鲜重和胚芽干重降低, 其中200 mg·L–1Cd单独处理更显著(<0.05), 较CK分别下降17.24%、4.67%和3.11%。不同浓度SNP处理均能促进Cd胁迫下娃娃菜幼苗的生长, 其中, 200 μM SNP处理使幼苗胚根长、胚芽长、胚根鲜重、胚芽鲜重、胚根干重和胚芽干重显著升高(<0.05), 较100 mg·L–1Cd单独处理分别升高22.80%、17.88%、7.02%、14.68%、7.17%和15.74%, 较200 mg·L–1Cd单独处理分别升高42.78%、22.837%、7.19%、10.01%、9.32%和22.48%。但随SNP浓度的升高, 促进效果就越不明显(表3)。双因素方差分析表明, 胚根鲜重、胚芽鲜重、胚根干重、胚芽干重、胚根长、胚芽长均受到Cd、SNP以及Cd与SNP交互作用的显著影响(表2)。

表1 不同浓度SNP对Cd胁迫下娃娃菜种子萌发的影响
注: 同列中不同小写字母表示差异显著(<0.05)。

表2 Cd、SNP以及Cd与SNP交互作用对娃娃菜种子萌发和幼苗生长影响的双因素方差分析
注: *表示方差分析结果为极显著性差异(<0.001)。下同。
3.3 外源NO对Cd胁迫下娃娃菜幼苗叶片Chl和Car含量的影响
两种浓度Cd胁迫均使娃娃菜幼苗叶片Chla、Chlb、Chl总量和Car含量降低, 其中200 mg·L-1Cd单独处理更显著(<0.05), 较CK分别下降33.03%、77.91%、40.54%和27.66%。不同浓度SNP处理均能促进Chl合成, 其中, 200 μM SNP处理使Chla、Chlb、Chl总量和Car含量增加, 较100 mg·L–1Cd单独处理分别升高6.19%、1.16%、5.35%和1.58%, 较200 mg·L–1Cd单独处理分别升高15.77%、158.42%、24.62%和3.39%(<0.05)。但当SNP浓度继续增加时, Chl含量不再增加, 且略有下降的趋势(图1)。双因素方差分析表明, Cd、Cd与SNP交互作用对Chlb含量有显著影响, Cd、SNP以及Cd与SNP交互作用对Chla、Chl总量和Car含量有显著影响(表4)。

注: 图中不同小写字母表示各处理间差异显著(P<0.05)。下同
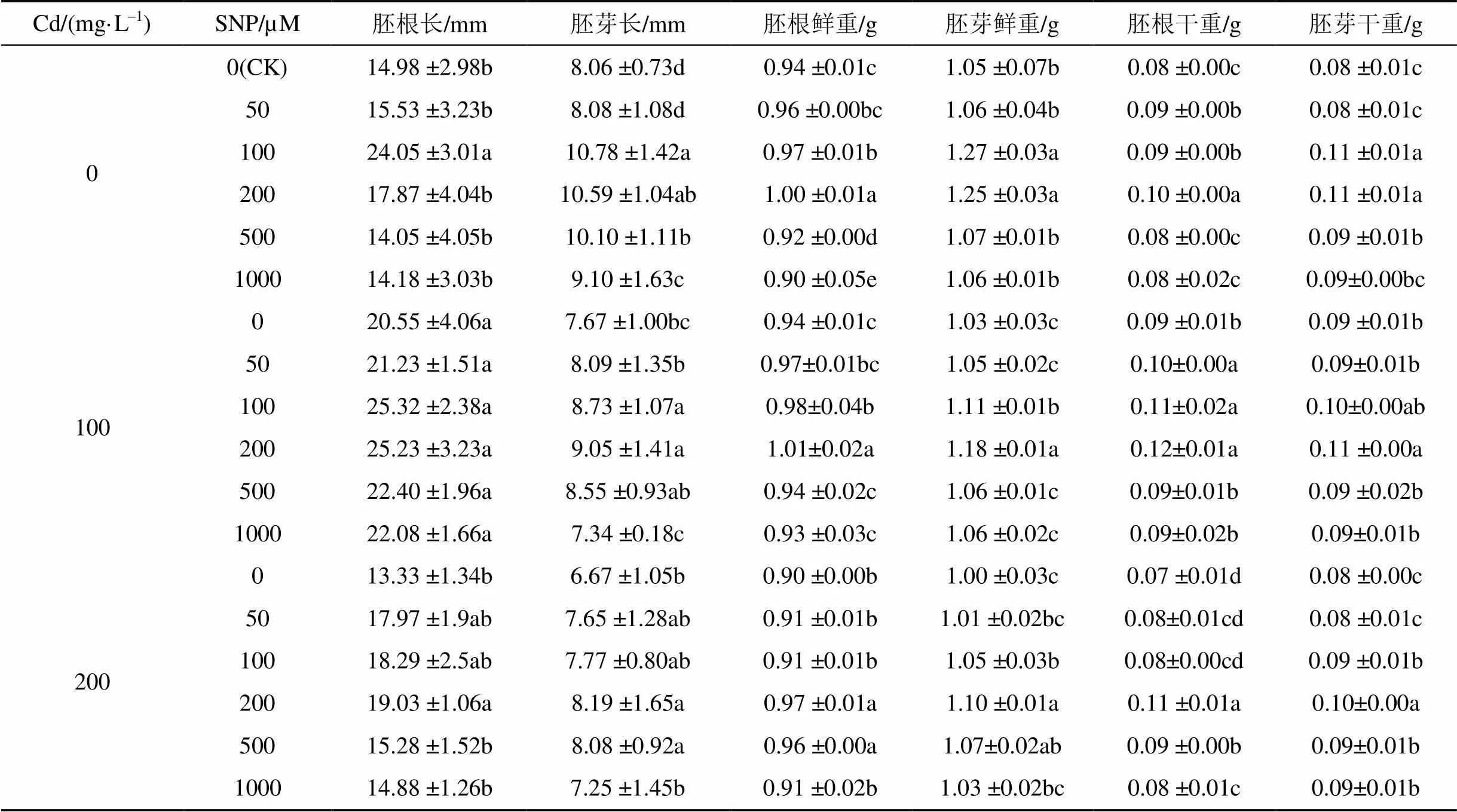
表3 不同浓度SNP对Cd胁迫下娃娃菜幼苗生长的影响
3.4 外源NO对Cd胁迫下娃娃菜幼苗叶片REC和MDA含量的影响
Cd 胁迫下, 娃娃菜幼苗叶片REC和MDA含量均显著增加(<0.05), 100 mg·L–1Cd胁迫下, 分别较CK增加40.55%和171.28%, 200 mg·L–1Cd胁迫下, 分别增加183.46%和211.00%。不同浓度 SNP均能不同程度降低REC 和 MDA 含量, 其中, 200 μM SNP处理降低效果最显著(<0.05), 较100 mg·L–1Cd单独处理分别降低32.39%和19.91%, 较200 mg·L–1Cd单独处理分别降低53.77%和40.15%。但当 SNP 浓度继续增加时, 降低效果略有下降(图2)。双因素方差分析表明, Cd 与 SNP 交互作用对 MDA 有显著影响, Cd、SNP以及Cd与SNP交互作用对REC有显著影响(表4)。
3.5 外源NO对Cd胁迫下娃娃菜幼苗叶片SOD、CAT和POD活性及Pro含量的影响
Cd胁迫下, 娃娃菜幼苗叶片抗氧化酶活性显著增强, Pro含量显著增加(<0.05), 100 mg·L–1Cd胁迫下, SOD、CAT和POD活性及Pro含量分别较CK增加177.92%、5.98%、100.02%和68.58%, 200 mg·L–1Cd胁迫下, 分别增加161.04%、19.06%、75.02%和74.53%。不同浓度SNP处理能进一步增强抗氧化酶活性, 增加Pro含量, 其中, 200 μM SNP处理增强效果最显著(<0.05), 较100 mg·L–1Cd单独处理分别增加20.56%、11.07%、100.02%和35.13%, 较200 mg·L–1Cd单独处理分别增加54.73%、0.91%、150.00%和31.72%。但随SNP浓度增加, 抗氧化酶活性增强和Pro含量增加趋势越不显著(图3)。双因素方差分析表明, Pro、SOD、CAT 、POD受到 Cd、SNP以及Cd与SNP交互作用的显著影响(表4)。
4 讨论
Cd胁迫下, 进入植物细胞的Cd2+与酶活性中心或蛋白巯基结合, 取代蛋白反应中心的Ca2+、Mg2+、Fe2+等必需金属, 向细胞中释放自由离子, 也能刺激脂氧合酶和还原型烟酰胺腺嘌呤二核苷酸磷酸(nicotinamide adenine dinucleotide phosphate, NADPH)氧化酶活性, 使细胞内O2–·和H2O2等活性氧生成增加, 从而引起氧化胁迫和膜脂过氧化, 进而影响植物的呼吸作用以及氮素、碳水化合物和核酸等重要物质代谢, 导致细胞分裂与植物生长受到抑制[8, 21]。本研究中, Cd胁迫抑制娃娃菜种子萌发和幼苗生长, 幼苗叶片Chl含量显著降低, 直接影响光合作用和光合产物的积累, 造成植物株高和生物量的减少, MDA含量和REC显著上升, 幼苗叶片膜脂过氧化加速, 进一步证明了Cd胁迫对植物种子萌发和幼苗生长造成的影响。
作为一种新的植物生长调节信号分子, NO广泛参与植物生长、发育及各种生物和非生物胁迫[22–24]。已有研究表明, NO也参与了植物对各种重金属胁迫的响应, SNP能促进低浓度 Pb 胁迫下杂花苜蓿()[25]、Cd胁迫下水稻()[7]和Cu胁迫下小麦()[12]种子的萌发。本研究中, 不同浓度SNP能提高Cd胁迫下娃娃菜种子的发芽率、发芽势、发芽指数和活力指数, 这与前人研究结果相似, 双因素方差分析表明, Cd、SNP以及Cd与SNP交互作用对发芽指数和活力指数有显著的影响, 进一步证明了外源NO可以缓解Cd胁迫对娃娃菜种子萌发的抑制作用。
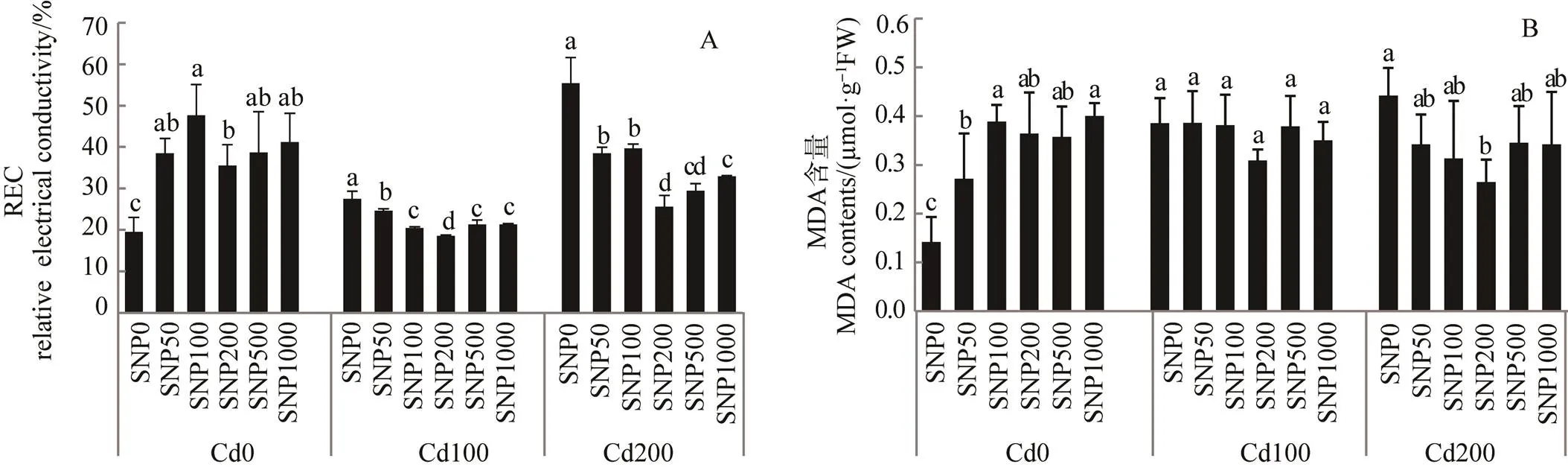
图2 NO对Cd胁迫下娃娃菜叶片REC(A)和MDA含量(B)的影响

图3 NO对Cd胁迫下娃娃菜叶片SOD(A)、CAT(B)、POD(C)活性和Pro(D)含量的影响
叶绿素是植物进行光合作用所必需的重要物质, 决定着植物的光合效率[26]。一定浓度SNP处理能促进Cd胁迫下娃娃菜幼苗叶片中Chl合成, 促进幼苗生长, 双因素方差分析表明, Cd、SNP以及Cd与SNP交互作用对Chla、Chl总量、Car含量、胚根鲜重、胚芽鲜重、胚根干重、胚芽干重、胚根长、胚芽长均有显著影响。可能是SNP增加了Ca、Fe、Cu、Zn等矿质元素的吸收, 使叶绿素结构破坏过程减缓, 减弱了Chl的降解反应, 或者是活化了叶绿素生物合成过程中的某些酶类, 增加了叶绿素的合成, 从而维持叶绿素含量[27], 提高了叶片细胞的渗透调节能力和耐毒能力, 促进了幼苗生长。
生物膜与植物抗性有关, 正常条件下, 植物体内ROS的产生与清除处于动态平衡状态, 植物遭受逆境胁迫时, 这种动态平衡被破坏, 引发膜脂过氧化反应[8]。外源NO能减少逆境胁迫下植物体内ROS的积累从而缓解氧化胁迫对植物的伤害[28]。本研究中, Cd胁迫诱导了娃娃菜幼苗细胞膜脂过氧化的产生, 而SNP能在一定程度上缓解这种损伤, 抑制MDA含量的上升, 降低REC, 可能是NO同ROS或脂质过氧化自由基发生了反应, 使氧化胁迫中断[29]。另外, 本研究中, 适当浓度的SNP处理能进一步增强SOD、CAT、POD活性, 促进Pro积累, 表明NO也能通过激活抗氧化系统, 增强活性氧的清除效率, 从而较好地维持细胞膜的稳定性, 减轻Cd胁迫诱导的氧化损伤, 这与NO缓解Cd胁迫对水稻[7,9]、蚕豆()[8]及亚麻()[30]幼苗伤害的研究结果一致。
但是, 这种缓解作用对NO浓度有一定的依赖性。本研究表明, 当SNP浓度低于 200mM 时, SNP处理能促进娃娃菜种子萌发和幼苗生长, 使发芽势、发芽率、发芽指数和活力指数增大, 幼苗的胚根长、胚芽长及胚根、胚芽生物量增加, 幼苗叶片Chl含量增加, MDA含量和REC降低, Pro含量增加, 抗氧化酶活性增强, NO有效地缓解了Cd胁迫下娃娃菜叶片中ROS的伤害。但当SNP浓度超过 200mM 时, 娃娃菜种子萌发和幼苗生长随SNP浓度的增加而受到抑制, Chl含量降低, MDA含量和REC增大, Pro含量降低, 抗氧化酶活性减弱, 使娃娃菜幼苗受害更加明显。这与冉烈等[31]、陈银萍等[32]和杨志娟等[33]的研究结果一致, 表明娃娃菜对Cd胁迫的抗性并不是随SNP使用浓度的增加而增强的。

表4 Cd、SNP以及Cd与SNP交互作用对娃娃菜叶片生理特性影响的双因素方差分析
低浓度NO可作为抗氧化剂清除ROS, 也能诱导抗氧化酶基因的表达, 但高浓度的NO会与ROS发生作用, 生成大量的过氧亚硝酸阴离子(OONO-), OONO-质子化后形成具有强氧化性的过氧亚硝酸(HOONO), 破坏生物大分子的结构与功能, 具有更强的生物毒性。因此, NO在植物体内以适宜的浓度存在才能起到最佳的生物学效应, 浓度过高或过低, 都无法调控下游的信号转导, 表明NO在生物体内的作用具有浓度依赖性。
5 结论
外源NO可提高Cd胁迫下娃娃菜种子萌发率、幼苗叶片中Chl含量、Pro含量和抗氧化酶活性, 缓解MDA的积累, 降低REC, 保护细胞膜免受或减少损伤, 缓解对娃娃菜种子萌发及幼苗生长的抑制作用, 提高植物抗Cd能力。使用SNP浓度过低和过高均不理想, 200 µM SNP对娃娃菜Cd胁迫的缓解效果最佳。
[1] XU X H, ZHAO Y C, ZHAO X T, et al. Sources of heavy metal pollution in agricultural soils of a rapidly industrializing area in the Yangtze Delta of China[J]. Ecotoxicology and Environmental Safety, 2014, 108: 161–167.
[2] 崔秀敏, 吴小宾, 李晓云, 等.铜、镉毒害对番茄生长和膜功能蛋白酶活性的影响及外源NO的缓解效应[J]. 植物营养与肥料学报, 2011, 02: 349–357.
[3] AGRAWAL S B, MISHRA S. Effects of supplemental ultraviolet-B and cadmium on growth, antioxidants and yield ofL. [J]. Ecotoxicology and Environmental Safety, 2009, 72(2): 610–618.
[4] 张凯, 张玉鑫, 陈年来, 等. 甘肃省高原夏菜种植气候区划[J]. 西北农林科技大学学报: 自然科学版, 2012, 40(5): 179–185.
[5] SAXENA I, SHEKHAWAT G S. Nitric oxide (NO) in alleviation of heavy metal induced phytotoxicity and its role in protein nitration[J]. Nitric Oxide, 2013, 32: 13–20.
[6] CHEN Y P, ZHANG J, YANG Z J, et al. Relationship between nitric oxide accumulation, anti-oxidative system and freezing tolerance in the leaves ofduring cold adaptation[J]. Journal of Animal and Plant Sciences, 2012, 22(4): 1133–1141.
[7] HE J Y, REN Y F, CHEN X L, et al. Protective roles of nitric oxide on seed germination and seedling growth of rice(L.) under cadmium stress[J]. Ecotoxicology and Environmental Safety, 2014, 108: 114–119.
[8] 李源, 魏小红, 朱蕾, 等. 外源 NO 和 H2O2对Cd胁迫下蚕豆种子萌发及幼苗生长的保护效应[J]. 安徽农业科学, 2009, 37(11): 4942–4944, 4959.
[9] PRERNA S, KAVITA S. Evidences for reduced metal-uptake and membrane injury upon application of nitric oxide donor in cadmium stressed rice seedlings[J]. Plant Physiology and Biochemistry, 2014, 83: 180–184.
[10] JOZEF K, BOŘIVOJ K, PETR B, et al. Nitric oxide donor modulates cadmium-induced physiological and metabolic changes in the green alga[J]. Algal Research, 2015, 8: 45–52.
[11] JOZEF K, PETR B, JOSEF H, et al. Manganese-induced oxidative stress in two ontogenetic stages of chamomile and amelioration by nitric oxide[J]. Plant Science, 2014, 215-216: 1–10.
[12] HU K D, HU L Y, LI Y H, et al. Protective roles of nitric oxide on germination and antioxidant metabolism in wheat seeds under copper stress[J]. Plant Growth Regulation, 2007, 53: 173–183.
[13] 蒋红英. 娃娃菜农残消降组合方法及对品质的影响[J].食品与发酵工业, 2011, 37(4): 92–96.
[14] 湛长菊. 娃娃菜的特征特性及高产栽培技术[J]. 现代农业科学, 2008, (19): 63–65.
[15] 张晓梅, 严湘萍. 高寒地区娃娃菜品种引种比较试验初报[J]. 长江蔬菜, 2009, (1): 46–47.
[16] 杨兵丽, 张国斌, 周箬涵, 等. 不同形态氮素配施对娃娃菜光合和Chl荧光特性的影响[J].浙江农业学报, 2015, 27(5): 761–768.
[17] 孙群, 胡景江.植物生理学研究技术[M]. 陕西: 西北农林科技大学出版社, 2006.
[18] 李合生, 孙群, 赵世杰, 等. 植物生理生化实验原理和技术[M]. 北京: 高等教育出版社, 2000, 164–261.
[19] 樊怀福, 郭世荣, 焦彦生, 等. 外源一氧化氮对NaCl胁迫下黄瓜幼苗生长、活性氧代谢和光合特性的影响[J]. 生态学报, 2007, 27(2): 546–553.
[20] 李忠光, 李江鸿, 杜朝昆, 等. 在单一提取系统中同时测定五种植物抗氧化酶[J]. 云南师范大学学报: 自然科学版, 2002, 22(6): 44–48.
[21] 陈世军, 张明生. 外源NO供体SNP对Cd胁迫下辣椒种子萌发的影响[J]. 种子, 2009, 28(2): 21–25.
[22] 陈银萍, 陶玲, 杨莉, 等.一氧化氮对水分胁迫下种子玉米种子萌发及幼苗生理特性的影响[J].广西植物, 2010, 30(5): 672–677.
[23] ZHENG C F, JIANG D, LIU F L, et a1. Exogenous nitric oxide improves seed germination in wheat against mitochondrial oxidative damage induced by high salinity[J]. Environmental and Experimental Botany, 2009, 67: 222–227.
[24] XU Y F, SUN X L, JIN J W, et al. Protective effect of nitric oxide on light-induced oxidative damage in leaves of tall fescue[J]. Journal Plant Physiology, 2010, 167: 512–518.
[25] 向华, 南丽丽, 李春晓. 一氧化氮对铅胁迫下杂花苜蓿种子萌发及幼苗生理特性的影响[J]. 草原与草坪, 2014, 34(2): 77–80.
[26] 刘燕, 蒋光霞. 硒对铅胁迫下油菜酶活性及叶绿素含量的影响[J].安徽农业科学, 2008, 36(18): 7554–7555, 7849.
[27] 张义凯, 崔秀敏, 杨守祥, 等. 外源NO对镉胁迫下番茄活性氧代谢及光合特性的影响[J]. 应用生态学报, 2010, 21(6): 1432–1438.
[28] 赵宝泉, 万宇, 杨世湖, 等. 外源 NO 供体硝普钠(SNP)对重金属Cd胁迫下水稻幼苗膜脂过氧化及抗氧化酶的影响[J]. 江苏农业学报, 2010, 26(3): 468–475.
[29] MATA C G, LAMATTINA L. Nitric oxide induces stomatal closure and enhances the adaptive plant responses against drought stress[J]. Plant Physiology, 2001, 126: 1196–1204.
[30] 徐严, 魏小红, 李兵兵. 外源NO对重金属Cd胁迫下亚麻幼苗叶片抗氧化能力的影响[J]. 甘肃农业大学学报, 2012, 47(5): 45–49.
[31] 冉烈, 李会合, 田秀英. 外源NO对镉胁迫下东南景天生长和镉累积的影响[J]. 中国农业通报, 2012, 28(19): 60-64.
[32] 陈银萍, 蘧苗苗, 苏向楠, 等. 外源一氧化氮对镉胁迫下紫花苜蓿幼苗活性氧代谢和镉积累的影响[J].农业环境科学学报, 2015, 34(12): 2261–2271.
[33] 杨志娟, 陈银萍, 苏向楠, 等. 外源NO对铅胁迫下三叶鬼针草幼苗活性氧代谢的影响[J]. 广西植物, 2015, 35(5): 648–655.
Protective effects of nitric oxide on seed germination and seedling growth ofunder cadmium stress
YU Peidong, CHEN Yinping*, YAN Zhiqiang, KE Yunqi, YANG Bo, DING Jungang
School of Environmental and Municipal Engineering, Lanzhou Jiaotong University, Lanzhou 730070, China
To investigate the effects of nitric oxide (NO) on mitigating Cd stress on, the seeds (under 100 and 200 mg·L–1CdCl2stress) were treated with 0, 50, 100, 200, 500, and 1000 µmol·L–1sodium nitroprusside (SNP) as an exogenous NO donor, as well as the seed germination, seedling growth, and physiological indicators of leaves were measured. The results showed that Cd stress inhibited seed germination and seedling growth, decreased chlorophyll (Chl) andCarotenoid (Car) contents, increased malondialdehyde (MDA) and relative electrical conductivity (REC), and accelerated membrane lipid peroxidation. SNP at different concentrations performed well to promote seed germination and seedling growth, and to decrease MDA content and cell membrane permeability, increase Chl, Car and proline (Pro) contents, and activate the antioxidant enzyme system in seedlings under Cd stress. It was shown that exogenous NO not only mitigated the inhibiting effects on seed germination and seedling growth and mitigated the lipid peroxidation induced by Cd stress, but also enhanced the plants resistance to Cd stress. The most optimal SNP concentration was suggested at 200 µmol·L–1.
nitric oxide (NO); Cd stress;; seed germination; seedling growth
10.14108/j.cnki.1008-8873.2018.05.002
X173;Q142
A
1008-8873(2018)05-007-09
2018-01-05;
2018-03-04
国家自然科学基金项目(31560161, 31260089, 31640012)
余沛东(1993—), 男, 甘肃天水人, 硕士, 主要从事环境生态学, E-mail: yupeidong7878@foxmail.com
通信作者:陈银萍(1974—), 女, 甘肃榆中人, 教授, 主要从事环境生态学, E-mail: yinpch@mail.lzjtu.cn
余沛东, 陈银萍, 闫志强, 等. 外源 NO对Cd胁迫下娃娃菜种子萌发及幼苗生长的保护效应[J]. 生态科学, 2018, 37(5): 7-15.
YU Peidong, CHEN Yinping, YAN Zhiqiang, et al. Protective effects of nitric oxide on seed germination and seedling growth ofunder cadmium stress[J]. Ecological Science, 2018, 37(5): 7-15.
猜你喜欢
舰船科学技术(2022年11期)2022-07-15
青年文学家(2021年4期)2021-03-18
农民致富之友(2020年19期)2020-07-18
西藏农业科技(2019年3期)2019-11-04
家庭百事通·健康一点通(2019年11期)2019-09-26
现代园艺(2018年3期)2018-02-10
上海农业学报(2017年3期)2017-04-10
小学生导刊(低年级)(2016年8期)2016-09-24
学苑创造·A版(2016年6期)2016-06-20
饮食科学(2014年8期)2014-08-22
