
大型生物气田形成与富集条件研究
——以东地中海黎凡特盆地为例
2018-11-02 01:35贾怀存康洪全王春修邱春光阳怀忠
石油实验地质 2018年5期
贾怀存,康洪全,王春修,邱春光,阳怀忠,白 博
(1.中海油研究总院有限责任公司,北京 100028; 2.中国海洋石油国际有限公司,北京 100027)
随着油气勘探技术的不断发展,世界石油工业在近几年的勘探中呈现出二大特征:一是逐步向深水迈进;二是生物气的发现逐步成为各大石油公司油气勘探的热点。如我国的柴达木盆地、埃及的尼罗河三角洲盆地、俄罗斯的西西伯利亚盆地以及孟加拉湾等多个含油气盆地均发现了大量的生物气资源。近几年,尤其是在2009—2011年,在东地中海黎凡特(Levantine)盆地相继发现的塔玛和利维坦生物气田均是当年全世界十大油气发现之一,2个气田的探明+控制可采储量(2P可采储量)分别为2 775×108m3和4 814×108m3。2011年,美国Noble公司在黎凡特盆地的塞浦路斯海域又发现了阿芙罗狄生物气藏,初步估算可采储量约为1 982×108m3。尽管世界各地的很多盆地均发现了生物气田,国内外很多专家学者也对生物气形成机制、成因类型判别、成藏特征和分布规律等方面开展了大量的工作[1-7],但对形成如此大规模的纯生物成因的生物气田的主控因素与预测等方面的研究还比较薄弱,尤其是在大型生物气田成藏条件与主控因素方面[8-15]。因此,借助对黎凡特盆地大型生物气田形成条件的探讨,可以丰富对大型生物气田成藏主控因素的认识。
1 区域地质概况
黎凡特盆地位于东地中海,从南向北依次跨越埃及、以色列、黎巴嫩和叙利亚领海。盆地西部部分位于塞浦路斯海域,盆地东部和北部分别以死海走滑断裂带和塔尔图斯(Tartus)断裂带为界,向西延伸至爱拉托辛海岭(Eratosthenes Sea Mount)和尼罗河三角洲(Nile Delta)。全盆地面积6.8×104km2,其中5%位于陆上,85%位于深水区。盆地从二叠纪以来开始接受沉积,沉积地层厚度最大可达15 000 m。
根据盆地构造变形机制与沉积地层关系,将黎凡特盆地分为5个构造单元,分别是东南冲断带、南部坳陷带、中央隆起带、北部坳陷带和北部冲断带(图1)。在构造演化上,黎凡特盆地经历了较为完整的威尔逊旋回。在晚二叠世,伴随着冈瓦纳大陆裂解和特提斯洋的形成,盆地经历了克拉通边缘裂谷阶段,在东南部发育了一系列的正断层。晚侏罗—早白垩世,盆地进入裂后的被动大陆边缘发育期,盆地构造活动稳定,地层沉积厚度较大。晚白垩世开始,伴随着特提斯洋的关闭和非洲—阿拉伯板块与欧亚板块的会聚,盆地进入挤压收缩的变形阶段,著名的“叙利亚弧”挤压构造带就是在这一时期形成。同时,盆内早期的正断层发生构造反转,北部冲断带和东南冲断带逐步形成。上新世—现今,受控于东部死海断裂带左旋走滑的影响,形成了一系列的压扭构造(图2)。
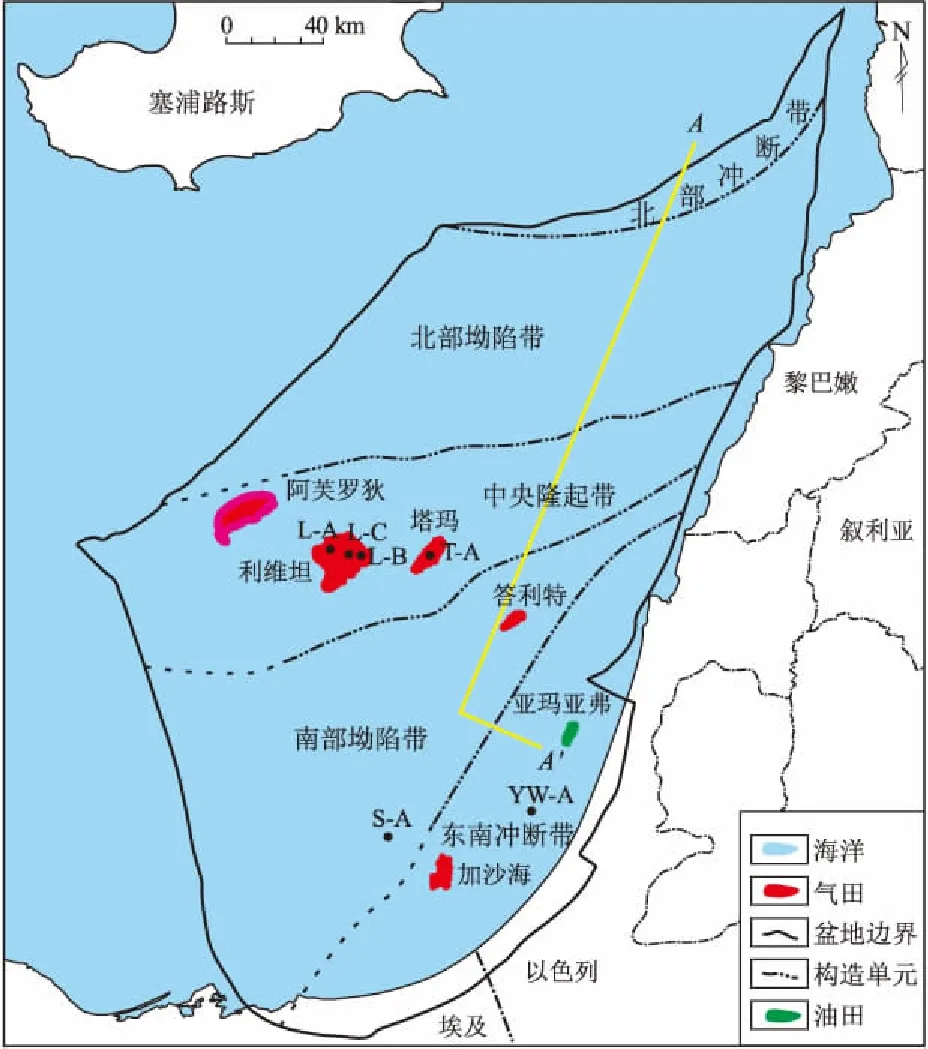
图1 黎凡特盆地构造纲要Fig.1 Tectonic units of Levantine Basin
2 生物气地化特征及成因类型
在黎凡特盆地中央隆起带发现的利维坦、塔玛和阿芙罗狄气田以及上新统气田,均为典型的纯生物气气田。生物气成因主要为乙酸发酵作用;生物气田的烃源岩为渐新统—中新统的海相泥岩,主力储层为渐新统—下中新统的塔玛组砂岩。天然气干燥系数一般在96%~99%之间,甲烷碳同位素(δ13C1)普遍在-61‰~-63‰。
甲烷碳同位素和重烃气组分含量(C2+3)是鉴别生物气的重要地化指标。微生物作用形成的生物气δ13C1一般小于-55‰,而热成因气δ13C1一般为-20‰~-55‰[16]。根据微生物发生生化作用的环境不同,又可以分为乙酸发酵和CO2还原作用。利用δ13C1的大小可以大致判别生物气形成途径,当δ13C1<-60‰时,生物气主要为乙酸发酵成因;当δ13C1>-60‰时,生物气主要为海相环境中的CO2还原作用形成的[17-21]。
根据天然气成因类型判别指标[16,22],在天然气成因类型判别图版上(图3),黎凡特盆地上新统气田[4]和渐新统—中新统的塔玛气田生物气均落在纯生物气区,表明黎凡特盆地气田属于典型的纯生物气田;甲烷碳同位素普遍小于-60‰,说明生物气形成途径主要为乙酸发酵作用。
3 大型生物气田形成与富集条件
3.1 优质烃源岩提供充足气源
黎凡特盆地中央隆起带发现的利维坦、塔玛等多个大生物气田证实,盆地生物气烃源岩具有类型好、品质优、厚度大和分布范围广的特征,为生成大量的生物气提供了充足的气源条件。
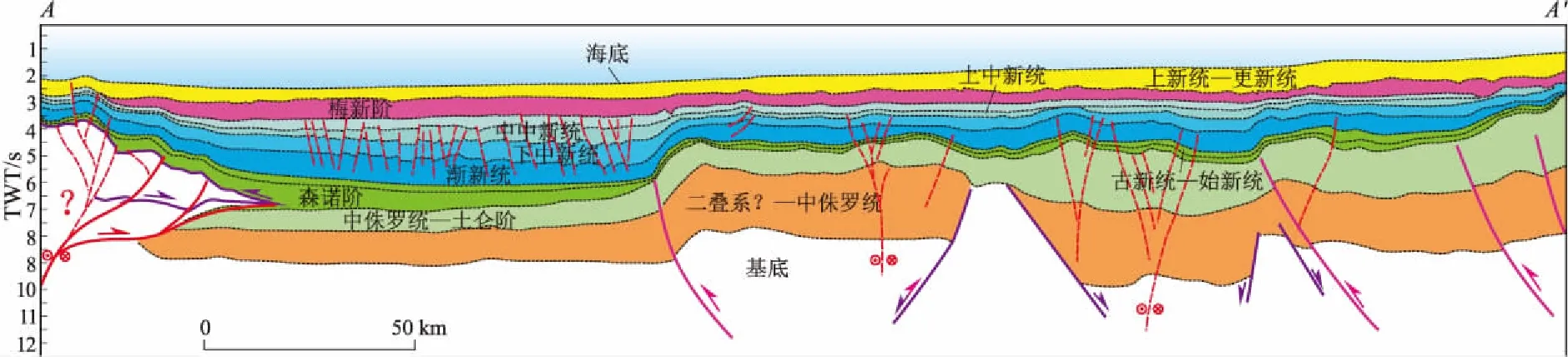
图2 黎凡特盆地区域地质剖面剖面位置见图1的AA’。Fig.2 Regional cross section of Levantine Basin

图3 黎凡特盆地生物气成因类型判别Fig.3 Generation type of biogas in Levantine Basin
黎凡特盆地渐新统和中新统海相烃源岩地化指标优,有机质类型好,利于生成大量生物气。前人研究表明,可溶性有机质是产甲烷菌优质母质,而富含藻类、微生物、类脂化合物的Ⅰ型和Ⅱ型有机质中富含可溶性有机质,更利于生成生物气[22-26]。黎凡特盆地利维坦气田的L-A井揭示,渐新统—中新统的海相泥岩有机质类型为Ⅱ2型,氢指数普遍在300 mg/g左右,生烃潜力S2普遍在2 mg/g左右。可见,黎凡特盆地渐新统—中新统烃源岩地化指标优,有机质类型利于生成大量生物气。
优质的生物气烃源岩厚度大,分布范围广,为黎凡特盆地形成大型生物气田提供了充足气源。黎凡特盆地渐新统地层厚度普遍为800~1400m,且横向连续分布,尤其是位于中央隆起带两侧的南部坳陷带和北部坳陷带,渐新统和中新统海相泥岩厚度最大可达1 600 m左右。巨厚富含可溶性有机质的海相泥岩,为中央隆起带形成利维坦和塔玛等大型生物气田提供了充足气源。
黎凡特盆地在渐新统—中新统烃源岩沉积时期具有较低的地温环境,具备生成大量生物气的地质条件。前人研究表明,产甲烷菌的存活一般要求较低的地温环境和厌氧环境,实验表明产甲烷菌存活的温度环境为0~80 ℃,主生气带的温度为30~65 ℃[27-29]。根据利维坦气田L-A井揭示的地温梯度为1.6 ℃/hm,是典型的冷盆。另外,盆地东南部的YW-A井古地温模拟(图4)表明,在渐新世—中新世期间古地温在30 ℃左右,进入中新世后受“叙利亚弧”挤压构造带快速形成的影响,地层温度也快速上升,至中新世末地层温度达到60 ℃左右,而现今地层温度为80 ℃。古地温的变化表明,渐新统—中新统的海相泥岩从沉积后一直到中新世末,均位于产甲烷菌最佳活跃温度范围,烃源岩处于主力生生物气期,而现今依然在生生物气。
黎凡特盆地具有快速的地层堆积速率,利于大量生物甲烷的生成与保存。快速的地层堆积一方面可以保持厌氧环境,同时也为大量的产甲烷菌存在提供了有效的生存空间[28-30],为形成高丰度的生物气烃源岩提供了基础条件。根据L-A井地层沉积速率分析,渐新世以来地层沉积速率普遍在200 m/Ma左右;晚中新世沉积速率最快,可达1 000 m/Ma。快速的地层堆积速率,一方面可以使沉积的有机质分布相对分散,有效烃源岩的生气范围较为广泛;另一方面可使可溶性有机质快速进入厌氧环境,免于有机质被氧化,利于产甲烷菌分解有机质生成大量生物气。另外,快速地层堆积可以及时对早期生成的生物气进行有效保存,提供了较为优越的保存条件。
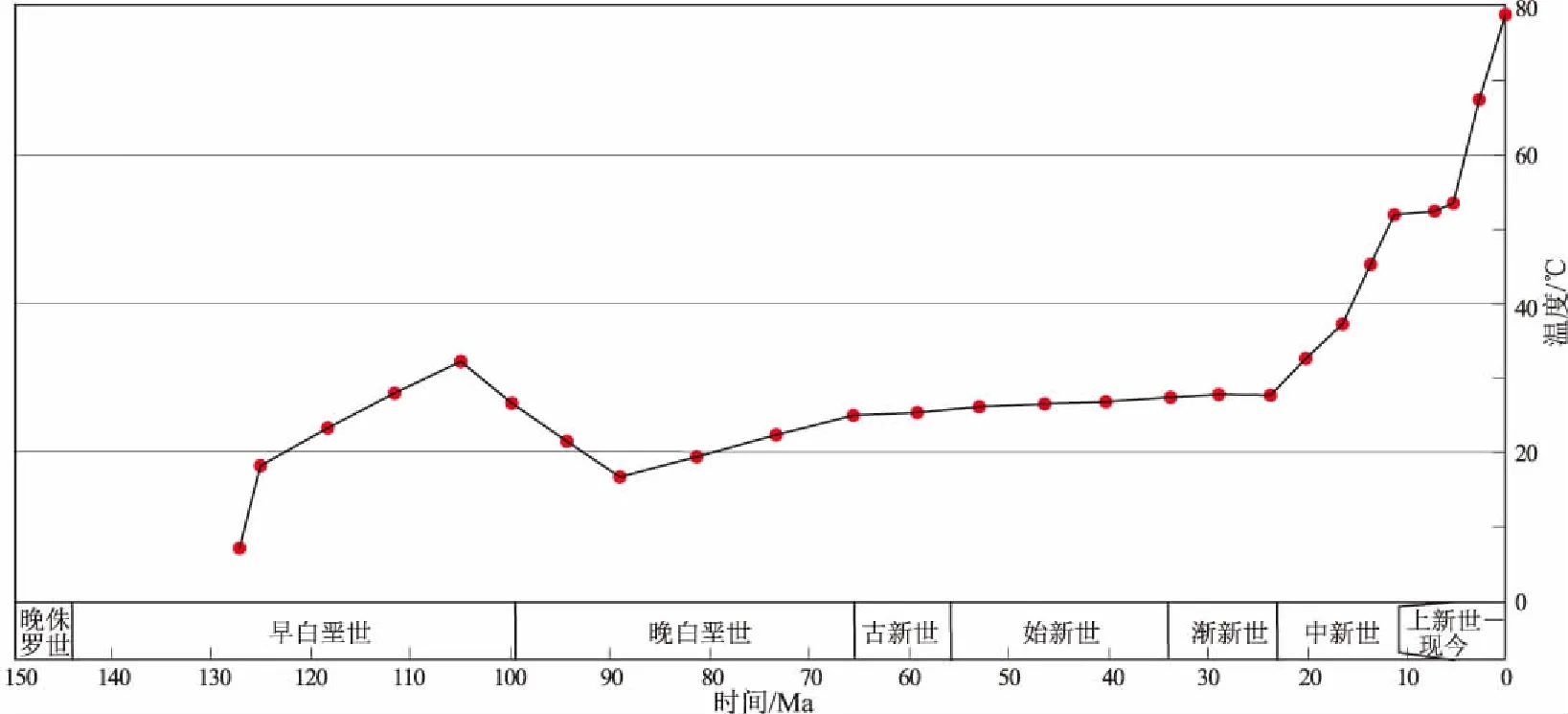
图4 黎凡特盆地YW-A井古地温模拟Fig.4 Geothermal model of well YW-A,Levantine Basin
3.2 广泛分布的海底扇砂岩储层提供高效汇聚通道
众所周知,生物气往往在初次运移就可聚集成藏,因此,及时高效的油气汇聚通道是生成大型生物气田的重要条件。黎凡特盆地在中新统发育广泛分布的海底扇砂岩(图5),尤其是在中央隆起带,下中新统海底扇砂岩储层平面上连片分布,垂向上砂岩厚度大,储层物性好,是渐新统和中新统海相泥岩生成的生物气的高效输导体。中央隆起带的利维坦气田、塔玛气田和答利特气田的多口钻井证实,下中新统塔玛组广泛发育3期海底扇沉积,以厚层砂岩为主;储层顶部最小埋深3 610 m,含砂率高,一般在55.7%~73.5%,单层砂岩最大厚度33.5~140 m,平均孔隙度为25%,平均渗透率为1 μm2。
在中央隆起带,多期海底扇和浊积水道垂向上叠置,平面上连片分布。下中新统广泛分布的厚层优质砂岩储层,为渐新统和中新统海相泥岩生成的生物气成藏提供了高效的汇聚通道。目前该套储层是利维坦、塔玛、答利特气田以及塞浦路斯海域的阿芙罗狄气田等多个大型生物气田的主力储集层,可采储量占全盆地可采储量的85%以上。因此,广泛分布的海底扇优质砂岩储层,为形成大型生物气田提供了高效的汇聚通道。
3.3 发育的继承性隆起提供有利聚集场所
继承性隆起构造带之上往往发育大型的、构造变形相对较弱的同沉积背斜圈闭,利于及时捕集早期生成的生物气,是大型生物气田的主要富集区带。而远离继承性隆起带,如斜坡带和冲断带大型背斜圈闭往往不发育,以发育水道等规模相对较小的岩性圈闭为主,形成的气藏规模一般较小。
黎凡特盆地中央隆起带是继承性发育的大型隆起带,发育大型同沉积背斜圈闭,而生物气往往是在早成岩阶段生成,因此,同沉积圈闭是生物气聚集的有利场所。通过构造恢复表明,中央隆起带在渐新世具有低幅度隆起特征;中新世以来,伴随着叙利亚弧挤压和死海断裂带的不断影响,中央隆起带受到持续的挤压应力作用,隆起幅度逐步增强,并形成了一系列的挤压背斜圈闭。如位于中央隆起带的利维坦、塔玛、答利特等背斜圈闭,伴随着晚白垩世时期的构造反转,在渐新世逐步开始形成,并在中新世的叙利亚弧Ⅱ期构造活动中最终定型。中央隆起带上的大型背斜圈闭一般背斜形态较为完整,圈闭规模大,圈闭面积往往在188~594km2

图5 黎凡特盆地渐新世(a)和早中新世(b)沉积相平面分布Fig.5 Sedimentary facies of Oligocene (a) and Lower Miocene (b),Levantine Basin
左右。这些发育在继承性隆起之上的大型同沉积背斜圈闭,为早期形成的生物气成藏提供了有利的聚集场所。
黎凡特盆地斜坡带和东南冲断带大型同沉积背斜圈闭不发育,主要形成水道砂岩体等岩性圈闭,圈闭规模较小。如位于斜坡带的S-A井,在中新统钻遇多支水道叠加的岩性圈闭,初步预测生物气地质资源量为170×108m3。
3.4 具备及时的保存条件
天然气成藏是一个充注与散失相互平衡的动态过程,上覆优质盖层的及时封盖对生物气成藏至关重要。当生物气上覆盖层厚度大于100 m、渗透率为0.1~1 μm2时,盖层封盖能力较好;当盖层厚度大于50 m、渗透率小于0.1 μm2时,盖层封盖能力则非常好[28,30]。巨厚优质的直接盖层和分布稳定的区域盖层,是形成大型生物气田的关键因素。
黎凡特盆地渐新统—中新统发育巨厚海相泥岩,可以对聚集的生物气提供及时的保存条件,具有优越的储盖配置条件。塔玛气田的T-A井和利维坦气田的L-A井、L-B井和L-C井均揭示,在主力储层渐新统塔玛组砂岩之上发育近500 m厚的海相泥岩,在上覆的中新统梅辛阶发育区域性沉积的巨厚盐岩,厚度可达1 500 m左右。巨厚的海相泥岩和区域性沉积的盐岩与塔玛组海底扇砂岩形成优质的储盖配置条件,可以为大型生物气田的形成提供及时的保存条件。
4 结论
(1)黎凡特盆地已发现的大型生物气田均为典型的纯生物气气田,甲烷碳同位素值普遍小于-60‰,生物气成因主要为乙酸发酵作用。
(2)黎凡特盆地形成大型生物气田主要受控于4项地质条件: ①渐新统—中新统发育巨厚优质烃源岩,为生物气聚集成藏提供了充足的气源;②中新统海底扇优质砂岩储层的连片发育,一方面为生物气成藏提供了优质储集条件,同时广泛分布的海底扇砂岩储层也为生物气运聚提供了高效的汇聚通道;③继承性隆起利于大型优质背斜圈闭的形成,为形成大型生物气田提供有利圈闭条件;④上覆巨厚的泥岩盖层提供了优越的储盖配置条件,可以对形成的生物气藏进行及时保存。
(3)黎凡特盆地继承性隆起带往往发育大型的同沉积背斜圈闭,广泛分布海底扇砂岩储层,是寻找大型生物气田的主要勘探方向。
猜你喜欢
化工管理(2021年7期)2021-05-13
小学阅读指南·低年级版(2021年3期)2021-03-19
矿产勘查(2020年9期)2020-12-25
中国海上油气(2020年5期)2020-10-20
学苑创造·A版(2020年9期)2020-10-13
小学科学(学生版)(2020年7期)2020-07-28
西南石油大学学报(自然科学版)(2018年1期)2018-02-10
海峡科技与产业(2016年3期)2016-05-17
当代化工研究(2016年7期)2016-03-20
油气地质与采收率(2014年6期)2014-12-16
