
躅蝽标记信息素释放规律研究
2018-10-17 02:45祁宁,吴惠惠,刘海学
吉林农业 2018年20期
祁宁,吴惠惠,刘海学

摘要:对捕食性天敌昆虫躅蝽标记信息素释放规律进行观察研究,结果表明,躅蝽会释放标记信息素标记它搜索过的区域,并且只有雌虫使用标记信息素,标记信息素的有效性取决于雌虫的发育时期和被搜寻的标记信息素的质量。雌虫标记信息素化学成分尚未确定,但其活性期明显较短。
关键词:躅蝽;标记信息素;释放规律
中图分类号: S476 文献标识码: A DOI编号: 10.14025/j.cnki.jlny.2018.20.033
躅蝽(Arma chinensis)可以捕食棉铃虫(Helicoverpa armigera)、三点盲蝽(Adelphocoris fasciaticollis)、绿盲蝽(Apolygus lucorum)(Zou et al.,2012)、双斑长跗萤叶甲(Monolepta hieroglyphica)(陈静等,2007)、马铃薯甲虫(Leptinotarsa decemlineata)(郭文超等,2011;Zou et al.,2013)和美国白蛾(Hyphantria cunea)(王文亮等,2012)等鳞翅目、鞘翅目及半翅目的多种害虫,是农林业上一种重要的捕食性天敌昆虫。多年以前,人们便注意到捕食性天敌对害虫的控制作用随天敌本身或害虫的密度增加而下降。近年来,我国学者发现,随着七星瓢虫密度的增加,对狭冠网蜻和桃蚜叫的捕食效率逐渐下降。相似的现象也在龟纹瓢虫、异色瓢虫、多异瓢虫、中华草蛉、叶色草蛉等捕食性天敌中发现。这种负密度效应由捕食者本身而不是猎物引起。近十余年的研究表明,捕食者之间的忌避作用常由标记信息素介导。目前我国在昆虫标记信息素,尤其是捕食性天敌昆虫标记信息素研究领域尚处于一片空白。本文试图从标记与识别,以及辨识标记留下的标记信息素入手,去揭示躅蝽对害虫的控制作用随天敌本身或害虫的密度增加而下降的内在本质。
1 材料与方法
1.1 供试虫源
本研究所用的躅蝽种群源自天津市植物保护研究所。躅蝽种群用柞蚕蛹饲喂,条件为27 ±1℃,16:8 (L:D)及75 ± 5% RH。在室内稳定扩繁4代后用于本研究。收集的躅蝽卵块放于培养皿(D = 9cm)内用蒸馏水喷湿的滤纸上。每天向滤纸喷施一次蒸馏水以保持一定的湿度。卵大约6d后孵化。将脱脂棉团用蒸馏水浸湿后放于310mL透明的塑料杯底部,之后将新孵化的一龄若虫转于湿脱脂棉上。3d后孵化为二龄若虫。
1.2 食物准备
柞蚕蛹购于天津市红旗农贸市场,于室内4℃冰箱存放。分别将150头二龄躅蝽若虫放于300mL纸杯内,用纱网盖住以防逃逸,单头饲喂。每个若虫饲喂一个柞蚕蛹并将用蒸馏水浸湿的脱脂棉放于网上以提供水分。蒸馏水每天加一次,柞蚕蛹每7d~12d换一次。这种饲养方法一直用到成虫。每天记录蜕皮情况。
成虫羽化后于450mL纸杯中配对饲养,每杯一对。用双面胶将6cm×1.5cm的玻璃管粘于纸杯底部,管内注水后将两棵大豆苗放于管内。玻璃管内的水每隔2d~3d加一次。每7d~12d更换一次蚕蛹。
1.3 标记与识别
所有实验都在一个实验平台进行,玻璃培养皿(直径6.0cm,高1.5cm),通过插入纸路障(6.0×1.5cm)分为相等的两半。一半作为处理区域,允许躅蝽成虫在此区域自由行动30分钟;另一半作为对照区,不允许躅蝽进入。30min后移除躅蝽成虫和纸路障,准备用这个培养皿进行单次的生物测定。
使用这个培养皿设计进行了许多实验。实验1目的是确定躅蝽个体是否会对自己的标记信息素和特定个体的标记做出回应。测试前者,使用细刷子将同一躅蝽个体,立即重新引入撤除纸路障的培养皿中心。
为了测试后者,立即释放一个不同的躅蝽个体进入撤除纸路障的培养皿中心。观察所有测试个体10min,确定其在对照区或处理区的时间。分别以雄虫、未经交配的雌虫和交配过的雌虫这个试验。另外,为了确保标记信息素的活性,试验进行1h后,重新引入之前用于领域标记的那只躅蝽爬行10min,释放标记信息素。试验1,重复20次。所有参与试验的躅蝽个体在试验前均需饥饿处理24h。
试验2的目的是研究躅蝽雌虫是否会避免重复搜索相同的领域,以及食物的存在是否会影响雌虫捕食者释放的标记信息素的质量。试验引入棉铃虫的二龄幼虫。本实验中使用的猎物幼虫通过-20℃冷冻致死,然后在25℃下保持数小时。12只猎物被放置在实验场一边。释放饥饿24h的交配过的躅蝽雌虫到试验场的另一边。30min后,路障和所有的猎物被移走,同样的雌虫被释放到竞技场的中心。观察到雌虫行为,试验重复20次。
试验3的目的是确定饥饿状态的影响标记信息素的利用率。试验开始前,每天用30只猎物的幼虫饲喂雌虫。除此之外,试验方法与试验1相同,试验重复20次。
1.4 数据处理
使用Wilcoxon符号秩检验分析(0.05水平)成虫在试验平台培养皿每一侧的提留时间。
2 试验结果分析
2.1 试验1
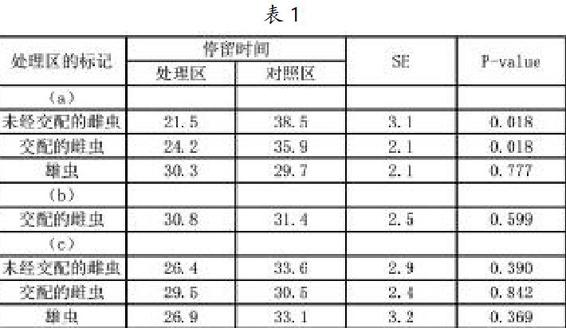
饑饿处理后雌虫在对照区的停留时间为明显长于处理区,在交配过和未经交配的雌虫之间,n = 20,Wilcoxon符号秩检验,z = 2.36,P <0.05交配过的雌虫;z = 2.36,P <0.05未经交配的雌虫;表1。饥饿处理后雄虫在对照区和处理区的停留时间在统计学上无差异,n=20,Wilcoxon符号秩检验,z= 0.27,P = NS;表1(a)。这些结果表明,饥饿状态下的雌性躅蝽会避免它们之前搜索过的区域,而雄性躅蝽不会。
与雌性成虫对自身标记信息素的反应相反,无论是经过交配的雌虫还是未经交配的雌虫对其他同种个体标记信息素均没有趋避反应,n=20,Wilcoxon符号秩检验,z=0.19,P=NS为交配过的雌虫;z = 0.85,P = NS为未经交配的雌虫。 雄性成虫也没有避免其他同种标记信息素的反应,n = 20,Wilcoxon符号秩检验,z = 0.89,P =NS;表1(c)。
移除饥饿的交配过的雌虫1h后,同一只雌虫在对照区和处理区停留的时间没有不同,n=20,Wilcoxon符号秩检验,z=0.51,P=NS;表1(b)。
2.2 试验2
引入猎物并不会影响雌虫释放的标记信息素的质量,n= 20,Wilcoxon符号秩检验,z=0.25,P=NS;表2(a)。
2.3 试验3
与试验1的结果不同,不处于饥饿状态的雌虫在对照区和处理区的停留时间没有区别,n=20,Wilcoxon符号秩检验,z=1.67,P=NS;表2(b)。
3 讨论
上述实验结果表明,天敌昆虫躅蝽会释放标记信息素标记其搜索过的区域,但只有雌虫使用标记信息素。并且這种标记的有效性取决于两个因素:雌虫的发育时期和被搜寻的标记信息素的质量。被剥夺食物的雌性捕食者在以前搜索过的食物缺乏地区的时间要比在新遇到的区域少得多。因此,雌虫可以区分先前搜索过的区域和之前没有搜索过的区域,分别通过雌虫自身释放一种标记化学物质来实现的。该标记信息素化学成分尚未确定,但其活性期明显较短。由于活动时间很短,雌性动物所留下的印记是它们最近访问过的食物不足地区的可靠指标。
昆虫标记信息素主要影响其自身的行为,有些也同种其他个体的行为。Nakashima等发现东亚小花蝽雌成虫释放标记信息素标记已搜索过的区域,为了避免重复,节省时间和能量。Gil等在温室研究了异色瓢虫成虫受同种幼虫标记信息素影响而扩散的问题,在温室植株上无同种幼虫时,成虫随机移动。当在温室的一半植株上释放同种幼虫时,成虫明显迁移到另一半没有释放幼虫的植株上。昆虫也可以利用标记信息素避免来自集团内的伤害,可以根据标记信息素宣告本身的存在,使其他捕食者远离而保证自身安全和猎物资源。信息素的接受者借此信号评估潜在的捕食者,避免在具有强势捕食者的区域取食和产卵,保证了自身安全和种群延续。
进一步提取和鉴定躅蝽标记信息素,分析各组分间的最佳配比,验证躅蝽标记信息素组分及配比的准确性。利用躅蝽标记信息素来修饰躅蝽行为,提高天敌昆虫对害虫的控制能力和效率值得深入研究。
参考文献
[1]Zou DY, et al. Taxonomic and bionomic notes on Arma chinensis (Fallou) (Hemiptera: Pentatomidae: Asopinae). Zootaxa, 2012,3328:41-52.
[2]Zou DY, et al. 2013. A meridic diet for continuous rearing of Arma chinensis (Hemiptera: Pentatomidae: Asopinae). Biological Control,2013,67(03):491-497.
[3]陈静,等,2007.敌对双斑长跗萤叶甲成虫的捕食功能研究.昆虫天敌,29(04):149-154.
[4]郭文超,等,2011.马铃薯甲虫持续防控技术研究与应用. 新疆农业科学,48(02):197-203.
[5]王文亮,等,2012.美国白蛾新天敌-敌捕食能力的初步观察.山东林业科技,1:1-14.
[6]Nakashima Y, Teshiba M, Hiorse Y. Flexible use of patch marks in an insect predator: effect of sex, hunger state, and patch quality. Ecological Entomology,2002,27(05):581-587.
作者简介:祁宁,在读本科生,研究方向:园林园艺。
通讯作者:吴惠惠,博士,助理研究员,研究方向:昆虫信息素。
