
李斯特菌生长预测模型的研究进展
2018-10-20 06:31:42刘珊娜
天津农学院学报 2018年3期
刘珊娜
李斯特菌生长预测模型的研究进展
刘珊娜1,2
(1. 天津农学院 食品科学与生物工程学院,天津 300384;2. 天津市农副产品深加工技术工程中心,天津 300384)
单核细胞增生李斯特菌是一种重要的食源性致病菌,能够污染多种类型的食品,危害食品安全。利用预测微生物学的研究方法建立微生物生长动力学模型,可对微生物进行风险评估和预测食品货架期。本文介绍了单核细胞增生李斯特菌的生长特点,简述了预测食品微生物学的初级、二级和三级模型的分类,对于单核细胞增生李斯特菌的生长动力学模型在培养基及不同类型食品中的应用进行了总结。
李斯特菌;生长模型;预测食品微生物学
李斯特菌属广泛分布于土壤、水、动植物和人体粪便中,包含单核细胞增生李斯特菌(单增李斯特菌,)、绵羊李斯特菌()、英诺克李斯特菌()、西尔李斯特菌()、威尔李斯特菌()、格氏李斯特菌()等[1]。单增李斯特菌是目前国际公认的重要食源性致病菌,也是我国国标规定的必检项目。单增李斯特菌是一种兼性厌氧的革兰氏阳性短杆菌,无芽孢,可运动。该菌通过污染的食物进入人体后诱发李斯特菌病,致死率高达25%~30%,主要针对免疫功能不全者、孕妇、新生儿和老年人[2]。由于该菌可在低温环境中存活并繁殖,增加了即食食品(Ready-to-eat food)和冷藏食品的食用风险。多种类型的食品都有被该菌污染的报道,欧美国家也曾发生多起该菌引发的食品安全事件[3]。
预测食品微生物学(Predictive food microbiology)将微生物学、数学、统计学与应用计算机知识结合在一起,通过建立数学模型来预测微生物在不同环境条件下的生长、残存和死亡等动态变化过程,为微生物风险评估和食品安全检测提供理论依据。食源性病原菌或腐败菌在食品加工、运输、销售等环节中的变化情况是预测微生物学的重要研究内容。通过建立和应用微生物生长规律模型,预测病原菌或腐败菌实际生长或死亡情况,可提供即时参考数据,分析食品的安全性和货架期。随着预测食品微生物学的广泛应用,预测模型也逐渐扩展到不同类型食品的研究中,指导改进生产工艺和保证质量安全。
1 单增李斯特菌的生长特点
单增李斯特菌最适生长温度在37 ℃左右,培养8 h后进入对数期,在14~18 h到达稳定期[4]。该菌对温度的抵抗力呈现出不对称性,即在低温条件下耐受性较强,在4 ℃的环境中也能生存,而对热的抵抗力较弱。温度会影响菌体生长的延滞期,温度越低,延滞期越长。菌株生长pH值在5.0~9.6之间[5],最适pH值为中性或弱碱性,这也是酸性抑菌的理论基础。当pH值为4.0时,不论温度和盐浓度如何变化,菌株的生长受到完全抑制[6]。单增李斯特菌能够耐受较高的盐浓度。周小红等[4]测定该菌在NaCl浓度为0.5%~3.5%的范围内生长良好,当NaCl浓度高于5.5%时菌株生长受到完全抑制。低剂量接种时(10和103CFU/mL),接种量低的菌液对环境的耐受性高于接种量高的菌液;随着接种浓度的提高(103和105CFU/mL),菌株耐受性增强,具体机制尚未阐明[7]。综合分析,单增李斯特菌的适应能力强,易污染营养丰富的基质,感染病例多由被污染的食品引起。
2 预测模型的分类和特点
动力学模型对微生物生长范围和生长速率进行模拟,分为生长(存活)模型和失活模型。预测模型可分为初级模型(Primary model)、二级模型(Secondary model)和三级模型(Tertiary model)。初级模型用来描述特定条件下微生物量随时间变化的情况,常用的有Gompertz模型、Logistic模型、Richards模型、Baranyi & Roberts模型(Baranyi模型)和Huang模型等,可得到延滞期、最大比生长速率等参数;二级模型反映一级模型中各参数如微生物最大比生长速率与环境因子间的关系,包括响应面方程(Response surface equation)、平方根方程(Square root model)和人工神经网络等;三级模型是在初级模型和二级模型的基础上组建数据库,用于直接预测的计算机软件或程序如Pathogen Modeling Program(PMP)、Food spoilage predictor、Combined database (ComBase)和Seafood Spoilage and Safety Predictor(SSSP)。
2.1 Gompertz模型
Gompertz方程式为双指数函数,表达式[8]为:
=+×exp{-exp[-(-)]}
是微生物在时间时常用对数值;是随时间无限减小时渐进对数值;是随时间无限增大时菌增量的对数值;是时的相对最大生长速率;是达到相对最大生长速率所需时间。
Gompertz模型使用简单,修正后的Gompertz模型为经验模型,是描述微生物生长所常用的初级模型之一。有报道指出,其在拟合鲜切生菜中沙门氏菌生长曲线时的相关系数随着储藏温度的下降而减小[9]。可能在于低温条件不在菌株最佳生长温度范围,曲线偏离“S”型而影响拟合效果。
2.2 Logistic模型
Logistic模型曲线具有断点,适用于菌株生长延滞期预测,Tienungoon等[10]采用非线性Logistic模型计算单增李斯特菌在不同温度、pH、盐浓度等培养基中的生长极限。王旭等[11]应用修正的Logistic方程拟合了37 ℃下单增李斯特菌在pH 5~9的培养基中的生长曲线,相关性大于0.99。该模型表达式[8]为:
=/{1+exp [4m(-)/+2]}
是微生物在时相对菌数的常用对数值,即Logt0;是相对最大菌浓度,Logmax0;m是生长速率;是延滞期。
2.3 Baranyi模型
Baranyi模型是根据微生物的生长代谢机理推导动力学方程,既能用于描述致死曲线也适用于动态环境,对延滞期的估计和曲线拟合较准确。马晨等[12]利用Baranyi模型拟合沙门氏菌在鲜切黄瓜中的生长情况,并建立了具有可靠性的Ratkowsky二级模型来描述最大生长速率与储藏温度之间的关系,为评估该菌在鲜食蔬菜中的生长情况提供依据。Baranyi模型表达式[13]如下:
=min+(0-min)exp {-max[-()]}
()=∫t0[r/r+s]d
是时菌数;0是0时菌数;min是最小菌数;max是最大相对死亡率;、是参数。
2.4 Huang模型
Huang模型属于三阶段线型模型,反映了恒温条件下微生物生长经历的延滞期、对数期和稳定期,有明显的延滞期。表达式[14]如下:
=0+max-ln{exp(0)+[exp(max)-exp(0)] exp [-×()]}
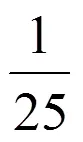
0和max分别为的初始值和最大值;为生长速率,为延滞期。
2.5 二级模型和三级模型
二级模型中包括两大类:一类是Ratkowsky的平方根模型和参数模型,另一类是神经网络模型和响应面模型。平方根模型建立了微生物生长速率与温度平方根之间的线性关系,通过温度变化考察微生物生长情况。其中Ratkowsky模型使用较多,能很好预测单一因素对最大比生长速率的影响,特别是温度的影响。Arrhenius模型是在酶促反应的基础上推广至多参数条件下微生物生长的描述。响应面模型通过确定试验和回归方程推导出未知参数值,提高拟合度和准确性。

表1 二级模型的种类和表达式
目前已开发出多种预测模型软件包括Growth Predictor、PMP、SSSP、Sym’ Previus、ComBase等,提供可靠性预测数据。这种基于三级模型的数据库系统能够比较不同环境因子对微生物生长的影响及相同环境因子对不同微生物生长的影响。虽然用于预测微生物生长的模型有多种,但目前并不能确定哪种模型具有明显的优越性,仍需通过实际培养微生物的数据来评价不同模型的拟合能力。
3 单增李斯特菌生长预测模型的应用
3.1 在培养基中的应用
目前,已建立单增李斯特菌在培养基中的预测模型,菌株随温度升高,生长速率变快;相同温度下初始菌浓度越大,最大比生长速率越大。丁甜等[16]使用Gompertz模型建立了不同温度下单增李斯特菌在营养肉汤中的生长模型,在此基础上建立的二级模型中,Ratkowsky模型在精确性上优于Linear和Square root模型,可更好预测不同温度下菌株的最大比生长速率。此外,Logistic回归模型用于单增李斯特菌生长/非生长条件的研究,反映了环境因子如温度、pH、有机酸等的交互作用对菌株生长的影响[6]。研究结果为优化生产工艺和提高产品的安全性提供了参考依据。
3.2 在肉制品中的应用
与培养基相对稳定的体系相比,食品原料对微生物的影响更为复杂。肉制品是极易发生腐败变质的食品类型,特别是在冷鲜肉中单增李斯特菌的污染在国内外都有报道。由于Gompertz模型形式相对简单,具有明确的生物学意义,已用于建立单增李斯特菌在肉制品中的生长预测模型[17],通过测定初始菌数,判断不同时间点的李斯特菌数。通过将Baranyi模型与改良Lotka-Volterra模型结合,建立了单增李斯特菌与假单胞菌的竞争生长模型[18]。
3.3 在蔬菜制品中的应用
通过Gompertz、Logistic和Baranyi的初级模型,可建立在鲜食蔬菜、凉拌菜中单增李斯特菌的生长模型[5, 15],并根据最大比生长速率建立二级模型,结合ComBase数据库得到三级模型,用于描述李斯特菌菌株在不同温度下的生长状态,经验证,较好预测了该菌在相应环境中的生长情况。Fang等[19]利用Logistic模型结合不同二级模型预测了4~43 ℃温度条件下单增李斯特菌在鲜切哈密 瓜上的生长情况。Danyluk等[20]通过Baranyi和Ratkowsky模型拟合了单增李斯特菌在鲜切哈密瓜、香瓜和西瓜中的生长情况,分析了温度与菌株生长的关系,揭示了低温环境下菌株生长量的持续增加。
3.4 在乳制品中的应用
李斯特菌在乳制品中的检出率较高,乳中丰富的营养给李斯特菌的生长提供了条件。特别是在低温储藏条件下,嗜冷菌成为原料乳中的优势微生物。例如美国近年就曾发生由巴氏杀菌乳、冰激凌和干酪污染引发的食品安全事件[21-22]。邱红玲等[23]利用Gompertz模型建立了原料乳或复原乳中单增李斯特菌的生长模型,Tiwari等[24]也建立了单增李斯特菌在干酪中的初级生长预测模型,可用于保障乳制品的质量安全。
3.5 影响模型准确性的因素
对李斯特菌在不同环境条件下的初级生长模型研究已展开,但是预测效果受到多重因素的影响。特别是食品原料、加工工艺和储藏环境的多样性,给预测微生物带来不确定性。在设计试验时首先要从现实情况出发,尽量排除人为干扰。此外要综合考虑整个食品加工过程,而不仅仅是某种特定条件,克服局限性。对于混合微生物的样品,不应忽视微生物之间的竞争或促进关系。因此多因素条件下微生物预测模型的建立将是重要研究方向。此外,为提高预测准确性,可采用多种模型拟合,不限定在单一模型。选择合适的模型评价方法,通过偏差因子、准确因子和均方根误差等指标进行可靠性分析,最终筛选最优模型。在软件方面,主要应用国外已有的预测模型和开放的数据库资源[25],还应加强预测模型的开发和相关数据库的建立,以适应多种环境因素。将多种生物学技术例如Real-time PCR、DNA杂交探针等应用于预测微生物学,也为综合评价模型的准确性提供了更多途径。
4 结论
李斯特菌在自然界广泛存在,可造成多种食品的污染,给食品安全带来潜在的危害。目前,李斯特菌病的中毒诊断方法尚未建立,仍需完善该病爆发和传播的相关数据。随着预测微生物学的发展,食源性致病菌在各种食品基质中的生长动力学模型逐渐建立,但是对影响其准确性的因素研究还不完善。只有不断对数据进行挖掘,建立健全预测系统,才有利于评估李斯特菌在食品加工和储运体系中的风险系数和制订有效的食品质量安全控制方案。
[1] Kuenne C,Billion A,Mraheil M A,et al. Reassessment of thepan-genome reveals dynamic integration hotspots and mobile genetic elements as major components of the accessory genome[J]. BMC Genomics,2013,14(1):47.
[2] Winkelströter L K,De Martinis E C. Effect of bacteriocins and conditions that mimic food and digestive tract on biofilm formation,in vitro invasion of eukaryotic cells and internalin gene expression by[J]. Probiotics & Antimicrobial Proteins,2013,5(3):153-164.
[3] Martinez-Rios V,Dalgaard P. Prevalence of,in European cheeses:A systematic review and meta-analysis[J]. Food Control,2018,84:205-214.
[4] 周小红,李学英,杨宪时,等. 培养条件对单增李斯特菌生长的影响[J]. 食品工业科技,2013,34(15):140-143,148.
[5] 郑丽敏. 即食凉拌菜中单增李斯特菌的风险评估与管理[D]. 上海:上海理工大学,2012.
[6] 周小红,李学英,杨宪时,等. 环境因子及接种量对单增李斯特菌生长/非生长界面的影响[J]. 食品工业科技,2015,36(7):53-59.
[7] 周小红,李学英,杨宪时,等. 接种量对单增李斯特菌生长期及生长界面的影响[J]. 食品工业科技,2014,35(18):180-184.
[8] 蒲月华. 纳豆菌抗菌肽对对虾中金黄色葡萄球菌和单增李斯特菌的控制效应及数学模型建立[D]. 湛江:广东海洋大学,2013.
[9] 朱奚慕. 鲜切生菜中沙门氏菌生长动力学预测模型的建立和验证[D]. 雅安:四川农业大学,2014.
[10] Tienungoon S,Ratkowsky D A,Mcmeekin T A,et al. Growth limits ofas a function of temperature,pH,NaCl,and lactic acid[J]. Applied & Environmental Microbiology,2000,66(11):4979-4987.
[11] 王旭,孙晓红,潘迎捷,等. 单增李斯特菌及其缺失突变株在不同pH下生长动力学的比较[J]. 食品与生物技术学报,2016,35(5):477-484.
[12] 马晨,李建国,程景. 沙门氏菌在鲜切蔬菜中生长行为及其预测模型构建[J]. 食品工业科技,2016,37(4):123-127.
[13] 周康,刘寿春,李平兰,等. 食品微生物生长预测模型研究新进展[J]. 微生物学通报,2008,35(4):589-594.
[14] 牛会敏,李苗云,赵改名,等. 低温条件下冷却猪肉中假单胞菌生长模型的比较分析[J]. 食品科学,2013,34(5):142-146.
[15] 张引成. 单增李斯特菌在鲜切结球莴苣中生长和抑菌预测模型的建立[D]. 南京:南京农业大学,2012.
[16] 丁甜,董庆利,王璐,等. 单增李斯特菌在营养肉汤中最大比生长速率的预测模型[J]. 华中农业大学学报,2010,29(4):522-526.
[17] 高继铭. 酱卤肉中代表性致病菌预测微生物学研究[D]. 哈尔滨:东北农业大学,2014.
[18] 陈尔凝,白羽,马凯,等. 猪肉中单增李斯特菌与假单胞菌的竞争生长预测模型建立[J]. 食品科学,2013,34(23):256-260.
[19] Fang T,Liu Y,Huang L. Growth kinetics ofand spoilage microorganisms in fresh - cut cantaloupe[J]. Food Microbiology,2013,34(1):174-181.
[20] Danyluk M D,Friedrich L M,Schaffner D W. Modeling the growth ofon cut cantaloupe,honeydew and watermelon[J]. Food Microbiology,2014,38(4):52-55.
[21] Cumming M,Kludt P,Matyas B,et al. Outbreak ofinfections associated with pasteurized milk from a local dairy-Massachusetts,2007[J]. Journal of the American Medical Association,2009,301(8):1097-1100.
[22] 张亚红,王娉,杨海荣,等. 预测微生物学在乳及乳制品中的应用[J]. 检验检疫学刊,2015,25(6):62-65.
[23] 邱红玲,唐晓阳,王李伟,等. 复原乳中三种常见致病菌的生长动力学模型构建[J]. 上海预防医学,2016,28(5):288-293.
[24] Tiwari U,Walsh D,Rivas L,et al . Modelling the interaction of storage temperature,pH,and water activity on the growth behaviour ofin raw and pasteurised semi-soft rind washed milk cheese during storage following ripening[J]. Food Control,2014,42(3):248-256.
[25] 刘静,杜广全,管骁. 食品预测微生物软件的分析与比较[J]. 食品与机械,2016,32(4):61-66,70.
责任编辑:张爱婷
Research progress in predictive models forgrowth
LIU Shan-na1,2
(1. College of Food Science and Bioengineering, Tianjin Agricultural University, Tianjin 300384, China; 2. Tianjin Engineering and Technology Research Center of Agricultural Products Processing, Tianjin 300384, China)
is an important food-borne pathogen which may contaminate many types of food and impair food safety. Research methods of predictive food microbiology can be used for building growth kinetic models, which may assess microbial risk and predict food shelf life. This article introduced growth characteristics of. The classification of primary, secondary and tertiary models of predictive food microbiology were briefly reviewed. Applications ofgrowth kinetic models in culture medium and different foods were summarized.
; growth model; predictive food microbiology
1008-5394(2018)03-0092-04
10.19640/j.cnki.jtau.2018.03.020
Q939.99
A
2018-02-26
国家自然科学基金青年项目(31501583)
刘珊娜(1984-),女,讲师,博士,主要从事食品微生物研究。E-mail:lsn_nxy@aliyun.com。
猜你喜欢
生命与灾害(2022年8期)2022-11-10 05:14:26
疯狂英语·初中天地(2022年1期)2022-07-07 08:37:54
党员文摘(2022年1期)2022-04-03 21:37:19
湖北农机化(2020年4期)2020-07-24 09:07:26
幼儿园(2019年7期)2019-09-05 17:49:18
上海农业学报(2017年4期)2017-04-10 12:40:24
河南科技(2014年14期)2014-02-27 14:12:22
小小说月刊(2013年6期)2013-05-14 14:55:19
食品科学(2013年24期)2013-03-11 18:30:42
中国科技术语(2012年3期)2012-03-20 14:36:15
