
川西獐牙菜SmDL7H基因原核表达及组织表达
2018-10-16 05:50:22李晓雪孙继奇朱晔荣向蓓蓓
西北植物学报 2018年8期
李晓雪,王 勇,孙继奇,朱晔荣,马 琳,向蓓蓓*
(1 南开大学 生命科学学院,天津 300071;2 天津中医药大学 中药学院,天津 300193)
川西獐牙菜(SwertiamussotiiFranch)是龙胆科獐牙菜属植物,俗称“藏茵陈”,藏名“斗大”、“桑蒂”。主要生长分布在青海、西藏、四川等海拔3 800 m以上的高寒地区。全草入药,是藏药中极具特色的治疗热症、肝胆病及血液病的常用药物。具有清热利胆、除湿、舒肝健胃、强心、养血、解毒等作用[1]。现已生产片剂、针剂等多种剂型的药物应用于临床治疗黄疸型肝炎、病毒性肝炎,血病等疾病[2]。
其主要有效成分为裂环烯醚萜类(獐牙菜苦苷、獐牙菜苷、龙胆苦苷等)、三萜类(齐墩果酸、乌苏酸等)、黄酮类等化合物[3]。裂环烯醚萜化合物是一类特殊的单萜化合物,其苷元结构特点为母核中环戊烷的C7-C8处断键成裂环状态。在植物界中此类化合物是由焦磷酸香叶酯(GPP)经生物合成途径生成臭蚁二醛再缩醛衍生而成,广泛分布于木犀科、茜草科、龙胆科、唇形科等双子叶植物中[4-5]。裂环烯醚萜在生物合成中常是吲哚生物碱的前体物质,研究表明,一些裂环烯醚萜类化合物对中枢神经、心血管和消化系统都有一定的生物活性[6]。川西獐牙菜中7-脱氧马钱子酸羟化酶(7-deoxyloganic acid-7-hydroxylase, SmDL7H)是其裂环烯醚萜类化合物合成途径的关键酶,该酶属于细胞色素p450氧化酶家族中的一员,是一种依赖于NADPH与氧分子的酶,能够催化7-脱氧马钱子酸的C-7位发生羟基化生成马钱子酸,马钱子酸再经过一系列的催化反应生成具有药理活性的龙胆苦苷、獐牙菜苦苷、獐牙菜苷等裂环烯醚萜类化合物[7-9]。7-脱氧马钱子酸羟化酶目前仅在长春花[7]、金银花[8]等少数植物中有对其功能进行研究的报道。
本研究依据实验室前期工作获得川西獐牙菜转录组,从中首次获得了川西獐牙菜7-脱氧马钱子酸羟化酶基因(SmDL7H)的全长cDNA序列。设计特异性引物,通过同源克隆得到了该基因。利用生物信息学方法,对其与其他植物中的DL7H基因进行分析比对,并构建进化树,分析同源关系,同时对其结构域以及信号肽等信息作出了预测分析。构建原核表达载体pET-28a-SmDL7H,转入大肠杆菌Rosetta(DE3)中进行原核表达,得到与预期蛋白大小一致的目的蛋白。利用荧光定量PCR对川西獐牙菜不同组织中SmDL7H基因的表达进行分析,结果发现SmDL7H基因在川西獐牙菜叶、茎、花、根、愈伤组织中均有表达,其中在叶片中表达量最高,在根中表达量最低。本研究为研究川西獐牙菜中的7-脱氧马钱子羟化酶的功能,以及为探索川西獐牙菜中裂环烯醚萜类化合物合成途径奠定了基础,同时也为其他近缘植物研究该类化合物生物合成途径提供了研究依据。
1 材料和方法
1.1 材 料
实验材料为在实验室中光照16 h、黑暗8 h、22 ℃培养种植的生长8个月已抽薹开花的川西獐牙菜(SwertiamussotiiFranch)植株。
1.2 方 法
1.2.1叶片总RNA提取及cDNA合成按照Eastep Super总RNA提取试剂盒说明书进行川西獐牙菜叶片总RNA的提取,用1.1%琼脂糖凝胶电泳和Nanodrop2000对RNA质量进行检测。根据Reverse Transcriptase M-MLV(RNase H-)试剂盒说明书对叶片总RNA反转录合成cDNA第一链,保存于-80 ℃备用。
1.2.2SmDL7H基因的克隆根据实验室前期工作,从川西獐牙菜转录组中获得了SmDL7H基因的全长cDNA序列,并结合pET-28a原核表达载体的酶切位点,利用Primer 5.0软件设计特异性引物7B UP(5′CCGGAATTCATGGGAATAAGCTTAGAGACC 3′)和 7B DP (5′ACGCGTCGAC-GAGTTTGTGCAATATCAAGTGA 3′,划线部分为酶切位点序列)。以川西獐牙菜叶片cDNA第一链为模板进行扩增,扩增条件为:95 ℃预变性2 min;95 ℃变性20 s,58 ℃复性20 s,72 ℃延伸90 s,31个循环;72 ℃终延伸5 min。获得的PCR产物用1%琼脂糖凝胶电泳进行鉴定,按照快速琼脂糖凝胶DNA纯化回收试剂盒的说明书进行切胶回收。回收目的片段与pEASY-Blunt Simple Cloning Vector按照说明书进行连接。转化EscherichiacoliTrans 5α,挑取阳性克隆送华大基因公司进行测序。
1.2.3SmDL7H基因序列的生物信息学分析利用NCBI网站,分析比对川西獐牙菜SmDL7H基因与其他植物中的DL7H基因的同源性及结构域;使用DNAMAN软件对川西獐牙菜SmDL7H与其他植物中的DL7H的氨基酸序列进行比对;用ExPASy Proteomics Server(http://www.expasy. ch/tools/protparam.html)网站对川西獐牙菜的SmDL7H基因编码的蛋白进行理化性质的分析。使用SignalP4.0 Server(http:// www.cbs.dtu.dk/services/SignalP/)在线网站进行信号肽的预测分析。用MEGA 7.0软件构建Neighbor-joining系统进化树,Bootstrap 1 050次。
1.2.4SmDL7H基因的原核表达载体的构建将测序正确的pEASY-Blunt Simple-SmDL7H与pET-28a原核表达载体,用EcoR I和SalI限制性内切酶进行双酶切。将酶切产物经过切胶回收和连接后,将连接产物转入EscherichiacoliRosetta(DE3)中,选取阳性转化子,送去华大基因公司进行测序。
1.2.5SmDL7H基因的原核表达挑取测序正确含有pET-28a-SmDL7H重组质粒的Rosetta(DE3)的单菌落,接种于LB(含有50 μg/mL卡那霉素)液体培养基中,于37 ℃、200 r/min进行小量培养12 h。至菌OD为0.6~0.8时接种1 mL于400 mL LB(含有50 μg/mL卡那霉素)液体培养基中,25 ℃、8 h扩大培养,至菌OD为0.6~0.8时加入0.1 mol/L IPTG,25 ℃进行诱导12 h。将诱导好的菌液进行5000 r/min、4 ℃、10 min离心,收集菌体弃去上清液。按照1∶10比例用Binding buffer(0.5 mmol/L NaCl;20 mmol/L 咪唑;20 mmol/L 磷酸钠)将菌体重悬后,进行超声破碎10 min。破碎后的液体,进行5 000 r/min、4 ℃、10 min离心,收集上清液弃去沉淀。取8 μL上清液与8 μL的2×上样缓冲液均匀混合后,取8 μL用SDS-PAGE(4.5%浓缩胶和12.5%的分离胶)进行电泳检测。
1.2.6川西獐牙菜叶片诱导愈伤组织剪取生长8个月已抽薹开花的川西獐牙菜的叶片,用10%次氯酸钠消毒7 min,用灭菌蒸馏水清洗5次后,放置于添加了3 mg/L 2,4-D和0.5 mg/L 6-BA的MS固体培养基上。放置于培养架光照16 h、黑暗8 h、22 ℃进行培养30 d。选取生长良好的愈伤组织放置于继代培养基(MS固体培养基+2 mg/L 6-BA),于培养架光照16 h、黑暗8 h、22 ℃进行培养15 d,获得长势良好的愈伤组织。
1.2.7SmDL7H基因表达分析取生长8个月已抽薹开花的川西獐牙菜的叶、茎、花、根,以及用叶片诱导生长45 d的川西獐牙菜愈伤组织。提取RNA与反转cDNA第一条链的方法同上。将cDNA稀释5倍用作qRT-PCR的模板,选用actin基因作为内参。按照SYBR Premix Ex Taq II的说明书设计引物,设计的引物为SmDL7HqRT-PCR up (5′ GTAAGTAAATGGGAGGAGAAGGT3′)和SmDL7HqRT-PCR dp (5′ CAGGGATGTGAACAGAACGAC-3′);actinup (5′ ACTGGTGTTATGGTTGGTATGG-3′)和actindp (5′ TCGGTGAGAAGTATAGGGTGC-3′)。qRT-PCR的25 μL反应体系为:SYBR Premix Ex TaqⅡ 12.5 μL;灭菌蒸馏水9.5 μL;上游引物0.5 μL;下游引物0.5 μL;cDNA 2 μL。qRT-PCR反应在iCycler Thermal Cycler的仪器上进行,反应程序为:95 ℃预变性30 s;95 ℃变性5 s,58 ℃复性30 s,72 ℃延伸30 s,40个循环。用actin作为内参,使用2-ΔΔCT方法来进行基因表达量的确定,其中参照物的表达量设置为“1”。
2 结果与分析
2.1 川西獐牙菜SmDL7H基因的克隆
以川西獐牙菜叶片的cDNA为模板,使用特异性引物进行PCR扩增,用1%琼脂糖凝胶电泳检测,在1 554 bp处有特异性片段(图1)。将该特异性片段连接在pEASY-Blunt Simple Cloning Vector送去华大基因公司进行测序。经过序列比对及生物信息学方法分析,获得的片段与我们预测分析的序列一致(图2),即为川西獐牙菜SmDL7H基因,GenBank登录号为MH243070。
2.2 川西獐牙菜SmDL7H基因的理化性质分析
SmDL7H基因编码蛋白理化性质预测与分析显示,SmDL7H基因共编码的517个氨基酸,等电点为9.02,蛋白分子量为59.5 kD,59个带负电荷的氨基酸残基,68个带正电荷的氨基酸残基,分子式为C2708H4257N715O747S23。稳定性指数为37.5,脂溶系数为91.82,亲水性平均数为-0.169,说明该蛋白为一个相对稳定的水溶性蛋白。

M. DL2000; 1.阴性对照; 2. SmDL7H 图1 SmDL7H基因的PCR扩增M. DL2000; 1. Negative control; 2. SmDL7HFig.1 PCR amplification of SmDL7H gene

图2 川西獐牙菜SmDL7H基因核苷酸序列与推测的氨基酸序列Fig.2 The nucleotide and putative amino acid sequences of SmDL7H gene from Swertia mussotii Franch

图3 SmDL7H蛋白的保守功能域Fig.3 The conserved domains of SmDL7H protein
2.3 川西獐牙菜SmDL7H蛋白的保守功能域与信号肽的推测分析
川西獐牙菜SmDL7H基因编码氨基酸序列,与其他植物中DL7H基因编码的氨基酸序列, Blast对比结果(图3)表明,川西獐牙菜SmDL7H编码的蛋白属于p450超家族。信号肽预测结果(图4)表明,川西獐牙菜SmDL7H编码的蛋白不含信号肽。
2.4 川西獐牙菜SmDL7H的聚类分析
为了了解川西獐牙菜SmDL7H基因的功能,利用MEGA 7.0软件对川西獐牙菜SmDL7H基因编码的氨基酸序列与NCBI上已公开的滇龙胆(AKI87771.1)、印度蛇根草(AGX93059.1)和小蔓长春花(AGX93061.1)等的DL7H基因编码的氨基酸序列进行比对,构建Neighbor-joining系统进化树。结果(图5)显示,川西獐牙菜编码的SmDL7H蛋白与滇龙胆编码的DL7H蛋白共聚一支,证明二者具有更近的亲缘关系。同时利用DNAMAN软件将滇龙胆、印度蛇根草和小蔓长春花等的DL7H氨基酸序列,与川西獐牙菜中的SmDL7H的氨基酸序列进行比对,结果(图6)显示,川西獐牙菜中的SmDL7H蛋白与多个物种的DL7H蛋白具有较高的相似性。
2.5 川西獐牙菜SmDL7H基因原核表达
将pET-28a原核表达载体与pEASY-Blunt Simple-SmDL7H质粒用EcoR I和SalI进行双酶切,得到SmDL7H基因全长序列与酶切后的pET-28a原核表达载体。将SmDL7H基因全长序列与酶切后pET-28a原核表达载体用T4连接酶进行连接,转入大肠杆菌Rosetta(DE3)中。将pET-28a原核表达载体转入Rosetta(DE3)中作为阴性对照,简称为Rosetta-pET-28a。将Rosetta-pET-SmDL7H与阴性对照用相同的条件进行诱导表达,将表达后的Rosetta-pET-28a和Rosetta-pET-SmDL7H用SDS-PAGE进行检测。在59.5 kD处有目的蛋白的出现(图7),与之前预测的蛋白大小是相一致的。

图4 川西獐牙菜SmDL7H基因编码的蛋白信号肽预测分析Fig.4 Prediction of signal peptide of SmDL7H protein encoding by SmDL7H gene

图5 川西獐牙菜SmDL7H蛋白进化树分析Fig.5 Phylogentic tree analysis of DL7H proteins

AKI87771.1. 滇龙胆;AGX93059.1.印度蛇根草;AGX93061.1.小蔓长春花; AGX93057.1.金鸡纳树;AGX93060.1.水甘草;AGX93062.1.长春花;AGX93058.1.蟾蜍树;AGX93056.1.金银花图6 川西獐牙菜SmDL7H与其他植物DL7H同源蛋白的多序列比对AKI87771.1 Gentiana rigescens; AGX93059.1.Rauvolfia serpentine; AGX93061.1.Vinca minor; AGX93057.1.Cinchona calisaya; AGX93060.1. Tabernaemontana elegans; AGX93062.1.Catharanthus roseus; AGX93058.1.Amsonia hubrichtii; AGX93056.1. Lonicera japonicaFig.6 Multiple sequence alignment of SmDL7H protein and DL7H homologous proteins from other plants
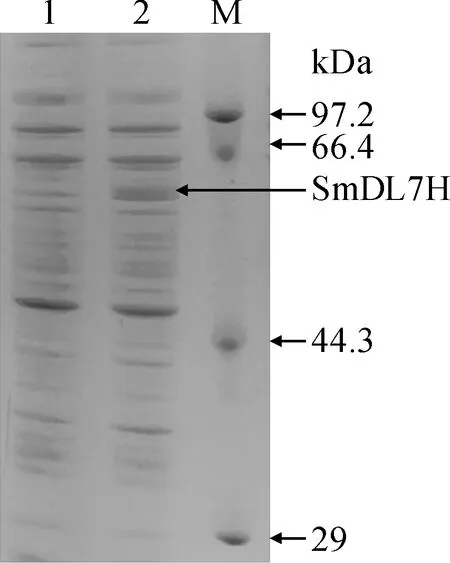
M. 蛋白质分子量标准;1. 转入pET-28a载体的Rosetta(DE3)菌株;2. 转入pET-28a-SmDL7H载体的Rosetta(DE3)菌株图7 重组蛋白的表达M. Protein marker(low); 1. Expression product of Rosetta-pET-28a;2. Expression product of Rosetta-pET-SmDL7HFig.7 Expression recombinant protein
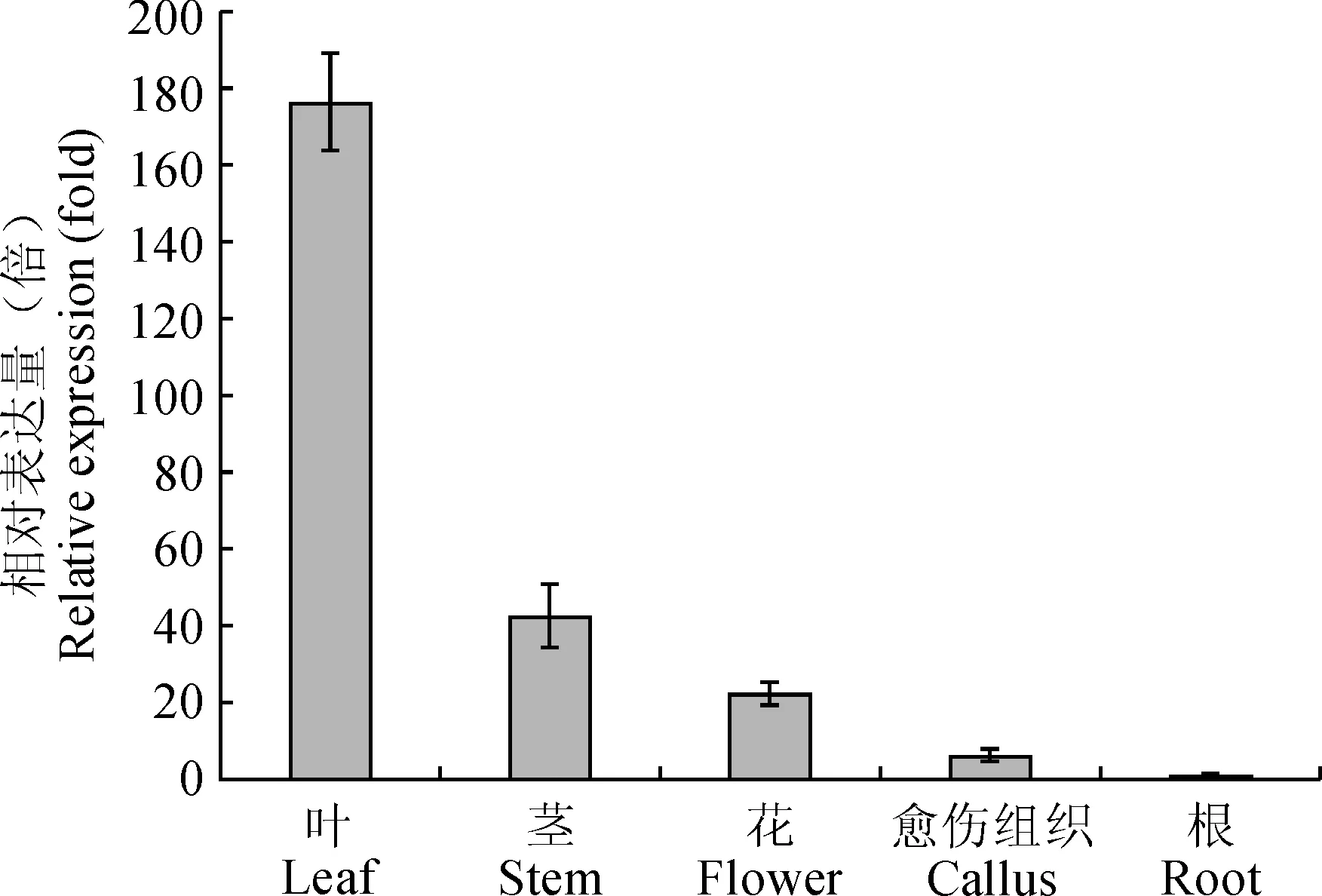
图8 川西獐牙菜SmDL7H基因在不同组织中的表达Fig.8 Different tissue expression profiles of SmDL7H gene in S. mussotii Franch
2.6 川西獐牙菜SmDL7H基因的组织表达特异性分析
荧光定量PCR结果(图8)显示,川西獐牙菜SmDL7H基因在川西獐牙菜的叶片、茎、花、根、愈伤组织中均有表达,其中在叶片中表达量最高,在根中表达量最少。推测,是由于裂环烯醚萜化合物合成途径在质体中进行,而叶片中叶绿体含量较多,故而该基因在叶片中表达量最高。
3 讨 论
“藏茵陈”是一类治疗肝胆疾病的药物的总称,川西獐牙菜是其基原植物之一,川西獐牙菜作为“藏茵陈”入药在许多典籍,例如《晶珠本草》、《四部医典》等藏医药典籍中均有记载。使用历史悠久、疗效甚佳。其有效成分主要为裂环烯醚萜类与三萜类等化合物[9-10]。川西獐牙菜生长于高寒地区,种子萌发率较低,并且无人工大规模种植。随着民族医药的兴起,对其的需求量日益增长,导致其野生资源受到严重的损害[11]。目前,对于川西獐牙菜在分子生物学方面的研究较少,对于其裂环烯醚萜生物合成途径的研究亦较少。本课题组在前期工作中完成了川西獐牙菜转录组测序,对川西獐牙菜裂环烯醚萜途径关键酶牻牛儿基焦磷酸合成酶(geranyl diphosphate synthase; GPPS)及环烯醚萜合成酶(iridoid synthase; IS)的功能进行了探索[12-13]。川西獐牙菜SmDL7H能够催化7-脱氧马钱子酸生成马钱子酸,是川西獐牙菜裂环烯醚萜生物合成途径中较下游位置的关键酶。对于DL7H在植物中的研究较少,目前仅有长春花[7]和金银花[8]中DL7H的功能研究报道。对川西獐牙菜中的7-脱氧马钱子酸羟化酶的深入研究,来提高川西獐牙菜中裂环烯醚萜类化合物的合成具有较大意义。
本研究首次在川西獐牙菜中克隆得到的7-脱氧马钱子酸羟化酶基因(SmDL7H)的全长cDNA序列,并且通过生物信息学方法进行分析,与已在NCBI上公布的植物中的DL7H的氨基酸序列进行分析比对,预测得知该基因亦属于p450超家族,并且无信号肽,且与滇龙胆、长春花[7]、金银花[8]等植物的DL7H具有较高相似性,故而推测SmDL7H与它们亦有相似的功能,能够催化7-脱氧马钱子酸生成马钱子酸。同时,对SmDL7H基因进行了原核表达工作,为后续的蛋白纯化工作、体外酶学实验、蛋白结晶解析蛋白构象等工作提供了坚实的基础。
本研究分析了川西獐牙菜不同器官中SmDL7H基因的RNA表达水平,该基因在叶、茎、花、根中均有表达,其中在叶片中表达量最高,在根中表达量最少,这与刘越等[14]研究结果一致。在赵忠娟等[15]的研究中,发现川西獐牙菜愈伤组织中有效成分种类与原植株中的一致,并且愈伤组织的生长速率远高于原植株。因此,川西獐牙菜的愈伤组织可能是一个较好的生物反应器,相较于原植物的根系能够产生较多的有效成分,同时缓解了川西獐牙菜野生资源的匮乏及相应的环境压力。在后续工作中,本课题组将继续研究川西獐牙菜的愈伤组织该途径中其他关键基因的表达谱及裂环烯醚萜类化合物积累的相关性,期望川西獐牙菜愈伤组织可作为生物反应器提高川西獐牙菜中裂环烯醚萜类化合物的含量,从而解决川西獐牙菜资源匮乏的难题。
猜你喜欢
昆明医科大学学报(2021年8期)2021-08-13 08:59:04
天然产物研究与开发(2018年11期)2018-11-30 02:06:34
天然产物研究与开发(2018年7期)2018-08-21 02:03:58
中成药(2017年4期)2017-05-17 06:09:26
中成药(2016年4期)2016-05-17 06:07:48
安徽医科大学学报(2015年9期)2015-12-16 11:09:46
动物医学进展(2015年10期)2015-12-07 05:46:21
动物医学进展(2015年10期)2015-12-07 05:46:17
现代检验医学杂志(2015年5期)2015-02-06 01:42:01
西部中医药(2014年6期)2014-03-11 16:07:37
