
渐危种黑黄檀宿存期种子萌发特性及对季节性干旱的适应
2018-10-15 03:17,,,,,
种子 2018年9期
, , , , ,
(1.西南林业大学生态与水土保持学院, 云南 昆明 650224; 2.昆明市园林科研所, 云南 昆明 650224; 3.中国林业科学院资源昆虫研究所, 云南 昆明 650224)
黑黄檀(DalbergiafuscaPierre),又名版纳黑檀、牛角木,属蝶形花科黄檀属,国家Ⅱ级重点保护植物[1]。其材质坚实,是重要的红木资源,在我国《红木》标准(GB/T 18107—2000)中属黑酸枝木类[2];黑黄檀世界范围主产区为越南、缅甸、老挝、柬埔寨等国[3];国内分布极其狭窄,主要集中于云南西双版纳和普洱地区[1],绿春、元江有零星分布[4];多集中分布在海拔900~1 400 m的干燥、贫瘠山地和干热河谷中,或散生在山地雨林或半落叶季雨林退化后形成的次生杂木林中[5]。当前,我国红木原料基本依赖进口,《濒危野生动植物种国际贸易公约》已将七种红木原材列入管制范围,红木资源生产国多已制定了种种限制出口政策,红木市场面临着严重的资源短缺危机。黑黄檀因独特的材性可替代进口红木,但因过度利用和生境丧失,其种群数量急剧下降,已成渐危种[6]。故近些年对黑黄檀的人工繁育、种群扩繁研究成为热点。
黑黄檀果实成熟于12月中下旬,种子具有宿存特征,可持续到翌年3月[7],自然分布区11月到次年4月间降水少,具有典型的“冬旱”气候特征[8],对于不具有休眠习性的黑黄檀种子而言,如何适应这种季节性干旱目前并不清楚。种子萌发是高等植物生活史中一个重要过程,也是构建植物种群的关键环节[9]。不同植物的种子成熟后具有各自独特的宿存特征、散布式样及生境适应特征,因受水分等外界环境以及生物自身特性的影响,具有不同的萌发策略[10-12],而种子萌发策略是否成功不仅影响该物种的生存和繁衍[13],在推动自然群落的演替中也扮演着重要角色[14-15]。在多变的环境中,植物种群的建成很大程度上取决于种子在多变条件下萌发与休眠的切换能力[16]。目前有关黑黄檀的基础研究较少,主要集中在木材性质、贮藏条件、种子特性等方面[17-19],现有研究无法系统阐释种子萌发特性及影响因素以及与当地特殊的干热河谷气候间的适应关系。黑黄檀种子具有宿存特性,期间伴随自然脱水过程,该过程中种子萌发率如何变化、果荚有何影响、种子忍耐渗透胁迫的能力怎样等问题目前还不清楚;另外,其自然分布区存在季节性干旱的特征,种子是如何响应这种短暂干旱气候的未见报道。无论从种质资源保护角度还是其优越的木材价值而言,探讨黑黄檀种子在不同宿存时长、果荚的有无、渗透胁迫强弱等条件下的萌发特性及原因,对黑黄檀的高效培育、种群扩繁具有理论指导意义。
1 材料与方法
1.1 种子采集地概况
黑黄檀果实的成熟期为12月中下旬,荚果成熟时不开裂,且宿存期较长,外业调查发现,少量荚果甚至到第2年幼果期时仍存于枝头。成熟荚果于2015年1月底采自澜沧江流域普洱段和澜沧段的河谷地带,海拔800~1 200 m,年平均气温17~21 ℃,年降水量1 200~1 500 mm,主要集中在6—9月,11月至次年4月为旱季,降水量占全年的10% ~ 20%(表1);土壤为红壤或砖红壤。黑黄檀为阳性树种,多分布于向阳山坡、山脊、疏林和半落叶的阔叶林中,也见与思茅松混生。在黑黄檀采集地,其主要伴生植物有香须树(Albiziaodoratissima)、黄豆树(Albiziaprocera)、火绳树(Eriolaenaspectabilis)、云南石梓(Gmelinaarborea)、西南木荷(Schimawallichii)、白茅(Imperatacylindrica)等。
1.2 种子形态性状、千粒重及含水量的测定
成熟种子的长、宽、厚形态指标采用游标卡尺随机测量100粒种子来计;千粒重和含水量的测定参照《林木种子检验规程》(GB 2772—1999)[20]的方法,随机数取100粒,用d=0.000 1 g的电子天平称量,8次重复,将均值换算成1 000粒种子的重量。含水量测定采用高温烘干法,随机选取25粒种子(称鲜重)置于已知质量样品盒中,于103 ℃烘箱中烘17 h,迅速盖好样品盒盖子,并放入干燥器里冷却至室温,再称出种子干重,4次重复,以种子失水占鲜重的百分比表示含水量。
1.3 不同宿存时间对种子萌发的影响
黑黄檀荚果成熟后具宿存特性,为探讨宿存时长对种子萌发的影响,同时排除诸如虫害、雨淋不均等因素的干扰,在自然变温条件下,将刚成熟的荚果随机分成12份统一采用透气纸袋装存,并以悬挂于室内的方式来模拟荚果在树枝上的宿存状态,分别在采后3、6、9、12个月时测定种子含水量及萌发参数。含水量测定同1.2方法;种子萌发实验参照《林木种子检验规程》(GB 2772—1999)[20],采用2层滤纸为基质,每皿25粒,4次重复,加蒸馏水到半浸没种子,于PGX-128 A多段可编程光照培养箱进行,设25 ℃温度、14 h/d的周期性光照,光照强度为3级,每24 h统计种子萌发情况。
表1 黑黄檀种子采集地(普洱和澜沧)月均温(MT)与月均降水(MP)

采集地1月2月3月4月5月6月7月8月9月10月11月12月普洱MT(℃)12.514.217.420.021.622.221.821.720.919.115.812.6MP(mm)13.418.323.851.0150.8219.5324.3309.6167.8125.473.619.6澜沧MT(℃)13.015.018.221.022.923.422.923.022.220.417.013.5MP(mm)11.816.020.144.8161.4278.6329.2293.0191.2135.679.420.7
注:数据来源于中国天气网http://www.weather.com.cn 1971—2000年统计资料。
1.4 果荚对种子萌发的影响
黑黄檀荚果成熟时不开裂且包裹严实,外业调查时发现刚成熟的果荚具有与花类似的酸香气味,宿存几个月的荚果其部分外荚上会有因微生物或昆虫作用形成的孔洞,为探明果荚是否对种子萌发存在影响以及是否含有抑制种子萌发的物质,进行如下处理:取上述室温条件下贮存3个月的荚果,随机分成4组,第1组剥除果荚仅留种子(对照,记作S,seeds only);第2组剥除果荚并剪碎与种子一起萌发置床,即(种子+果荚)组(S+P,seeds+broken pericarps);第3组对果荚戳孔但不剥除(P-H,pods with holes);第4组完整荚果(P,whole pods);每组4个重复,每个重复25粒种子(荚果以实际种子数计),放置培养皿前统一采用0.5%的甲醛消毒30 min,蒸馏水冲洗3次,实验控制条件同1.3。
1.5 种子吸水动态
将室温贮藏3个月的荚果随机分成2组,一组去荚仅留种子,一组带荚种子,于25 ℃、14 h/d光照的培养箱中进行种子吸水动态测定,每盘60粒种子(荚果以实际种子数计),3次重复。前12 h每隔2 h取样称鲜重,12 h后每隔6 h取样称重;带荚种子测定时剥除果荚,擦干种子表面,用精密天平称重;因带荚种子取样称重后不能再放回,故每次取样5粒,共计12次。用种子鲜重增加值与初始鲜重的比例即吸水率来反映吸水状况。
1.6 萌发对渗透胁迫的响应
黑黄檀种子不存在休眠习性,成熟期12月中下旬为当地的旱季,为弄清楚萌发初期水分条件如何影响种子萌发,参考齐淑艳等[21]的研究方法,做了如下处理:将室温贮藏3个月的荚果剥出种子,分别置于5%、10%、15%、20%、25%、30%等6个不同浓度的聚乙二醇(polyethylene glycol)PEG-6000溶液中浸泡24 h,根据Michel等[22]的换算方法,上述PEG溶液对应的水势分别为-0.05,-0.15,-0.32,-0.54,-0.81,-1.15 MPa,然后置于培养皿中进行萌发实验,其他萌发条件同1.3,以蒸馏水浸泡为对照,每组25粒,4次重复。
1.7 指标计算与数据处理
种子萌发特性选取萌发率和发芽势参数来表征,当胚根突破种皮并长至2 mm时,即视为萌发,萌发率采用最终萌发率即连续7 d没有种子再萌发时结束实验;发芽势采用前5 d对应的萌发率来表示。
萌发率(%)=最终萌发种子数/供试种子总数×100%;
发芽势(%)=5 d萌发种子数/供试种子总数×100%。
对PEG渗透胁迫的耐受指标参考张晓艳等[23]采用以下2个指标表示:
1) 耐渗透胁迫适宜浓度(%)=萌发率达到对照萌发率75%时对应的PEG溶液浓度。
2) 渗透胁迫半数抑制浓度(临界值)(%)=萌发率达到对照萌发率50%时对应的PEG溶液浓度。
种子形态大小、千粒重、含水量及萌发参数指标均以平均数±标准差表示。用Excel 2016软件进行原始数据统计,用SPSS 19.0软件进行单因素方差分析,用最少显著差数法(LSD)进行均值多重比较。
2 结果与分析
2.1 种子千粒重及形态性状
黑黄檀荚果偏平,舌状披针形,内含1~2枚种子,不开裂,荚果宿存期较长,种子扁肾形,长(9.51±0.52)mm,宽(5.96±0.33)mm,厚(1.06±0.10)mm。千粒重为(47.05±0.11)g,按《林木种子检验规程》中每千克是否超过5 000粒将种子分为大种子和小种子,黑黄檀属小种子。
2.2 不同宿存时间种子含水量及萌发参数变化
黑黄檀刚成熟种子含水量为(34.60±2.84)%,含水量随宿存时间延长显著下降,尤其在前3个月脱水快速,脱水率高达61.64%,宿存时长为6~12个月之间时,种子含水量随时间缓慢下降,但变化不显著,期间含水量稳定在5%~8%之间(图1)。而萌发率和发芽势则是刚成熟种子的最低,分别为55.67%和51.62%;随宿存时间延长,伴随种子脱水过程,萌发率和发芽势逐渐升高,3个月时显著增加到95.57%和86.60%;宿存时长达1年时,种子萌发率又显著下降,但仍然维持在81.85%左右,说明种子耐脱水能力非常强。
2.3 果荚对种子萌发的影响
黑黄檀种子成熟后革质的果荚依然紧闭,果荚剥除与保留2种处理存在显著差异。(种子+果荚)组萌发率和发芽势比纯种子(对照组)稍有下降,但差异不显著,说明果荚皮不含抑制种子萌发的物质;完整荚果组相比于对照,其萌发率和发芽势均显著降低,分别为48.53%和12.79%,分别比对照减少了48.20%和85.24%;而果荚戳孔组(模拟动物或微生物的作用)萌发率和发芽势可达79.17%和30.00%;比完整荚果显著提高,但与对照相比,极显著下降了15.50%和65.38%(图2),说明果荚的存在显著抑制了种子的萌发且推迟了萌发的启动时间,而经过诸如动物啃咬或微生物对果荚的分解作用(通过果荚戳孔模拟处理),增加了水分和氧气的渗入,有利于种子内生物酶的活化,促进种子萌发。
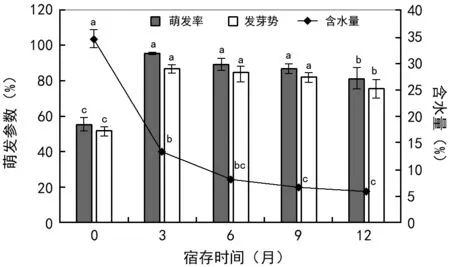
注:不同字母表示不同处理间存在显著差异。图1 宿存时长对种子含水量及萌发的影响

注:大、小写字母表示不同处理间达到极显著、显著差异;S为纯种子;S+P表示种子+果荚;P-H表示果荚戳孔;P为完整果荚。图2 果荚对黑黄檀种子萌发的影响
2.4 种子吸水动态及PEG渗透胁迫对种子萌发参数的影响
室温贮藏3个月的种子在吸水初期,吸水速率很快,最初4 h吸水重量可达自身重量的97.12%,4~8 h趋缓,接下来吸水速率又缓慢升高;24 h时种子鲜重增加到最初重量的1.58倍,此后趋于稳定(图3);说明纯种子吸水过程主要发生在24 h内;当吸水48 h时开始有种子萌出胚根;而带荚种子前8 h吸水率仅12.67%,10~24 h是快速吸水期,吸水率达110.35%,36 h后吸水率趋缓;带荚种子直到96 h吸水达到自身鲜重的1.43倍时才有个别种子萌出,发芽势仅为12.79%(图2),说明革质的果荚限制了水分的透过,延长了种子吸胀过程,推迟了初始萌发时间。
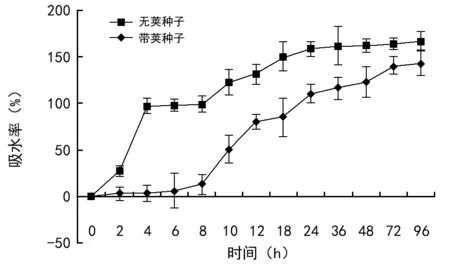
图3 黑黄檀种子吸水动态
黑黄檀种子在不同浓度PEG渗透胁迫下,萌发率和发芽势有显著差异(图4)。随PEG胁迫浓度增大,种子萌发率和发芽势均显著下降。但当PEG浓度为5%时,萌发率比对照稍有升高,但不显著;当PEG浓度达到30%时,萌发率从对照的93.47%下降到3.33%,发芽势亦从89.11%下降到0%;随着PEG胁迫程度升高,种子起始萌发时间推后,最终萌发的持续时间延长。由萌发率与PEG浓度的线性回归方程(y=-3.299x+106.892)可知,种子萌发耐渗透胁迫的适宜浓度为11.15%,PEG渗透胁迫的临界浓度为18.23%,表明,黑黄檀种子忍耐渗透胁迫的能力较强。
表2 黑黄檀种子萌发率与PEG浓度的回归分析

回归方程决定系数R2耐渗透胁迫适宜浓度(%)渗透胁迫半数抑制浓度(临界值)(%)y=-3.299x+106.8920.951**11.1518.23
注:y为种子的萌发率;x为PEG浓度;**表示在0.01水平显著。
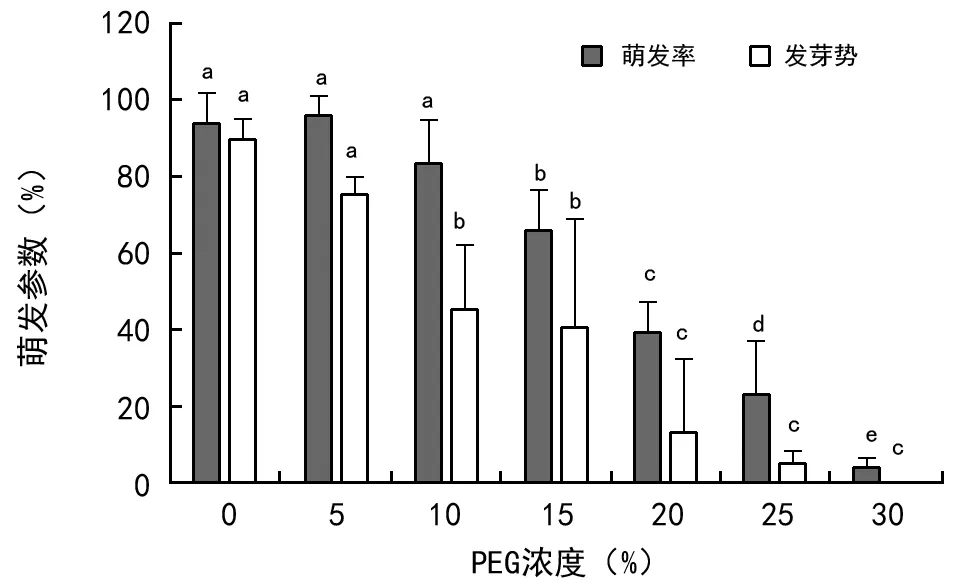
注:不同字母表示不同处理间存在显著差异。图4 PEG渗透胁迫对种子萌发的影响
3 讨论与结论
3.1 种子脱水耐性和果荚熟后不裂的生态适应意义
正常性种子在母株上成熟过程中会逐渐脱水,当脱落时含水量已较低,通常能进一步干燥到1%~5%的含水量仍具有萌发活性[24]。本研究发现,分布于澜沧江河谷地带的黑黄檀种子刚成熟时含水量为34.60%,具有熟时逐步脱水习性,属正常性种子。熟后3~6个月含水量可快速下降到13.27%至8.26%,前期快速脱水,可避免微生物滋生,减少种子损失。与此同时,伴随自然脱水过程,种子萌发率经历先升后降的过程,当自然脱水到含水量5.61%时,浸泡复水后萌发率仍可达81.85%;另外,种子萌发对PEG渗透胁迫耐受的临界浓度可高达18.23%,说明黑黄檀种子具有较强的耐脱水能力和耐渗透胁迫能力,对水分多变生境具有良好的适应意义。
黑黄檀种子成熟后及贮藏过程中荚果依然紧闭不裂,革质果皮的包裹能对种子起到很好的保护作用,这也可能是历经一年或更长时间贮藏的黑黄檀种子依然能保持较高萌发率的原因之一。然而,在自然繁殖过程中,果荚不开裂;带荚种子24 h吸水率比纯种子减少了30.26%,发芽势降低了85.24%,表明果荚对种子的紧密包裹导致通气和透水性差,并影响到萌发初期的种子吸胀过程,进而抑制酶的活化、DNA及蛋白质的合成,细胞生长受阻、胚根难以突破种皮[25];这与大多数存在透水性差的种皮或果皮限制种子萌发的研究[26-28]结果一致;此外,黑黄檀果荚戳孔后种子萌发率比完整荚果上升了30.63%,说明果荚的机械束缚也会对胚的生长具有一定的阻碍作用[29]。黑黄檀种子保持萌发活性的时间较长,有研究显示,至少贮藏18个月仍具有较高的萌发率,虽然结构致密且不开裂的果荚对种子萌发具有阻碍效应,但此期间动物或微生物的噬蚀作用可增加果荚的通气透水性,利于胚根萌出[18,30];由此可见,黑黄檀在自然更新过程中,需要在果荚遭动物或微生物破坏后所产生的2种效应——提高萌出率与种子损失风险间进行权衡。但目前有关该物种的生物学研究还不够深入,如宿存种子释放规律、种子活力最长持续时间、植冠和土壤种子库动态、果荚因生物作用所致残损的时空变化等问题仍不清楚,故其生态权衡机制有待进一步探讨。
3.2 宿存种子萌发与季节性干旱气候的适应关系
黑黄檀种子具有宿存特性,野外调查发现荚果于12月中下旬成熟,多数荚果可宿存枝头2~4个月,少量可持续到次年幼果期。与此同时,以种子采集地之一普洱为例,当地1—3月降水量约为18.5 mm/月,从5月开始进入多雨季节,直到10月底月平均降水量约216.32 mm,雨季降水量占全年的86.67%(表1),存在明显的冬春季节性干旱现象。研究发现黑黄檀种子不具有休眠习性,但刚成熟种子萌发率较低,仅55.67%(图1),宿存习性使种子与其萌发所需条件隔离,有效避免了不利季节(1—3月份)形成太多幼苗引发较高的死亡率[31];而宿存3个月后,萌发率和发芽势分别提高到90.13%~95.57%和84.33%~6.67%,此时脱落的种子萌发所形成的幼苗正好遇上水热条件均佳的季节,有利于幼苗快速生长和发育,尽快形成营养竞争力;而少量种子宿存时长达10个多月,仍保持约81.85%的萌发率,即便种子萌发,幼苗建成失败,植物在该部分所分配的资源也非常有限[32];而对应的自然条件恰恰是下一个旱季且气温较低,无论是植冠还是地面种子库中种子都趋向于“休眠”[33]并不进行吸胀萌发,故该部分种子可视为植物种群更新的冗余备份,以分摊随机风险。可见水分在时间上分布不均的生境中,植物倾向于形成较大的种子库[31],并通过宿存一段时间来调整释放时间,以便在最适合种子萌发和幼苗建成的时机释放种子,这种伴随含水量、萌发率联动的季节性差异,可认为是该物种对当地季节性干旱气候的一种适应。
与此同时,野外调查发现,植株能正常开花且大量结实,而且种子不具休眠习性,适宜的水热条件下即可萌发且萌发率高,然而这些研究结果无法解释黑黄檀林下幼苗少、分布狭窄的现象,说明导致种群濒危的原因除了人为因素外,还可能存在其他限制因素,诸如种子捕食、传播途径[34]、生境中的理化限制条件[35]、幼苗建成过程的“环境筛”效应[36]等,目前相关影响机制并不清楚。
3.3 种子萌发特性对人工繁育的启示
由于黑黄檀种子不具休眠习性,存在熟后伴随自然脱水过程萌发率增高的现象,且耐脱水和耐渗透胁迫能力均较强,建议采种时间为果实熟后3个月左右,采后即播;若无法立即播种,可于常温下带果荚置于纸袋中保存,待播时剥除果荚浸种24 h后置床萌发。
猜你喜欢
中国农机化学报(2022年11期)2022-10-27
花生学报(2022年1期)2022-08-16
林业与生态(2022年5期)2022-05-23
中草药(2022年8期)2022-04-19
江苏农业科学(2022年3期)2022-03-03
现代园艺(2020年19期)2020-10-02
种子(2019年6期)2019-07-13
种子(2019年4期)2019-05-28
农机化研究(2019年9期)2019-05-24
现代农村科技(2019年4期)2019-01-06
