
人工合成甘蓝型油菜减数分裂期染色体行为观察
2018-10-15 07:54田燕飞朱莉莉许聪聪杨妍位芳田保明
生物学杂志 2018年5期
田燕飞, 朱莉莉, 许聪聪, 杨妍, 位芳, 田保明
(郑州大学 生命科学学院, 郑州 450001)
种间杂交是物种形成的重要途径之一,在植物进化过程中起着至关重要的作用。通过种间杂交,可以实现跨物种间的遗传物质的交换,产生变异或形成新物种。如今,在植物杂交育种过程中,通过种间杂交可以扩大原有的遗传基础,将具有优异农艺性状的野生植物基因资源转入栽培作物[1],是作物新品种选育的重要手段。然而,在植物杂交育种过程中,在整个基因组水平上,能否成功实现基因(组)间遗传物质的交换取决于基因组间的遗传相似性[2]。当栽培作物和野生种间具有较高的亲缘关系或遗传相似性时,二者杂交后基因组一般会产生较为频繁的遗传交换。例如,栽培甜菜(Betavulgaris)和沿海甜菜(B.maritime)[3]或栽培萝卜(Raphanussativus)和野生萝卜(R.Raphanistrum)[4]。
芸薹属植物包括3个基本种,即白菜(Brassicarapa, AA,2n=20)、甘蓝(Brassicaoleracea, CC, 2n=18)和黑芥(Brassicanigra, BB, 2n=16),都为二倍体植物。在自然界中,这3个基本种可以两两杂交形成复合种,也就是四倍体杂交种,包括甘蓝型油菜(Brasssicanapus,AACC 2n=38)、芥蓝型油菜(Brassicajuncea,AABB 2n=36)和埃塞俄比亚芥(Brassicacarinata,BBCC, 2n=34)[5]。通过基因组比较分析,研究发现,白菜和甘蓝基因组结构高度相似,两者的基因组可能来源于共同的祖先[6-9]。因此,由于在进化关系上A基因组(来源于白菜)和C基因组(来源于甘蓝)[7-8,10]亲缘关系较为接近,在细胞学水平上,A和C基因组在减数分裂过程中能够部分联会与配对[11-12]。目前,自然界存在的白菜(B.rapa)和甘蓝(B.oleracea)的基因组间仍保持着较高的同源性和共线性,而且染色体间也可以实现联会与配对,并且也存在基因间的遗传重组[13]。因此,通过人工合成甘蓝型油菜对油菜杂交育种和种质资源创新具有重要意义。
本研究中,我们通过胚挽救技术将白菜(AA)和甘蓝(CC)杂交获得了人工合成甘蓝型油菜。以栽培种甘蓝型油菜(来源于自然界中两个基本种自然杂交)为对照,我们初步分析了新合成油菜花粉母细胞减数分裂过程中染色体的行为,明确了新合成油菜中A、C染色体组在减数分裂过程中与栽培种油菜的差异,从细胞学水平上初步了解不同来源基因组间的相互作用,为通过人工合成手段选育甘蓝型油菜提供细胞遗传学证据。
1 材料和方法
1.1 材料
以甘蓝(CC)为父本,在花期与白菜(AA)人工授粉,授粉一周后取幼胚进行离体培养(胚挽救)获得杂交种单倍体油菜(AC)[13],经0.1%秋水仙素加倍处理,获得新合成甘蓝型油菜(AACC)。栽培种油菜品系JY-7由本实验室选育并提供。
1.2 方法
将油菜种子置于潮湿滤纸上,光照培养。待种子萌发,取下根尖,浸泡于0.2%的秋水仙素中预处理2 h,转入卡诺固定液固定。将固定后的根尖置于1mol/L盐酸中60℃酸解10 min,压片、碘化丙啶(PI)染色观察染色体数目。
选取生长健壮、无病虫害的新合成油菜以及栽培种油菜开花植株,于上午8:00到11:30采集花序置于卡诺固定液中,固定脱色24 h后转入70%乙醇中冰箱冷藏备用。制片时择取减数分裂时期的花蕾,剥出花药,用混合酶液(5%纤维素酶和5%果胶酶)于37℃烘箱中酶解过夜,清水低渗30 min,涂片、碘化丙啶(PI)染色、镜检。于荧光显微镜下观察染色体行为,同时统计新合成油菜减数分裂过程中花粉母细胞内出现异常情况的频率。
2 结果与分析
2.1 新型甘蓝型油菜染色体数目确定
利用根尖细胞学压片技术,对人工合成的甘蓝型油菜进行染色体数目鉴定,如图1所示,染色体形态较好,染色体数目2n=38条。

图1 新合成甘蓝型油菜染色体数目鉴定
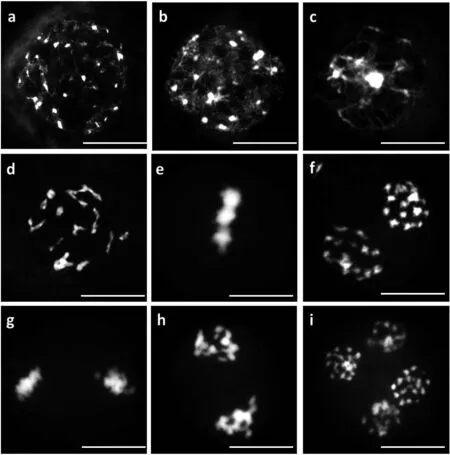
a细线期:染色体盘旋折叠,核内出现细长的染色体结构; b偶线期:开始出现同源染色体配对现象;c粗线期:染色单体间出现一个或数个交叉;d终变期:染色体变得短而粗且更浓,可清楚地观察到染色体数目;e减数第一次分裂中期:染色体逐渐移向赤道板;f减数第一次分裂后期:二价体在纺锤丝的牵引下分别移向两级;g减数第二次分裂中期:染色体整齐地排列在赤道板上;h 减数第二次分裂后期:染色体在纺锤丝的牵引下分别移向两极;i减数第二次分裂末期:移向两极的染色体开始聚集浓缩
图2栽培油菜花粉母细胞减数分裂过程
Fig 2 The meiosis process in the cultivatedBrassicanapus
2.2 栽培种油菜减数分裂过程观察
如图2所示,在栽培种油菜减数分裂过程中,其花粉母细胞减数分裂过程基本正常,一个发育正常的花粉母细胞经过 2 次核分裂和 1 次细胞质分裂,形成四分体,最后发育成有活力的雄配子体。减数第一次分裂前期,染色体呈念珠状的染色质丝(图2-a)随后开始联会(图2-b、c),然后逐渐缩短变粗,最后变成交叉端化的19个二价体,二价体构型为“V”型、“X”型和棒状(图2-d);减数第一次分裂中期,染色体排列在赤道板上(图2-e);减数第一次分裂后期,同源染色体分离分别移向两极(图2-f);此后进入第二次分裂过程。减数第二次分裂中期,2 个子细胞中的染色体都排列在各自的赤道板上(图2-g);减数第二次分裂后期,每条染色体上的2个姐妹染色单体分开,分别走向细胞一极,此时在细胞内可看到4组染色体(图2-h);减数第二次分裂末期,染色体螺旋化,新的核仁核膜重新形成(图2-i)。

a细线期:染色体盘旋折叠,核内有细长的染色体结构出现; b偶线期:同源染色体配对现象开始;c粗线期:染色单体间呈现一个或数个交叉;d终变期:染色体变粗变浓,染色体数目清晰可见,有多价体存在;e减数第一次分裂中期:染色体向赤道板聚集;f 减数第一次分裂中期:有单价体围绕在赤道板周围;g 减数第一次分裂后期:染色体分裂异常存在滞后单价体;h减数第一次分裂末期:二价体由纺锤丝的牵引移向两级; i减数第二次分裂中期:出现多个滞后单价体;j减数第二次分裂后期:每个染色单体由纺锤丝牵引移向两极;k减数第二次分裂后期:着丝点分裂为二,每个染色体具有独立着丝粒,且在纺锤丝的牵引下分别移向两级;l减数第二次分裂末期:移向两极的染色单体开始聚集
图3新合成甘蓝型油菜花粉母细胞减数分裂过程
Fig 3 The meiosis process in syntheticBrassicanapus
2.3 新合成甘蓝型油菜减数分裂过程观察
与栽培种油菜相比,新合成甘蓝型油菜在减数分裂早前期染色体形态并无明显差异(图3-a、b、c),但在终变期,除正常联会配对的二价体外,还形成大量的多价体和单价体(图3-d)。中期,由于联会和分离的不规则性,部分细胞出现游离在赤道板周围的单价体,不能正常的排列在赤道板上(图3-e、f、i)。后期,同源染色体和姐妹染色单体分离异常,落后染色体和染色体桥频繁出现(图3-g、j、k),没有正常配对的单价体在后期分裂时可能形成落后染色体而丢失,最后形成分裂不均等的4个子细胞(图3-k)。
2.4 新合成油菜减数分裂染色体异常行为统计分析
与栽培油菜相比,新合成油菜花粉母细胞减数第一次分裂中期、后期和减数第二次分裂中期、后期分裂过程中出现了异常现象,包括多价体(图3-d)、单价体(图3-f)、落后染色体和染色体片段(图3-f、i)、分裂不同步(图3-g)、不均等分裂(图3-j)等现象。统计分析,减数第一次分裂中期和减数第二次分裂中期出现异常染色体行为的频率分别为48.93%、56.52%,较高频率的染色体异常现象(图4所示),尤其是多价体的出现,对新合成甘蓝型油菜A、C基因组之间的重新排列提供了可能,并促进后代基因组的重组和优良性状的导入,扩大了种质的遗传背景。已有研究表明,多价体和单价体产生是由于同源染色体异常联会造成的;也有学者提出多价体和单价体的形成可能是由于纺锤体结构异常而导致的[14-17]。我们推测,在新合成甘蓝型油菜减数分裂过程中,出现的多价体构型有利于不同来源的染色体组间交换,促进基因组间的相互易位。
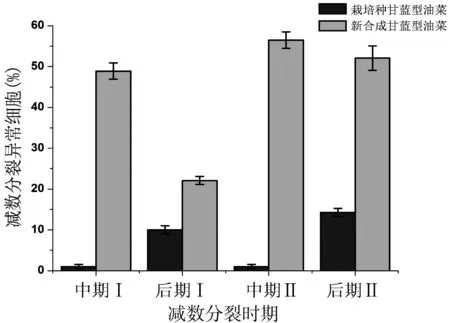
图4 新合成甘蓝型油菜减数分裂中期和后期染色体行为异常行为统计
此外,减数第一次分裂后期和减数第二次分裂后期分别有22.12%和52.17%的花粉母细胞出现染色体迁移、聚集异常,形成落后染色体或染色体片段(图3-g),表现为非正常的二极分裂(图3-i)。落后染色体或染色体片段游离于主核之外,常以微核的形式存在[18-19]。本研究中,部分减数分裂后期的子细胞中染色体数目不相同,或者数目减少,推测存在落后染色体或片段丢失的现象。此外,减数分裂的第一次分裂后期(图3-g)和第二次分裂后期(图3-j)还存在不同步分裂和不均等分离的现象,少部分染色体并未随其他染色体移动到细胞两极,其原因除了与减数第一次分裂前期异常联会有关外,还可能因纺锤丝异常和多极纺锤体引起,由于分裂过程的不协调性,导致后代细胞中染色体数目各不相同,最终导致花粉败育。
3 讨论
芸薹属各种属之间具有平行的用途和相似的形态类型,而且许多研究支持芸薹属中相似亚种的近距离进化。这都为不同种属之间优良性状的集中和种质遗传背景的扩大提供了理论支持[20-22]。A、C染色体组之间可通过染色体易位、重组、外源导入等行为进行基因交换,使杂种表现出更多的优良性状,由此创造出较多的育种新材料,从而推动油菜育种的研究[23-24]。天然杂交的甘蓝型油菜复合种表现出自交亲和而人工合成的甘蓝型油菜复合种往往表现出自交不亲和[25-26]。
自然界中,由于环境条件的影响,甘蓝型油菜减数分裂的时期在同一药室会表现出部分不同步现象。本实验中,供试材料的花粉母细胞减数分裂过程,在同一个花药的压片中均可观察到几个不同的分裂时期,存在分裂期不同步现象,从而造成四分体形成不同步,导致雄配子体形成不同步。这种不同步可能是由花粉母细胞之间胞间连丝的有无和多少、绒毡层的发育程度不同、营养不良以及外界环境影响而引起的。
现有栽培种油菜的研究结果表明,其花粉母细胞的减数分裂过程中,均为19个正常的二价体,极少的出现异常现象。而由基本种甘蓝(CC)和白菜(AA)杂交得到的新型甘蓝型油菜中,由于A、C染色体组之间难以协调,联会配对失败,造成减数分裂过程异常,染色体不同步分离,形成落后染色体,而部分配对成功的染色体,在后期分离时可能会出现染色体桥,由此形成染色体数随机不等的配子。虽然减数分裂过程中出现了较为明显的细胞学异常现象,但随着分裂过程的进行,部分细胞在减数分裂末期趋于正常。新合成的甘蓝型油菜在减数分裂过程中染色体的配对是紊乱的,天然甘蓝型油菜在形成与进化过程中必定经历了一系列选择性的变化导致了染色体行为的二倍化。继续深入了解甘蓝型油菜染色体的配对机制对培育和扩大油菜新品种,挖掘油菜种质资源具有重要意义。
猜你喜欢
数学小灵通·3-4年级(2022年10期)2022-12-31
华人时刊(2022年15期)2022-10-27
大陆桥视野(2020年8期)2020-11-26
西藏农业科技(2019年3期)2019-11-04
西藏农业科技(2019年1期)2019-07-25
西藏农业科技(2018年4期)2018-04-25
小学生学习指导(低年级)(2017年3期)2017-09-11
安徽医药(2014年9期)2014-03-20
植物营养与肥料学报(2012年3期)2012-10-26
中央社会主义学院学报(2012年4期)2012-03-20
