
塔里木河上游灰叶胡杨克隆分株种群数量格局动态及其影响因素
2018-10-08 06:25,2*,2
塔里木大学学报 2018年3期
,2* ,2
(1 新疆生产建设兵团塔里木盆地生物资源保护利用重点实验室, 新疆 阿拉尔 843300)(2 塔里木大学生命科学学院, 新疆 阿拉尔 843300)
灰叶胡杨(PopuluspruinosaSchrenk.)隶属于杨柳科(Salicaceae)杨属(Populus),在世界上见于中亚西亚、巴尔喀什、邑尔——阿姆达里亚、中国等地,多沿河道分布。灰叶胡杨在中国仅在新疆有分布,天然分布于塔里木盆地西南部,集中分布在北纬37°~41°、东经75°30'~82°之间的叶尔羌河、喀什河、和田河一带;向东分布到拉依河湾阿拉尔、奥干河等地,即东经86°53';南抵若羌瓦石峡之西,北达达坂城白杨河出山口,伊犁河也有少量分布,其分布范围较胡杨狭窄[1]。它是新疆极端干旱区荒漠河岸林植被中重要的建群种,常以纯林或与同属的胡杨(PopuluseuphraticaOliv.)混交构成荒漠河岸林生态系统结构的主体。灰叶胡杨与胡杨相似,以种子繁殖和克隆繁殖维系种群的更新。但在河水资源日趋减少的情况下,依赖于地表水源的种子繁殖更新难以实现,克隆繁殖成为灰叶胡杨种群更新的主要方式。因此,研究灰叶胡杨克隆生长过程、克隆分株种群数量和种群空间分布格局及其与环境的关系,对揭示灰叶胡杨种群恢复的生态适应策略具有重要意义。
植物种群空间分布格局是指个体在群落中的空间分布状况,它是种群自身的特性、种间种内关系及与环境因素相互作用的结果,反映出种群的一种生态适应机制[2-4],其变动趋势是影响种群数量的主要原因[5]。种群的空间格局不仅因种而异[6-8],而且同一个种在不同发育阶段、不同生境条件、不同空间尺度下也有明显差异[9-12],因而种群空间分布格局通常反映一定环境因子对个体行为、生存和生长的影响[13-14]。生境条件变化对种群空间分布格局的影响也是显而易见的,如在塔里木河下游荒漠河岸林退化程度不同的4个典型断面,胡杨种群整体分布格局为聚集分布,并且幼苗和小树发育阶段的胡杨种群分布格局因植株数量的不断减少,逐渐由聚集分布过渡为随机分布,而中龄和老龄发育阶段的胡杨种群均为聚集分布,认为影响胡杨种群结构和分布格局变化的主要因子是地下水埋深,浅层土壤含水率对其影响不大[15]。还有研究指出,胡杨在离河不同距离大都表现为聚集分布,在离河道最远的地方有扩散趋势,呈随机分布,离河不同距离的断面均表现出随离河距离加大胡杨格局规模加大、聚集强度加强的趋势[16]。
对灰叶胡杨种群分布格局的研究表明,灰叶胡杨在不同发育阶段的空间格局均呈聚集分布,随发育进程其聚集强度逐渐减弱,被认为不同种群和不同发育阶段的空间分布格局差异与其生态生物学特性和种群间的竞争排斥有关,而且与物种的生境密切相关[17-18]。无性系植物种群分布格局反映了无性系分株在水平空间的结构,种群空间分布格局不仅会受生物和生态学特性影响,也会受光照、气候、土壤等可利用资源和种群的小生境影响[19]。灰叶胡杨在3个不同生境条件下的克隆分株种群空间分布格局均为集群分布[20],但关于灰叶胡杨克隆分株种群分布格局及动态与土壤水资源变化的关系研究尚未见报道。本研究拟揭示灰叶胡杨克隆分株种群密度的动态变化规律和克隆分株空间分布格局动态,解析克隆分株种群数量格局变化与土壤理化因子的关系,阐明洪水漫溢对灰叶胡杨河岸林克隆繁殖更新的影响,挖掘洪水漫溢在促进灰叶胡杨种群克隆繁殖更新中的应用潜力。
1 材料与方法
1.1 研究地点
研究地点位于中国新疆塔里木河上游天然灰叶胡杨林(80°50′E,40°29′N,平均海拔高度1 023 m)。研究区地势平坦,地表为细沙质,局部地段有盐结皮,每年洪水可漫及部分林地。
1.2 调查方法
样方设置在垂直塔里木河河道的方向。在距河道200 m的林缘、距河道400 m林内分别设置3个30 m×30 m的样方。采用相邻网格法,在5 m×5 m取样尺度下进行调查。在2014年和2015年的4月~10月每20天调查一次。调查内容如下:
(1)每丛克隆分株数:在30 m×30 m样方中随机选取10个克隆分株从挂牌标记,定期调查统计每丛克隆分株数,统计每丛克隆分株数的增量。
(2)克隆分株种群密度:调查统计5 m×5 m样方(定义小样方25 m2为单位面积)中克隆分株丛数,计算克隆分株种群密度(丛/单位面积)。
(3)克隆分株空间定位:调查统计每个5 m×5 m样方内每丛克隆分株的空间定位(以西南角为坐标原点,以每个样方的一个边为x轴,垂直边为y轴,确定每丛克隆分株的空间坐标值)。
(4)在每个样方内挖1 m×1 m的土壤剖面10个,自下而上每20 cm用环刀采集剖面土样。土样分为三份,一份用于烘干法测定土壤含水量,一份用于用环刀法测定土壤容重、土壤孔隙度和田间持水量;一份用于测定土壤有机质及土壤速效养分。
1.3 数据处理
种群空间分布格局采用偏离指数的t检验和Morisita指数的F检验[20]来判定。种群聚集强度采用负二项指数(K )、Cassie指标(1/K)、Green指数(GI)、Lloyd平均拥挤度(m*)、聚块性指数(PAI)比较判定。种群空间分布格局主要判定公式如下:
①偏离指数(C)及其t检验
当C>1时,判定种群为集群分布;C=1时,判定种群为随机分布;C<1时,判定种群为均匀分布;为检验C偏离Poisson 分布的显著性,应进行t 检验,其表达式为:
②Morisita指数(I )及F检验
当I >0时,种群为集群分布;当I = 0时,种群为随机分布;当I <0时,种群为均匀分布。Morisita指数用F检验,其表达式为:
种群聚集强度主要判定公式如下:
①负二项参数(K )
K值用来度量种群聚集程度,K值愈小聚集程度越高;当K值趋于无穷大时(一般是8以上),则逼近Poisson分布;K>0时,为聚集分布;K<0时,为均匀分布。
②Cassie指标(CA)
当CA>0时,为集群分布;当CA=0,为随机分布;当CA<0,为均匀分布。
③Green指数(GI)
当GI>0时,判定种群为集群分布;当GI=0时,判定种群为随机分布;当GI<0时,判定种群为均匀分布。
④Lloyd的平均拥挤度(m*)
m*表示个体在一个样方中的平均邻居数,它反映了样方内个体间的拥挤程度,其值越大聚集强度就越大,说明一个个体受其它个体的拥挤效应越大。
⑤聚块性指标(PAI)
当PAI=1时,为聚集分布;PAI>1时为随机分布;PAI<1时,为均匀分布。其值越大,集聚性越强。
2 结果分析
2.1 克隆分株种群数量的动态变化
图1a显示,2014年林缘、2014年林内及2015年林缘生境,每丛克隆分株数变化规律相似,均呈现4~7月逐渐增加,并在7月中旬达到最大值,随后每丛克隆分株数量逐渐减少。这是由于8月份开始每丛克隆分株中不断有分株死亡,另一方面每丛中不再有新的分株增加。因此,9月以后每丛克隆分株数累计值保持不变。2014年林缘、林内同期比较,林内每丛克隆分株数在整个生长季均低于林缘。
图1b显示,三个生境克隆分株种群密度变化规律相似,均表现为4~6月上旬克隆分株种群密度迅速增大,达最大值后基本保持不变。林缘、林内同期比较,克隆分株种群密度林缘显著高于林内。林缘不同年份同期比较,克隆分株种群密度2014年显著高于2015年。
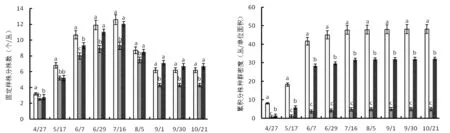
a b
2.2 克隆分株种群空间分布格局的动态变化
在5 m×5 m的取样尺度上进行格局判定,种群空间分布格局根据偏离指数的t检验和Morisita指数的F检验[21]判定。表1、2、3的判定结果显示,偏离指数(C)均大于1,且都通过了t检验,Morisita指数(I)也均大于1,都通过了F检验,Green指数(GI)和Cassie指标(CA)均大于0,聚块性指标(PAI)均大于1,即三种生境条件下,灰叶胡杨克隆分株种群的空间分布格局均呈集群分布。
Morisita指数不但可以较为准确地进行空间分布格局类型的判定,而且可以量化比较聚集强度。比较表1、表2、表3的Morisita指数可看出,Morisita指数均随着时间进程呈下降趋势,说明克隆分株的聚集强度随时间进程呈减弱的趋势。三个生境同期相比较,除4月份Morisita指数2014年林内大于2014年林缘外,其余时期的Morisita指数均为2015年林缘﹥2014年林缘﹥2014年林内。不同生境克隆分株种群聚集强度的差异可能受生境因子差异的影响。
负二项参数(K)可用于度量集群程度,客观反映格局强度,负二项参数(K)值愈小,聚集度愈大。Lloyd平均拥挤度(m*) 反映了样方内个体间的拥挤程度,其值越大聚集强度就越大,说明一个个体受其它个体的拥挤效应越大。表1、2、3显示,2014年林缘、2014年林内、2015年林缘的负二项参数(K)均是在4月值最小,表明克隆分株爆发式发生带来最大的聚集度,随时间进程负二项参数(K) 、Lloyd平均拥挤度由小增大,说明聚集度在4~6月较大;个体间的拥挤效应也越大,此后负二项参数(K) 、Lloyd平均拥挤度基本保持不变。分析认为,这与克隆分株种群密度在该时期迅速增加有密切关系。
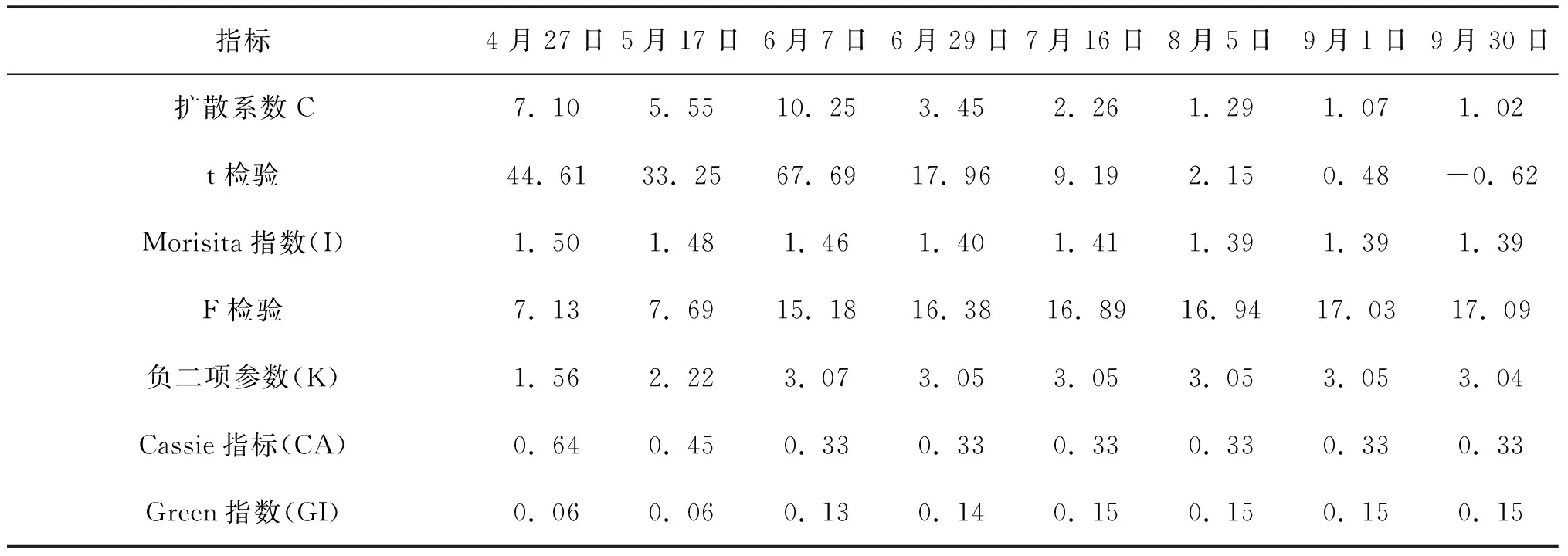
表1 2014年林缘克隆分株种群空间分布格局动态
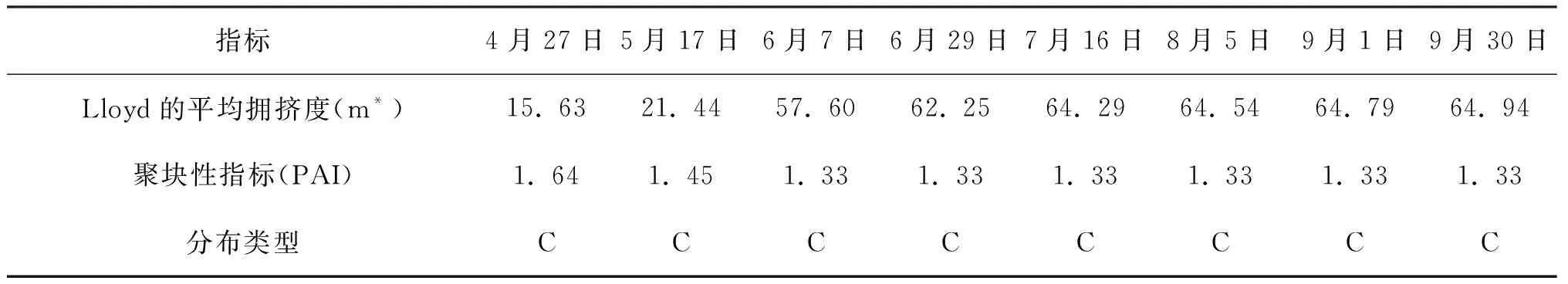
续上表
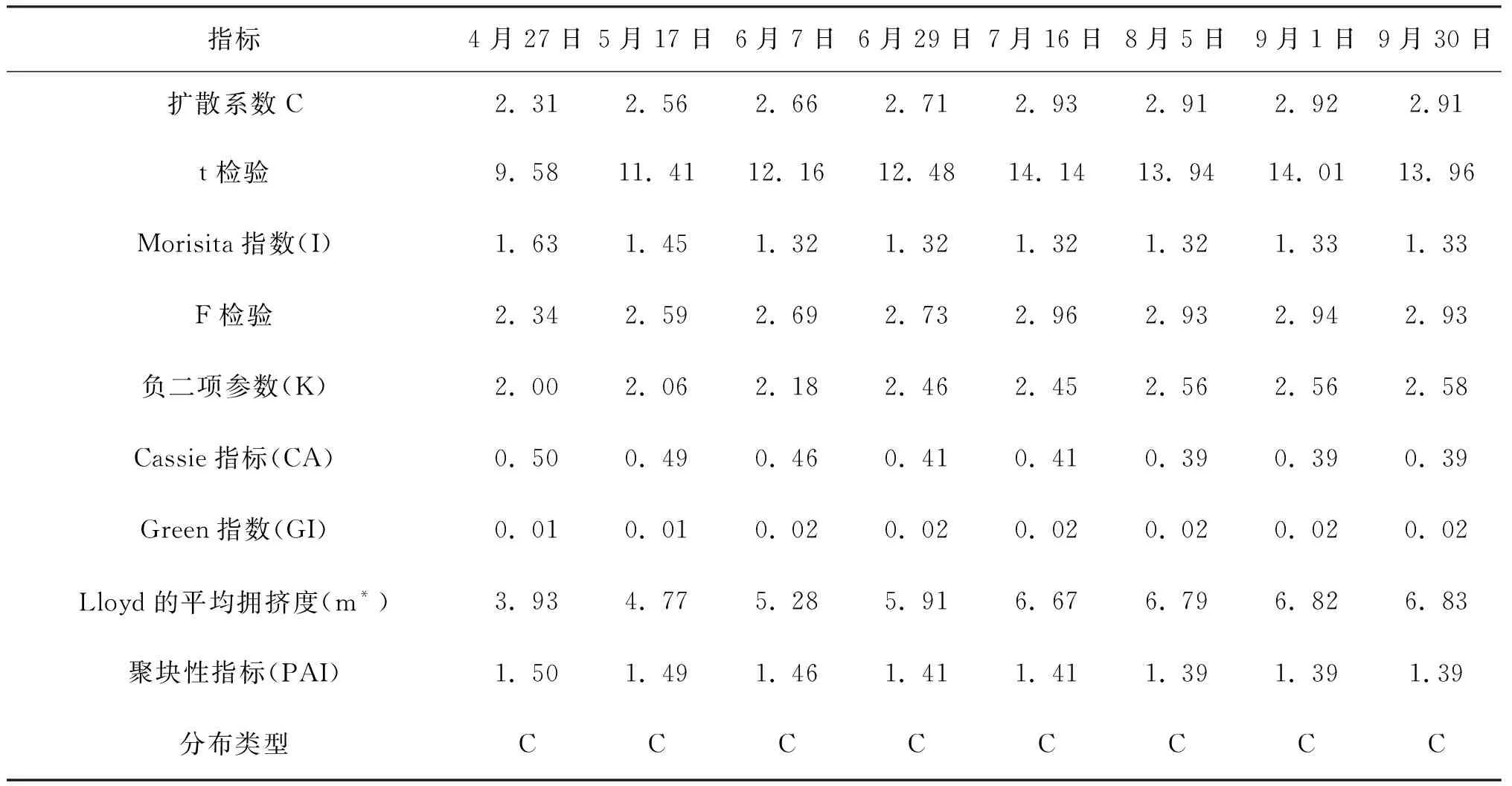
表2 2014年林内克隆分株种群空间分布格局动态
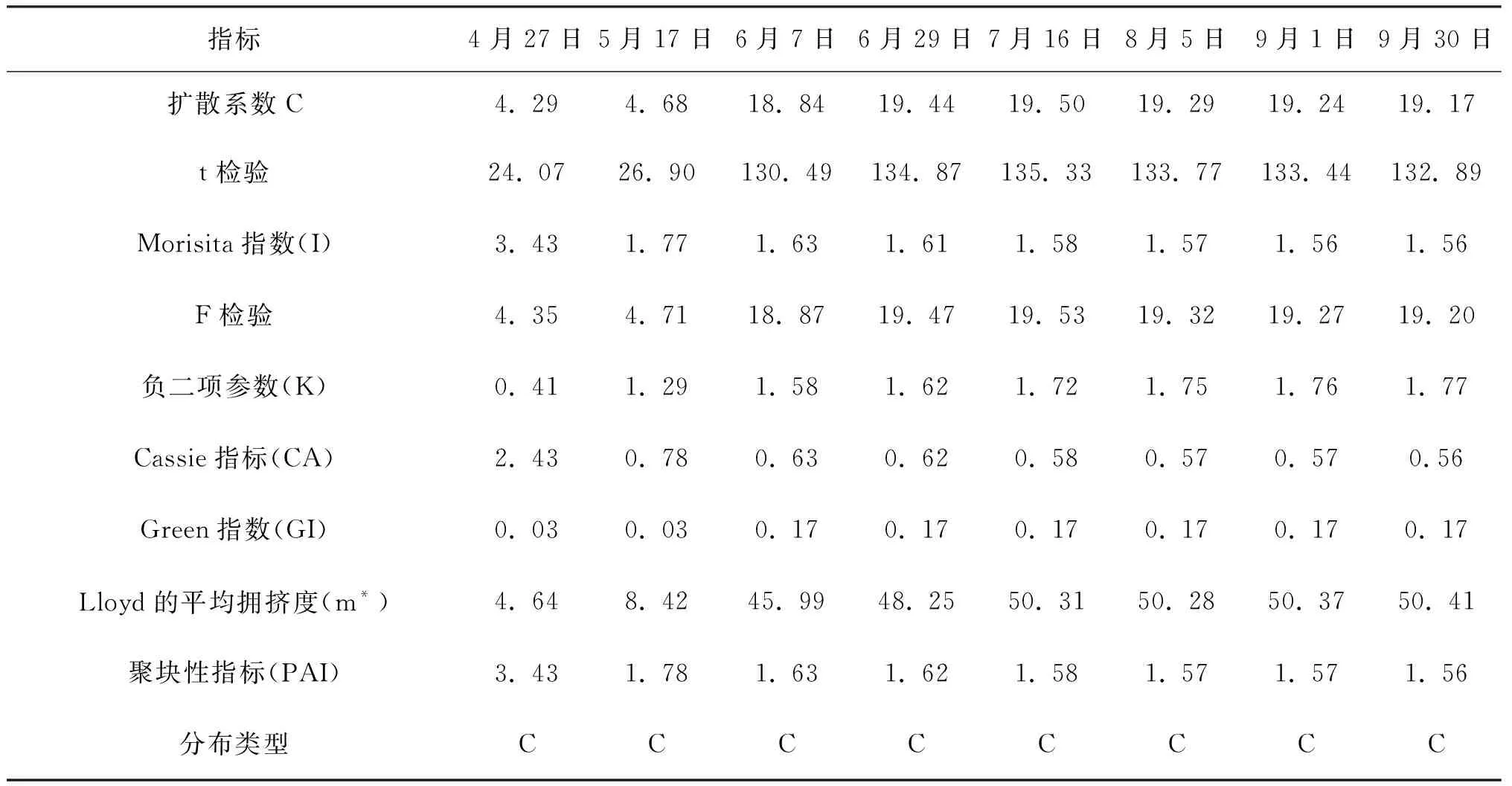
表3 2015年林缘克隆分株种群空间分布格局动态
2.3 种群密度与聚集强度的关系
从图2可以看出,克隆分株种群密度变化与Morisita指数、负二项参数(K) 、Lloyd平均拥挤度变化存在关联性,在4~6月上旬种群密度显著变化的阶段,也是Morisita指数、负二项参数(K) 、Lloyd平均拥挤度发生明显变化的阶段,说明克隆分株种群聚集强度的变化受分株种群密度变化的影响。从3个聚集度指标来看,三个生境聚集强度由大到小的排序为2015年林缘>2014年林缘>2014年林内。
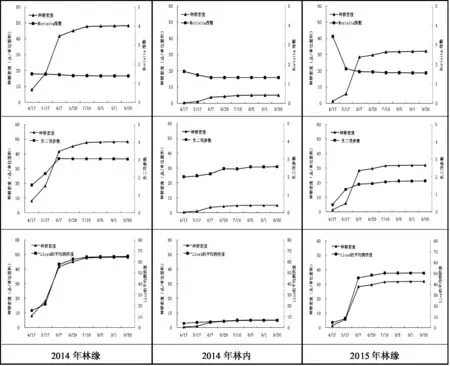
图2 种群密度与聚集度指标的关系
2.4 克隆分株种群密度与土壤理化因子的关系
表4显示,2014年林缘生境,洪水漫溢前的土壤含水量、全盐与克隆分株种群密度呈显著相关,其次是土壤田间持水量、土壤有机质和pH值;而洪水漫溢后克隆分株种群密度只受到土壤有机质含量的影响。2014年林内生境,洪水漫溢前克隆分株种群密度受土壤含水量变化的显著影响,其次是土壤有机质和全盐含量;而洪水漫溢后克隆分株种群密度也只受到土壤有机质含量的影响。2015年林缘生境,洪水漫溢前克隆分株种群密度会受到土壤含水量、土壤容重、土壤孔隙度的影响,而洪水漫溢后只受土壤碱解氮含量的影响。初步表明,当年生灰叶胡杨克隆分株种群密度的变化与生境土壤理化因子变化有关。
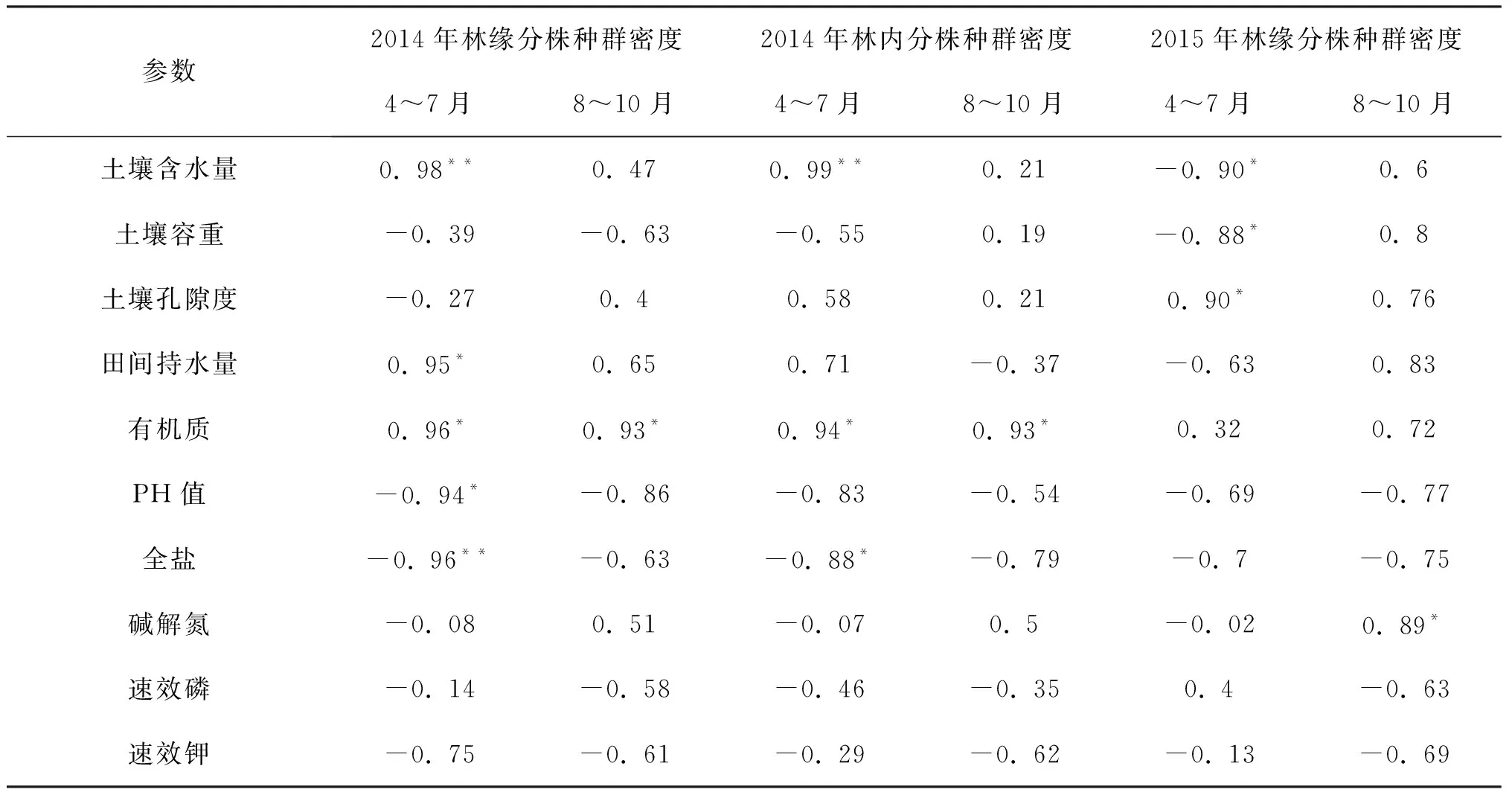
表4 克隆分株种群密度与土壤理化因子的相关性
3 结论与讨论
3.1 灰叶胡杨克隆分株种群数量的动态变化及其影响因子
灰杨克隆分株种群数量变化反映在两个方面,一方面是每丛克隆分株数的变化。灰杨不定芽集中发生的空间特征决定了克隆分株呈丛状,每丛包含克隆分株1到多个[22];另一方面是单位面积克隆分株丛数的变化,即克隆分株种群密度的变化。研究表明,2014年林缘、2014年林内及2015年林缘生境,每丛克隆分株数、克隆分株中获取密度均呈现4~6月逐渐增加,随后每丛克隆分株数量逐渐减少的变化规律。这是由于8月份开始每丛克隆分株中不断有分株死亡,另一方面每丛中不再有新的分株增加。因此,9月以后每丛克隆分株数累计值保持不变。克隆分株种群密度均呈现4~6月上旬迅速增大,随后保持不变。灰叶胡杨克隆分株种群数量的变化特征与其不定芽集中发生的空间特征和集中发生的时间特征直接相关[23],同时受到生境因子的影响,如林缘、林内同期比较,每丛克隆分株数、克隆分株种群密度在整个生长季均是林缘显著高于林内。相关分析结果表明,种群密度与土壤水分含量、田间持水量、有机质及碱解氮含量以及土壤孔隙度呈极显著/显著正相关,与土壤全盐含量和PH值以及土壤容重呈显著负相关,进一步说明当年生灰叶胡杨克隆分株种群数量变化与生境土壤理化因子变化有关。
3.2 生境因子对克隆分株种群空间分布格局的影响
干旱荒漠区植物生长对地下水有很强的依赖性,地下水埋深直接影响着与植被生长关系密切的土壤水分和养分动态,是决定荒漠区植被分布、生长、种群演替以及荒漠绿洲存亡的主导因子[24-26]。塔里木河下游地区干旱环境使得降水对植被生长的影响微乎其微,土壤浅层含水量极小,维持天然植被生长所需的水分主要来自浅层地下水[27]。对塔里木下游胡杨种群空间分布格局的研究表明,胡杨种群整体呈聚集分布,且随着地下水位的不断下降、退化程度的加剧,分布格局的聚集强度不断增强[15-16]。不同生境中各发育阶段的分布格局不同,总体趋势表现为随着退化程度的加剧,幼苗和小树的分布格局逐渐由聚集分布过渡为随机分布,而中龄和老龄则聚集度更加明显,认为塔里木河下游胡杨种群结构与分布格局主要与胡杨生长繁殖的主要限制因子地下水埋深及以根蘖的无性繁殖为主有关[16],地下水位和土壤含水量下降导致根蘖萌生的分株少[28]。
我们在5 m×5 m取样尺度下分析了不同生境、同一生境不同年份灰叶胡杨种群空间分布格局的类型及其动态变化规律,发现灰叶胡杨克隆分株种群分布格局在整个生长季呈集群分布,集聚强度由4~6月的逐渐减弱至7~10月的基本保持不变,集聚强度受克隆分株种群密度的影响,与赵正帅等人(2016)的研究结果相似[20]。分析认为,灰叶胡杨在不同生境下、以及生长季节的不同时期形成这种集群分布格局,首先是其自身克隆繁殖特性决定的。灰叶胡杨以横走侧根为克隆繁殖器官,在一条横走侧根上可以多处产生克隆分株丛、每丛幼苗数量多的特点,再加上不同母株的横走侧根水平交织在一起,在环境条件适宜条件下多个母株的横走侧根会同时或先后产生克隆分株一个部位,且大多几株拥挤在一起生长,因此呈聚集分布。这种聚集分布格局在某种程度上有利于种群形成幼苗生长的小环境,同时增强种群对其它植物侵入产生竞争等不利环境的抗性,增加其在群落中的竞争力以及种群对营养空间的占据与巩固。
其次,克隆分株种群集群分布格局的动态变化还受生境因子的影响。灰叶胡杨克隆生长过程中生境有洪水漫溢,带来了林缘和林内土壤水分含量的差异[23],以及漫溢前后其它土壤理化因子的变化和差异。相关分析显示,土壤水分含量、土壤有机质和全盐含量是洪水漫溢前影响种群密度的重要因子,洪水漫溢后土壤有机质及碱解氮含量是影响种群密度的重要因子。土壤水分含量通过影响不定芽、未出土克隆分株和出土克隆分株数量来影响克隆分株种群密度[29]。因生境因子影响克隆分株种群密度,进而影响克隆分株种群聚集强度,这就不难解释,为什么在生境资源水平低的林内生境分株种群密度最低,分株种群聚集强度最低和个体间的拥挤效应最小。尽管在林缘、林内的各个始期空间分布类型都是聚集分布,但2015年林缘的聚集强度高于2014年林缘,2014年林缘聚集强度高于2014年林内,聚集强度随密度的降低而加强,这一结果与张绘芳等[14]对不同退化区胡杨空间分布格局的研究结果相似。2014年林缘是土壤水分状况最好、种群密度在整个生长季都最高,但种群聚集强度却是最低的,说明灰叶胡杨通过调节聚集强度以减少克隆分株间的资源竞争,以确保一定克隆分株密度下的克隆生长。
猜你喜欢
金桥(2021年11期)2021-11-20
落叶果树(2021年6期)2021-02-12
北广人物(2020年47期)2020-12-09
大众文艺(2019年17期)2019-09-09
今日农业(2019年13期)2019-08-12
人民调解(2019年5期)2019-03-17
现代园艺(2018年1期)2018-03-15
创新作文(1-2年级)(2018年6期)2018-01-22
体育科技(2016年2期)2016-02-28
浙江大学学报(农业与生命科学版)(2015年4期)2015-12-15
