
不同生境内厚朴种子萌发和幼苗生长研究
2018-10-30 10:28:28杨志玲程小燕李公荣马文明
生态与农村环境学报 2018年10期
谭 美,杨志玲①,杨 旭,程小燕,李公荣,马文明
(1.中国林业科学研究院亚热带林业研究所/ 浙江省林木育种技术研究重点实验室,浙江 杭州 311400;2.浙江省磐安县园塘林场,浙江 磐安 322300)
种子是植物生活史的重要阶段,也是植物在时空上逃避不利环境以确保成功定居和更新的保障。种子萌发和幼苗生长是种群更新过程的重要环节,种子萌发特性和自然分布区气候与生境条件密切相关,是植物对分布区自然生境长期适应的结果[1]。当种子萌发形成幼苗时,个体即从一生中风险最小的阶段跨越到风险最大也最为脆弱的阶段[2],使种子向幼苗、幼树过渡的生长期成为众多珍稀濒危植物的生长瓶颈之一[3]。种子萌发和幼苗生长受生物和非生物因素综合影响,不仅反映植物对环境资源的竞争能力和利用效率,且关系到种群生存和发展以及森林发育、演替和更新[4]。
厚朴(Houpoöaofficinalis)为木兰科(Magnoliaceae)厚朴属(Houpoöa)植物,一般分布在海拔300~1 500 m的山地林间湿润肥沃向阳处[5],是我国珍贵的药、材两用树种,大量研究证明厚朴具有的酚类成分对治疗炎症、癌症和心脑血管类疾病等有重要作用[6]。由于长期不规范管理和过度砍伐,厚朴野生群体和个体急剧减少,已被列为我国二级保护植物[7]。近年来,学者对厚朴野生资源濒危机制和保护策略开展大量研究[8],在野外调查中发现厚朴野生种群天然更新十分困难,且在不同群落类型内分布存在差异[9],但对厚朴种子成熟脱落后种子萌发和幼苗生长的影响因子等鲜有报道。
厚朴可通过无性繁殖和有性繁殖2种方式更新种群,但无性繁殖限于树蔸处产生的根蘖苗,无法在林中自由扩散,对种群更新和重建实质意义不大。有性繁殖才是野外种群更新和种群重建的主要方式,实生苗对维持和扩展种群基因多样性和种群规模有重要意义,但实生苗定居时较高死亡率和较慢生长速度影响种群更新[10]。
种子萌发并形成正常苗,除受到外界自然条件影响外,还与种子自身特性和其他动物干扰有关。假种皮是种子外覆盖的一层特殊结构,因内含多种种子萌发抑制物,对种子萌发有一定限制作用[11]。而有些种子种皮坚硬,产生使活性胚在适宜环境中不能萌发的种皮障碍[12]。另外,动物对种子的取食会减少土壤种子库中种子含量,又会破坏种子内部结构,影响种子正常萌发和幼苗健康生长,使用铁丝网罩能减少外界动物损坏厚朴种子的几率,提升种子萌发率或生长成幼苗的机会。因此,笔者在已开展相关研究[9,13]基础上,设计厚朴种子野外人为散播试验,观察针叶林、针阔混交林和阔叶林3种不同生境内厚朴种子萌发和幼苗生长特征,阐述不同林型所在生境对厚朴种子萌发和幼苗生长影响机制,研究厚朴假种皮和动物取食对野外厚朴种子萌发影响程度,发现厚朴野生种群建立的障碍,为厚朴野生资源保护和科学开发利用提供理论依据。
1 材料与方法
1.1 研究区概况
实验基地位于浙江省遂昌县桂洋林场,地理位置为28°21′01″ N,119°08′31″ E,海拔1 100 m,年均温12.3 ℃,夏季最高温30.2 ℃,冬季最低温-12.5 ℃,年均降水量2 400 mm,平均相对湿度80%,年日照时数为1 515.5 h,属于典型的中亚热带海洋性季风气候区。土壤为红壤,山地坡度一般为30°~35°,峡谷坡度达45°~50°。基本满足厚朴喜凉爽、湿润、光照充足和土壤疏松、肥沃、腐殖质多、排水良好等生长要求,是厚朴传统种植地区和道地产区之一,厚朴在林场各林型内均有不同程度分布。针叶林中主要树种为杉木(Cunninghamialanceolata)、黄山松(Pinustaiwanensis)和日本柳杉(Cryptomeriajaponica)等;针阔混交林主要树种为黄山松、枫香(Liquidambarformosana)和甜槠(Castanopsiseyrei)等;阔叶林中主要树种为木荷(Schimasuperba)和东南石栎(Lithocarpusharlandii)。
1.2 研究方法
1.2.1样地设置
2016年11月厚朴种子雨结束后,在针叶林、针阔混交林和阔叶林3个不同林型样地分别设置12个1 m×1 m小样方进行种子萌发野外实验。实验设计去除假种皮且安装铁丝网罩(A)、去除假种皮不安装铁丝网罩(B)、保留假种皮且安装铁丝网罩(C)、保留假种皮不安装铁丝网罩(D)4个处理,每个处理重复3次,3个林型共有36个小样方。假种皮去除方法为种子从球果上剥落后,用冷水浸泡至假种皮软化,用水搓洗掉假种皮。铁丝网罩长×宽×高为1 m×1 m×0.1 m,网眼大小为0.5 cm×0.5 cm。每个样方均匀散播100粒新鲜饱满的厚朴种子,散播前清除小样方内杂草和枯落物,并挖去3 cm左右深土壤,种子散播后将挖出的土壤、枯落物和腐殖质均匀回填在小样方内。
1.2.2样地调查
温度、湿度(TSTORC-HT601A 智能便携式数据仪,国产品牌)和[光]照度(TES-1336A光照测定仪,台湾泰式)分别于晴天(前期至少7 d无降雨)08:00、10:00、12:00、14:00和16:00在各样地中心地表以上0.5 m高处测定;同时测量样地内枯落腐殖层厚度。取各样地表层(0~20 cm)土壤,测定土壤有机质含量(重铬酸钾氧化外加热法)、水解氮含量(碱解扩散吸收法)、有效磷含量(钼锑抗比色法)、速效钾含量(火焰光度法)和土壤含水率(烘干法)[14]。3种不同生境环境因子及土壤成分见表1。
表1不同生境环境因子及土壤成分
Table1Environmentalfactorsandsoilcompositionofdifferenthabitats
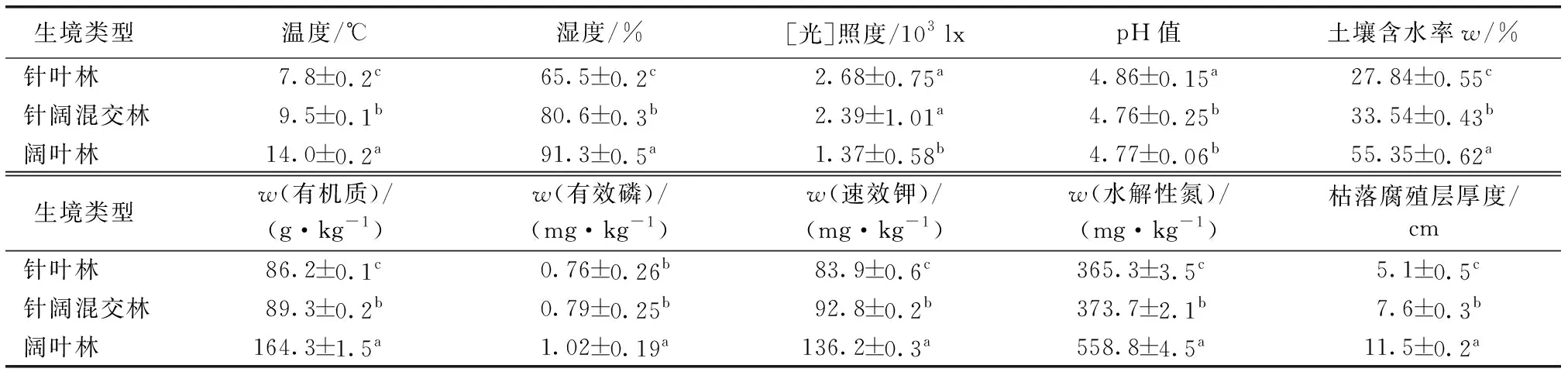
生境类型 温度/℃湿度/%[光]照度/103 lxpH值土壤含水率w/% 针叶林7.8±0.2c65.5±0.2c2.68±0.75a4.86±0.15a27.84±0.55c 针阔混交林9.5±0.1b80.6±0.3b2.39±1.01a4.76±0.25b33.54±0.43b 阔叶林14.0±0.2a91.3±0.5a1.37±0.58b4.77±0.06b55.35±0.62a 生境类型 w(有机质)/(g·kg-1)w(有效磷)/(mg·kg-1)w(速效钾)/(mg·kg-1)w(水解性氮)/(mg·kg-1)枯落腐殖层厚度/cm 针叶林86.2±0.1c0.76±0.26b83.9±0.6c365.3±3.5c5.1±0.5c 针阔混交林89.3±0.2b0.79±0.25b92.8±0.2b373.7±2.1b7.6±0.3b 阔叶林164.3±1.5a1.02±0.19a136.2±0.3a558.8±4.5a11.5±0.2a
同一列数据后英文小写字母不同表示不同林型间某指标差异显著(P<0.05)。
1.3 数据统计与分析
于2017年4月开始调查厚朴种子萌发率、存活率和幼苗生长状况。在幼苗生长末期(10月),将每个小样方内存活幼苗连根挖出,用塑料袋装好带回实验室后用清水洗净,用滤纸吸干表面水分,测量幼苗株高和主根长;采用电子天平称量根、茎和叶片鲜重后,用80 ℃烘箱烘干,称量干生物量。萌发率=样地出苗量/样地播种量×100%,存活率=当月存活数量/萌发数量×100%,根茎比=主根长/茎长(株高),根冠比=根干质量/茎叶干质量。数据分析和作图采用Excel 2013和SPSS 19.0软件。
2 结果与分析
2.1 不同生境对野外种子萌发的影响
2.1.1不同生境环境因子及土壤成分对比
对针叶林、针阔混交林和阔叶林的环境因子和土壤成分进行单因素方差分析和多重比较(表1)发现,3种生境中温度,湿度,枯落腐殖层厚度,土壤有机质、速效钾、水解性氮含量和土壤含水率差异显著(P<0.05),而阔叶林的[光]照度和有效磷含量与针叶林和针阔混交林差异显著(P<0.05),充分说明3种生境存在较大异质性。
2.1.2不同生境各处理厚朴种子萌发率对比
不同生境各处理种子萌发率见表2。2017年4月未见厚朴种子萌发,5月初发现2处样地有种子萌发生芽,5月底厚朴种子萌发基本完成。统计发现,A处理种子萌发率最高,B处理次之,C和D处理萌发率几乎为0(表2),说明去除假种皮后种子萌发率明显提高,保留假种皮的种子几乎不萌发。A处理的种子萌发率高于B处理,C处理种子萌发率高于D处理,即不管是否去除假种皮,安装铁丝网罩都有利于种子萌发。A和B处理在不同生境内的种子萌发率均呈显著差异(P<0.05),表明微生境对于野外种子萌发有一定影响。
表2不同生境各处理厚朴种子萌发率
Table2Houpoöaofficinaisseedgerminationpercentageunderdifferenttreatmentsandhabitats
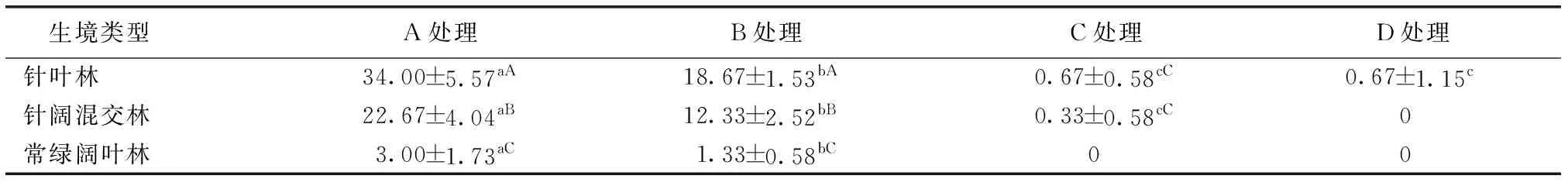
%
A为去除假种皮且安装铁丝网罩处理;B为去除假种皮不安装铁丝网罩处理;C为保留假种皮且安装铁丝网罩处理;D为保留假种皮不安装铁丝网罩处理。同一行数据后英文小写字母不同表示处理间种子萌发率差异显著(P<0.05),同一列数据后英文大写字母不同表示生境间种子萌发率差异显著(P<0.05)。
2.2 不同生境对厚朴幼苗生长的影响
2.2.1不同生境厚朴幼苗存活情况
9月针叶林内厚朴存活率最高,为(29.74±6.25)%;针阔混交林次之,为(15.24±4.23)%;阔叶林内厚朴幼苗全部死亡(图1)。从5月底种子萌发到7月底,即初期(0~8周)幼苗死亡最严重。这是因为初期幼苗根系发育不完全,且7月浙江磐安县降雨量大,土壤含水量过高造成幼苗根部腐烂,致使幼苗大量死亡,故6—7月幼苗死亡率较大。8—9月厚朴幼苗存活逐渐稳定,针叶林中幼苗存活情况最佳,这与针叶林中光照强度远高于其他2种林型有关。9月,针叶林和针阔混交林内A和B处理厚朴平均存活率分别为29.05%和22.68%。
2.2.2不同生境厚朴幼苗生长情况
从6月开始,不同生境厚朴幼苗生长差异明显,针叶林和针阔混交林明显优于阔叶林(P<0.05),每月增高量也大于阔叶林(表3),说明针叶林与针阔混交林更适合厚朴幼苗生长。通常7—8月因高温幼苗基本停滞生长,但因实验地海拔较高,7—8月气候仍适宜厚朴幼苗生长。
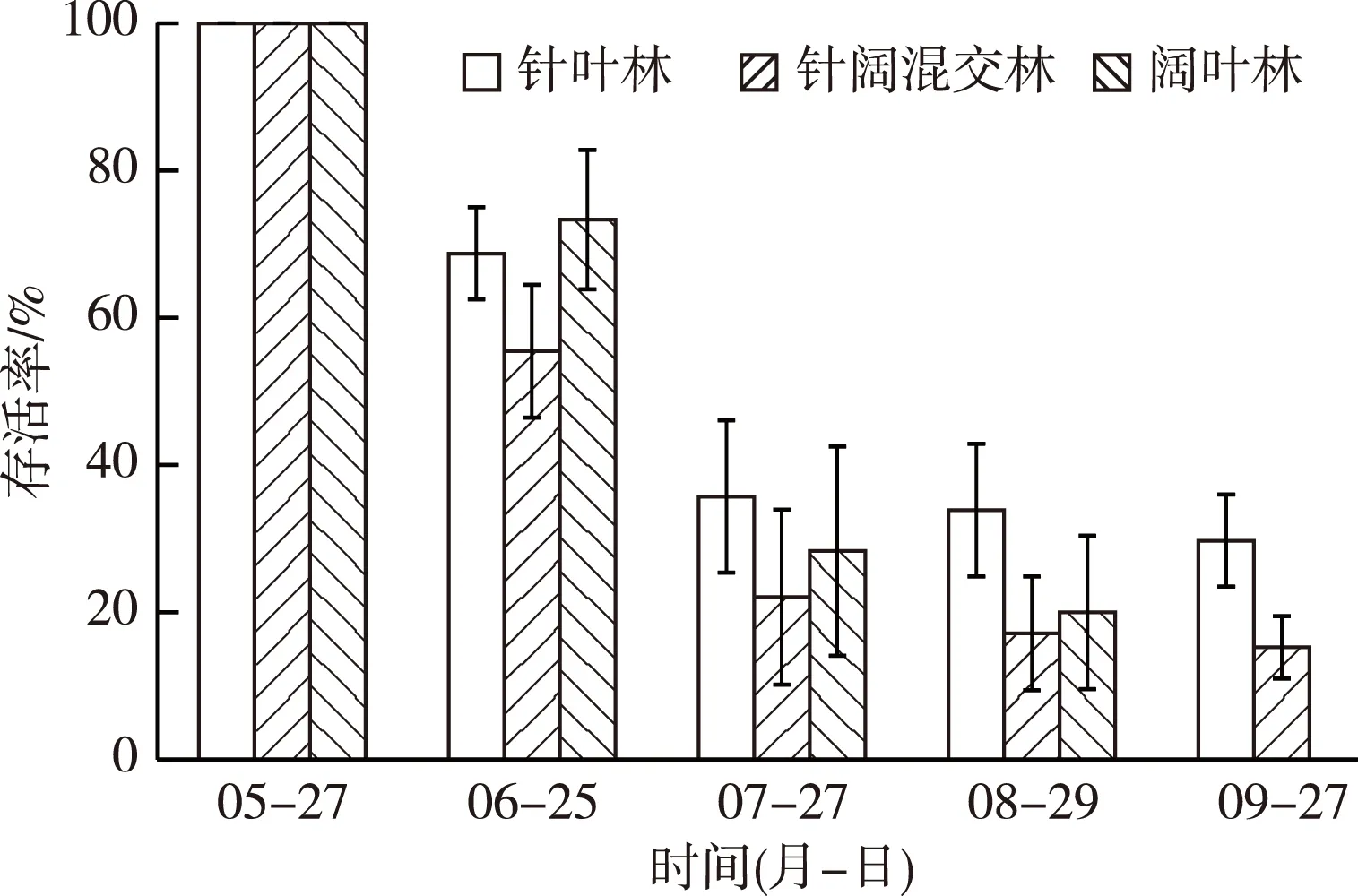
图1 厚朴幼苗在不同生境内的存活情况
表3不同生境厚朴幼苗生长情况
Table3HeightandbasaldiameterofHoupoöaofficinalisseedlingsindifferenthabitats

生境类型 苗高/cm地径/mm6月7月8月6月7月8月 针叶林3.72±0.85aC5.51±0.53aB8.72±1.16aA2.09±0.09aB2.60±0.35aB3.60±0.27aA 针阔混交林3.55±0.45aC4.61±0.40bB7.78±0.81bA2.04±0.05aB2.18±0.08bB3.30±0.24bA 阔叶林2.57±0.59bB3.45±0.58cB4.54±0.59cA1.86±0.14bA2.02±0.09bA2.21±0.11cA
同一行数据后英文大写字母不同表示不同月份间某指标差异显著(P<0.05),同一列数据后英文小写字母不同表示不同林型间某指标差异显著(P<0.05)。
相同处理条件下,针叶林中幼苗主根长显著高于针阔混交林(P<0.05),同一种生境A处理厚朴幼苗主根长和根茎比均显著高于B处理(P<0.05,表4)。
表4不同生境厚朴幼苗主根长和根茎比
Table4LengthofmainrootandRSRofHoupoöaofficinalisseedlingsindifferenthabitats
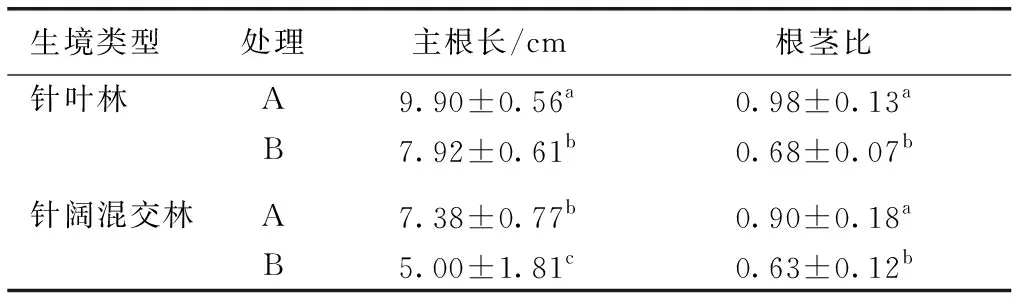
生境类型处理主根长/cm根茎比 针叶林A9.90±0.56a0.98±0.13aB7.92±0.61b0.68±0.07b 针阔混交林A7.38±0.77b0.90±0.18aB5.00±1.81c0.63±0.12b
A为去除假种皮且安装铁丝网罩处理;B为去除假种皮不安装铁丝网罩处理。同一列数据后英文小写字母不同表示不同处理间某指标差异显著(P<0.05)。
由图2可知,不同生境厚朴幼苗不同部位生物量分配大致为叶生物量>根生物量>茎生物量。相同处理条件下,针叶林和针阔混交林幼苗不同部位生物量差异显著(P<0.05),针叶林幼苗根冠比虽略大于针阔混交林,但差异不显著。相同生境条件下,针叶林内A处理厚朴幼苗不同部位生物量小于B处理,针阔混交林内A处理则大于B处理,但处理间差异不显著。

A为去除假种皮且安装铁丝网罩处理;B为去除假种皮不安装铁丝网罩处理。同一组直方柱上方英文小写字母不同表示不同处理间某指标差异显著(P<0.05)。
2.3 环境因子对厚朴种子萌发和幼苗生长的影响
环境因子对植物生活史不同阶段影响也不同。由表5可知,种子萌发与温度、湿度和枯枝落叶层厚度呈显著负相关(P<0.05),与光照强度呈显著正相关(P<0.05),即温度低、湿度大、枯枝落叶层厚且光照不足的环境会增加种子被真菌感染、霉变和虫蛀腐烂的几率,影响野外厚朴种子萌发。
表5环境因子与种子萌发、幼苗生长和生物量的相关系数
Table5Correlationanalysisbetweensproutingofseeds,traitandbiomassofseedlingsandenvironmentalfactors
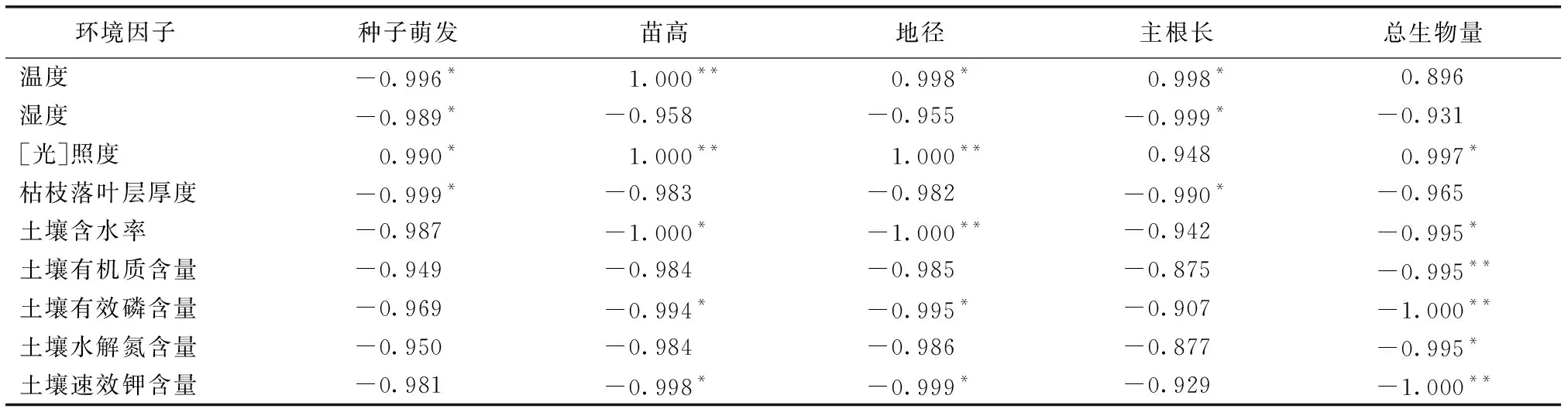
环境因子种子萌发苗高地径主根长总生物量 温度-0.996* 1.000** 0.998* 0.998*0.896 湿度-0.989*-0.958-0.955-0.999*-0.931 [光]照度0.990*1.000**1.000**0.9480.997* 枯枝落叶层厚度-0.999*-0.983-0.982-0.990*-0.965 土壤含水率-0.987-1.000*-1.000**-0.942-0.995* 土壤有机质含量-0.949-0.984-0.985-0.875-0.995** 土壤有效磷含量-0.969-0.994*-0.995*-0.907-1.000** 土壤水解氮含量-0.950-0.984-0.986-0.877-0.995* 土壤速效钾含量-0.981-0.998*-0.999*-0.929-1.000**
*表示P<0.05;**表示P<0.01。
苗高、地径和主根长反映幼苗生长发育状况,对实生苗个体能否进入主林层有重要意义。由表5可知,幼苗苗高和地径与土壤含水率、土壤有效磷和速效钾含量呈显著负相关(P<0.05),与温度、[光]照度呈极显著正相关(P<0.01);主根长与温度呈显著正相关(P<0.05),与湿度和枯枝落叶厚度呈显著负相关(P<0.05),说明光照充足、温度适宜的环境有利于厚朴幼苗生长,而过高的土壤含水率、有效磷和速效钾含量不利于厚朴幼苗生长。
总生物量反映幼苗生长过程中物质积累情况,是幼苗生长状况的表征指标之一。厚朴幼苗总生物量与[光]照度呈显著正相关(P<0.05),与枯枝落叶厚度,土壤含水率以及土壤有机质、有效磷、速效钾和水解氮含量均呈显著负相关(P<0.05),即适宜的光照、土壤水分和养分含量有利于幼苗生长和物质积累。
3 讨论
3.1 厚朴种子野外萌发的影响因子
种子萌发是植物生活史关键环节之一,种子对萌发条件的响应反映了其适应环境的生态对策,不同物种种子萌发时对土壤水分要求不同。舒枭等[13]研究表明厚朴萌发适宜土壤含水率w为20%~25%,超过30%时种子萌发率开始降低。笔者研究发现针叶林、针阔混交林和阔叶林3种生境内去除假种皮且安装铁丝网处理的厚朴种子萌发率分别为(34.00±5.57)%、(22.67±4.04)%和(3.00±1.73)%,阔叶林厚朴种子萌发率极低可能与阔叶林土壤含水率过高(w为55%)有关。
植物繁殖体在生命周期中总是面临其他各类生物取食的风险,研究发现去除假种皮后,安装铁丝网条件下的厚朴种子平均萌发率(19.89%)高于不安装铁丝网(10.78%),这是由于铁丝网能减少外界动物对厚朴种子捕食、破坏的几率。
相关研究表明,假种皮对种子萌发有一定抑制作用[15],厚朴种子假种皮为红色、肉质,含有较多种子发芽抑制物质,内种皮坚硬,能阻碍种子吸水,影响种子萌发[16]。笔者通过人工去除假种皮和保留假种皮的2种处理模拟野外厚朴土壤种子库,发现保留假种皮的厚朴种子几乎不萌发,这与刘继生等[17]开展的桃叶卫矛(Euonymusbungeanus)种子萌发实验结果基本一致。
3.2 厚朴幼苗野外存活的影响因子
厚朴是耐荫树种,幼苗虽能在林冠下正常生长,但也需要一定阳光条件[18]。从3种不同生境内幼苗存活率和生物量来看,针叶林林分较稀疏,透光率好,幼苗存活率明显高于其他2种,为(29.74±6.25)%。而阔叶林林冠下,枝叶浓荫,林下植物种类多,透光率极低,不利于厚朴幼苗生长,幼苗最终全部死亡。
6—7月厚朴幼苗死亡率达最高,这是因为厚朴抗旱耐热能力较差,只有在较阴凉湿润的环境中才能成功越夏[19],而7月降雨量是影响厚朴生长的主要生态因子之一[20],降雨量过高和过低均能影响厚朴幼苗存活和生长,浙江为亚热带季风气候区,7月降雨量和温度均达最高,对厚朴幼苗存活会产生一定影响。
9月去除假种皮条件下,安装铁丝网的幼苗存活率(29.05%)高于不安装铁丝网(22.68%),这是由于动物对厚朴幼苗存活有一定影响,受动物伤害过的种子(种子胚乳或外壳留有明显咬痕)萌发的幼苗也更容易死亡[21],而铁丝网罩能阻碍动物对厚朴种子和萌芽的捕食及践踏。调查还发现针阔混交林内厚朴幼苗叶片被昆虫取食的现象非常严重,食叶昆虫取食会减少幼苗叶面积或使幼苗落叶,降低幼苗光合作用能力,最终导致幼苗死亡[22]。
3.3 厚朴幼苗野外生长的影响因子
微生境异质主要体现在光照和微气候变化上,局部微气候引起植被下层组成、温度和降雨量变化,对种子萌发、幼苗生长和存活产生不同程度影响[23]。分析表明厚朴幼苗生长与温度、[光]照度呈极显著正相关(P<0.01),即光照充足、温度适宜的环境有利于厚朴幼苗生长。但8月针阔混交林内厚朴幼苗增长量高于针叶林,这是因为较强的光照使幼苗叶片吸收过多光能,光能不能及时有效利用或耗散会使植物遭受强光胁迫,降低光合能力,产生包括耗散过剩光能的光保持机制运转、过剩光能对光合机构的破坏等光抑制效应[24]。吴富勤等[25]研究发现土壤水分和养分含量过低无法满足植物需求,导致幼苗生长缓慢,但土壤水分和养分含量过高会造成根际缺氧而抑制幼苗生长,超过植物忍受阈值甚至会导致烂根死苗。笔者分析表明厚朴幼苗生长与土壤含水率,土壤有机质、有效磷、速效钾和水解氮含量呈显著负相关关系(P<0.05),即过高的土壤含水率、有效磷和速效钾含量不利于厚朴幼苗生长。
植物生物量差异是植物自身和环境因素共同作用的结果,它反映了植物对周身环境不同的适应程度和利用状态[10],不同林型下其微生境不同,针叶林与针阔混交林相比,光照强度更有利于厚朴这类喜光树种的生长,这也是研究中针叶林厚朴幼苗生物量高于针阔混交林的原因之一。生物量的分配在一定程度上反映了树木对环境因子的空间利用情况,是植物适应环境的结果。该研究中厚朴幼苗生物量分配表现为叶>根>茎,与杨志玲等[26]研究发现厚朴种源间苗期生物量分配格局为叶>茎>根的结果有所不同,这是因为微生境对厚朴幼苗生物量分配格局有一定影响[27]。
4 结论
通过设置假种皮(保留和去除)和铁丝网(笼罩和裸露)的不同处理,模拟野外厚朴种子散播机制,观察针叶林、针阔混交林和阔叶林3种不同生境中厚朴种子萌发和幼苗生长状态发现,针叶林最适合厚朴种子萌发和幼苗生长,针阔混交林次之,阔叶林最差。假种皮、动物取食和生境异质性是导致厚朴种子野外萌发率低、死亡率高的主要原因。保护野外厚朴需要加强恢复破碎化生境,建立适合厚朴生长的生态环境,对厚朴生长区进行适当人工管理,减少动物对种子的捕食和幼苗的践踏,提高种子萌发率和促进幼苗健康生长。
猜你喜欢
阅读(科学探秘)(2023年3期)2023-04-10 10:23:27
今日中国·西班牙文版(2021年4期)2021-04-09 11:22:46
西北园艺·果树(2021年1期)2021-02-21 08:21:23
西北园艺(果树)(2021年1期)2021-02-07 02:30:10
——铁丝网丢失记
数学小灵通·3-4年级(2018年11期)2018-11-16 08:22:36
现代农业研究(2018年4期)2018-08-11 10:25:02
中国绿色画报(2018年5期)2018-08-07 07:12:26
森林工程(2018年4期)2018-08-04 03:23:16
现代园艺(2018年2期)2018-03-15 08:01:03
广西林业科学(2016年1期)2016-03-20 05:33:02
