
湿地植物供碳功能与优化
2018-09-28 08:44赵德华吕丽萍安树青
生态学报 2018年16期
赵德华,吕丽萍,刘 哲,张 淼,安树青
南京大学生命科学学院,南京 210023
当前,污水处理厂尾水排放标准日趋严厉,湿地生态工程作为一种环境友好型的污水处理方法,可作为污水处理厂提标改造、提高尾水回用利用率或是排放标准的一个重要环节[1]。与常规污水处理技术相比,湿地生态工程技术具有投资少、运营成本低、不产生二次污染等优点,特别在低浓度污水的深度处理方面具有明显的优势。目前,尾水湿地生态工程在技术研发和应用方面均得到了迅猛地发展[2- 3]。但是,污水处理厂常采用A/O 工艺,由于A/O 工艺反应器消耗了大量的COD,特别是其中的BOD,使得湿地接受的有机物负荷相对较低,造成湿地来水C/N 严重失衡,限制反硝化作用,降低湿地生态工程的净化功能[4- 7]。
针对碳氮失衡尾水湿地氮去除效率低这一问题,国内外已发展了多种解决方案,如混合高碳氮比污水、添加低分子碳水化合物和添加植物秸秆[5,8]。但是,这些方法在实际应用过程中还存在明显的不足。湿地植物是人工湿地重要组成部分,当前,对于湿地植物的功能和作用途径却一直存在争议[9- 10]。近期的一些研究认为,对于人工湿地氮的去除而言,水生植物的直接吸收作用几乎可以忽略不计[9,11],而供碳促反硝化是湿地植物最重要的功能之一[12- 13]。总体而言,虽然研究者已意识到湿地植物供碳功能的重要性,但对其供碳动态的定量化研究,以及其影响机制与调控方法的研究非常薄弱。
本文针对尾水湿地碳源严重不足问题,以提供高效、稳定碳源、降低湿地植物分解引起二次污染风险等为目标,提出基于湿地植物解决或缓解碳氮失衡湿地的解决方案:首先,综述了湿地植物供碳促反硝化的途径及其主要的影响因素,为湿地植物供碳促反硝化功能的提升与工程化应用提供理论基础或研究方向; 然后,基于相关研究,构建冷、暖季型湿地植物生物量积累动态与供碳动态的一般性模式,提出通过湿地植物配置构建自供碳尾水湿地的方案及其供碳动态,为污水深度净化湿地的健康发展与利用提供理论依据。
1 湿地植物供碳途径
湿地植物通过光合作用生成有机物,而生成的有机物以枯叶落叶、死亡残体,以及分泌物等形式进入水体、土壤表层或被底泥覆盖,直接参与或间接影响微生物的多种生物过程,影响湿地多种元素的地球化学循环。湿地植物向湿地生态系统提供碳源是其最为重要的生态功能之一[13]。湿地植物可以通过多种途径向湿地生态系统供应碳源,从而促进反硝化作用和湿地氮去除,其供碳途径主要包括(图1):
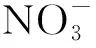
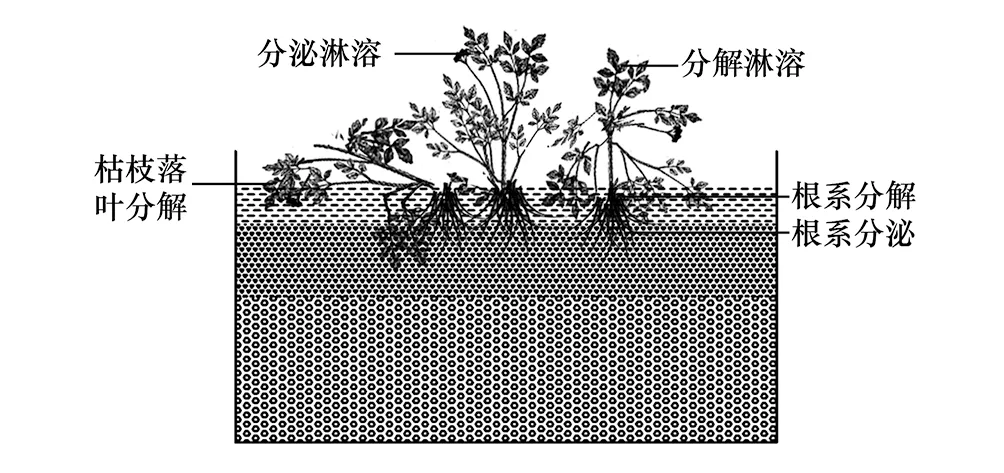
图1 生长期与凋亡期湿地植物为湿地反硝化提供有机碳途径
(2)根系自溶分解作用。与直立部分的地上部分不同,湿地植物根系生长在水中或土壤(底泥)中,根系释放的有机质可直接参与氮的反硝化作用。在生长期,除了根系分泌释放有机质外,随着新根的出现,老根的不断死亡与分解,根系脱落物和死亡老根通过自溶分解作用可以向土壤中释放大量的有机碳源,相对于植物地上部分,老根的死亡脱落可能并非是程序化事件,这一途径的碳源补充量主要是多种环境条件影响的结果[20]。而在凋亡期,死亡根系的分解可为反硝化作用直接提供碳源,湿地植物具有较高的地下生物量,这是湿地植物提供碳源的重要途径。不同湿地植物类型其地上干物质占总生物量的比值变异极大,多介于18%—72.5%之间[21- 22]。由于根系位于缺氧或厌氧环境中,其分解速度相对较慢[23],根系残体分解作用可长期为反硝化作用提供碳源。当前,有关生长期根系通过自溶分解供碳动态及其定量分析方面的研究相对较少,已有的研究大多集中在死亡根系的分解供碳[24],而对于生长期根系脱落物分解供碳的研究相对较少。
(3)地上干物质分解。生长期,湿地植物地上部分会通过程序化的死亡或对逆境的适应,产生枯枝落叶,分解(淋溶)后向湿地系统补充碳源[25]; 植物茎叶的有机分泌物在雨水淋溶下也可补充部分有机质[26]。凋亡期,随着死亡残体的分解,湿地植物可向湿地系统中补充大量的碳源,由于地上部分干物质便于收割或打捞,其供碳过程人为调控性强,而且人工湿地地上干物质的积累量较大,因此地上干物质碳源供应特征及其调控受到极大的关注[13]。在自然湿地系统中,部分的地上干物质被好氧微生物分解和以腐殖质的形式较长期留存在土壤中,部分沥出物可直接作用微生物碳源。与陆生植物相比,湿地植物植株分解物微生物可利用性更高[27]。湿地植物地上干物质的分解过程可分为两个阶段:直立分解阶段和底泥表层分解阶段,其中,直立分解阶段的有机质分解速度相对较慢[28],而且分解形成的小分子有机物进入水生态系统的比例相对较低。在人工湿地中,添加植物碳源(主要是植物地上部分)已成为提升氮去除率的一个重要的措施,得到了广泛的应用[29],而且,通过适当的前处理或工程措施可提高地上干物质用于促进反硝化去氮率[29]。
2 影响湿地植物供碳量因子
湿地植物的供碳动态是植物类型、各种环境因子和人工管理措施等多种因素综合影响的结果,影响植物光合作用和植物残体分解的各种因素均可以影响湿地植物的供碳动态。因此,这里着重分析其主要的因素,或容易人为控制的因素,进而可为其工程化应用提供依据。
2.1 根系分泌物
影响根系有机质分泌的主要因素包括:
(1)植物类型。植物类型是影响根系有机质分泌量最为重要的因素,不同植物类型由于其光合效率、生物积累、根系类型等方面存在很大的差异,根系有机质分泌物的量与质均存在很大的差异[30],总体而言,与C3植物相比,C4植物的根系有机质分配比率相对较低(与总生物量相比)[31],但是,由于C4植物光合能力与单位面积生物量显著大于C3植物,C4植物根系有机质分泌也大于C3植物。
(2)生长阶段。一般的研究认为,幼嫩的根系有机物分泌最强,随着根系的成熟与衰老,单位重量根系的有机质分泌能力出现下降的趋势,但是,由于生长期植物根系生物量的快速增长,植物根系有机质总分泌量在生长期仍然表现为不断增长的趋势[14,31]。
(3)土壤与水体理化性质。土壤有机质含量丰富的土壤其根系分泌有机质的量相对较小,而有机质缺乏的土壤其根系分泌物有增加的趋势[32]。富营养化环境可加快植物光合作用,提高根系分泌有机质[19]。而养分、水分和重金属污染等环境胁迫可显著提高植物根系分泌有机质的量[33]。
(4)温度。温度可通过影响植物的光合作用效率来影响根系有机质分泌,而且一个地区的温度与植物的物候阶段密切相关,因此,温度是影响根系有机质分泌的最为重要的因素之一,温度对根系有机质分配的影响可用Q10值表征,一些研究表明,植物根系有机质分泌的Q10接近2[19]。
2.2 地上部分分解供碳
木屑、农作物秸杆、湿地植物残体等多种形式的植物碳源被添加到人工湿地,可显著提升人工湿地氮去除率[29,34]。利用收获的地上部分作为碳源添加到人工湿地已成为一项提升湿地净化效率的重要工艺,被广泛采用,是利用湿地植物构建自供碳人工湿地相关领域的一个重要方向[13],当前,湿地植物地上部分向湿地生态系统供碳影响因素及其调控的研究主要集中在两个方面:
(1)植株地上部分分解过程。湿地植物生物质的分解过程是一个受植物类型、温度(气温和水温)、pH、水文周期、水体养分含量、盐度等多个方面因素影响的结果[35- 37]。一般研究认为,随着营养的添加,植物残体有加快分解的趋势[36- 37]; 温度和水体pH的升高同样可加速植物残体的分解[35]; 盐度的增加会降低植物残体的分解速度[38]。由于植物残体在分解释放碳源的过程中会同步释放N、P等营养元素,造成二次污染,植物残体分解过程中的COD和N、P等营养元素的同步释放是相关领域广受关注的一个方向[39- 40]。此外,植物类型与分解过程的关系方面有大量的研究报道,研究认为,不同植物类型及植株不同部位其C/N、化学组成等存在较大差异,导致其分解过程存在较大差异,一般认为,与挺水植物和陆生植物相比,沉水植物由于其木质素含量较低,表现为更快分解速度[41];与茎杆相比,叶片中木质素含量较低,其分解速度更快[42]。沉水植物、浮叶植物和挺水植物地上部分植物残体的分解过程存在显著差异,但不同的研究结果差异较大:李文朝等研究表明,浮叶植物分解最快,挺水植物次之,而沉水植物最慢[43];曹培培等和曹勋等研究表明,浮叶植物分解最快,沉水植物次之,而挺水植物最慢[44- 45]。
(2)植物碳源前处理工艺与植物碳源的添加方式。作为植物材料重要组成部分的木质素在自然条件下很难分解,且影响纤维素和半纤维素等的分解,因此在人为添加植物碳源湿地中,需要先对植物材料进行前处理,破坏木质素结构,提高植物材料的分解供碳速率和效率,已发展了较多的前处理工艺,包括植物材料的机械粉碎、高压蒸煮和辐射处理等物理方法,以及酸处理、碱处理和氧化处理等化学方法[29, 46- 48]。此外,植物碳源的添加方式会影响其供碳效率和湿地的净化效率,如果将植物材料添加至人工湿地表层好氧区域,不仅会造成植物材料的损失且抑制硝化作用,造成反硝化底物不足,降低总氮的去除,因此,一般认为,直接将植物材料添加至人工湿地的厌氧区域,直接为反硝化提供碳源是较为理想的添加方式[46,49]。但是,出于便捷性考虑,也有研究认为,可以将植物碳源溶解(分解)在水中后再添加的方式[50]。
2.3 根系自溶分解供碳
当前,生长期老根死亡脱落分解的定量化观测及其影响因素的研究少见报道。有关根系自溶分解供碳影响因子的研究大多集中在植物根冠比[21- 22]、根系积累量动态和根系有机质分解过程[24]等方面。死亡根系的分解过程也是环境因子和植物根系特征综合影响的结果。环境溶氧量是影响根系分解最为重要的因素之一,总体而言,相对于地上部分,死亡根系主要处于缺氧或厌氧环境中,其分解速度相对较慢[23],因此,尽管根系的生物量低于地上生物量,但是根系对于底泥有机碳变化的作用可能要大于地上部分[51]。而水深的增加会降低表层底泥的溶氧量,降低好氧微生物活性,从而降低表层根系的分解速度,因此,持续处于淹水状态的湿地根系其分解速度小于周期性淹水湿地; 而水深对深层根系的分解速度影响相对较小[24]。温度是影响根系分解速度的另一重要影响因子,温度对根系分解速度的影响可以用Q10来表征,根系分解的Q10在2左右[24];但是,也有研究认为根系的分解速度对温度的变化不敏感[52]。不同来源植物根系由于其C/N和化学组成的差异较大,根系的分解速度也存在较大差异[24];此外,受底泥性质影响,与粗根系相比,细根系的分解速度可能更慢[23]。
3 促反硝化潜力及其季节动态
3.1 湿地植物促反硝化潜力
受生长条件和物种类型影响,湿地植物的光合能力与有机质生产能力必然存在剧烈的时空变异[13],以常见湿地植物芦苇为例,根据上述的供碳途径可对其供碳量进行初步的估算:①地上生物量,研究表明,芦苇地上生物量可在0.413—9.89 kg/m2范围内宽幅波动,而常见的波动范围为1.0—2.0 kg/m2[53],华东地区人工湿地芦苇地上最大生物量可达6.6 kg/m2[54]; ②地下生物量,湿地植物地下与地上生物量之比(根冠比)可在0.38—4.55[21- 22],甚至更大范围内宽幅波动[55],芦苇的根冠比在自然湿地中一般大于1,而在人工湿地中一般小于1[55],本文以冠根比0.8进行估算。③生长期的枯枝落叶和死亡根系,以生长期凋落物量占最大生物积累量的19.7%进行估算[56]。④根系分泌有机质量,以茭白、美人蕉和水柳生长期平均值0.61 mg g-1d-1估算[14]。不同类型植物碳源的脱氮容量同样也存在较大差异,本文以脱氮容量为0.05估算湿地植物供碳促反硝化脱氮潜力[13, 19, 57- 58]。因此,地上最大生物量为5 kg/m2的芦苇全年脱氮潜力可达0.57 kg N m-2。这一数值高于前人对湿地植物供碳促反硝化潜力的估算,其主要原因在于前人的估算中只考虑湿地植物地上部分生物量,忽略了对地下生物量、生长期凋落物和根系有机质分泌的估算[13]。
3.2 供碳促反硝化季节动态
湿地植物供碳促反硝化能力与生物量供应间存在密切相关性,因此,单位面积生物质积累季节变化规律可表征湿地植物供碳促反硝化动态特征。湿地植物生物质积累与分解动态受多种因素的影响,不仅受湿地植物类型、区域环境条件的影响,而且与湿地植物生长的小生境密切相关,人工湿地中湿地植物的供碳动态必然受湿地植物类型的选择、管理措施、基质选择、来水特征等多种因素的影响,因此,不同的人工湿地,湿地植物供碳动态必然存在极大的时空异质性。但是,根据已有的大量相关研究,仍然可归纳总结来一个相对普适性的规律或模式。以华东地区为例,该地区常见湿地植物可分为2类:一是以芦苇、菖蒲、鸢尾、美人蕉等为代表的暖季型人工湿地植物,二是以黑麦草、水芹、菹草等为代表的冷季型人工湿地植物。暖、冷季型湿地植物潜在可利用有机质量(地上和地下的当年积累有机质量+往年未分解有机质量+根系分泌物量)存在着季节性的波动(图2),暖季型湿地植物可利用有机质在夏季和秋季较高,而在冬季和春季较低; 而冷季型湿地植物可利用有机质量表现出相反的季节变化规律[35- 36, 39, 40, 53- 55, 59- 60]。潜在可利用有机质量与实际有机质供应量(以可溶性的有机酸、糖和蛋白质等形式存在)季节间表现出显著差异[14- 15, 24- 25, 36, 43, 61- 62](图3)。暖季型湿地植物实际有机质供应量在10月至12月间处于高位,6月到8月处于低位; 而冷季型湿地植物有机质供应量在4月至9月间位于年内高位,而其他月份相对较低。此外,暖、冷季型湿地植物提供的碳源中有近10%来自根系分泌的有机质。表明,在近自然不收割条件下,暖、冷季型湿地植物无论是潜在可利用有机质量,还是实际有机质供应量均存在显著的季节性互补性规律。
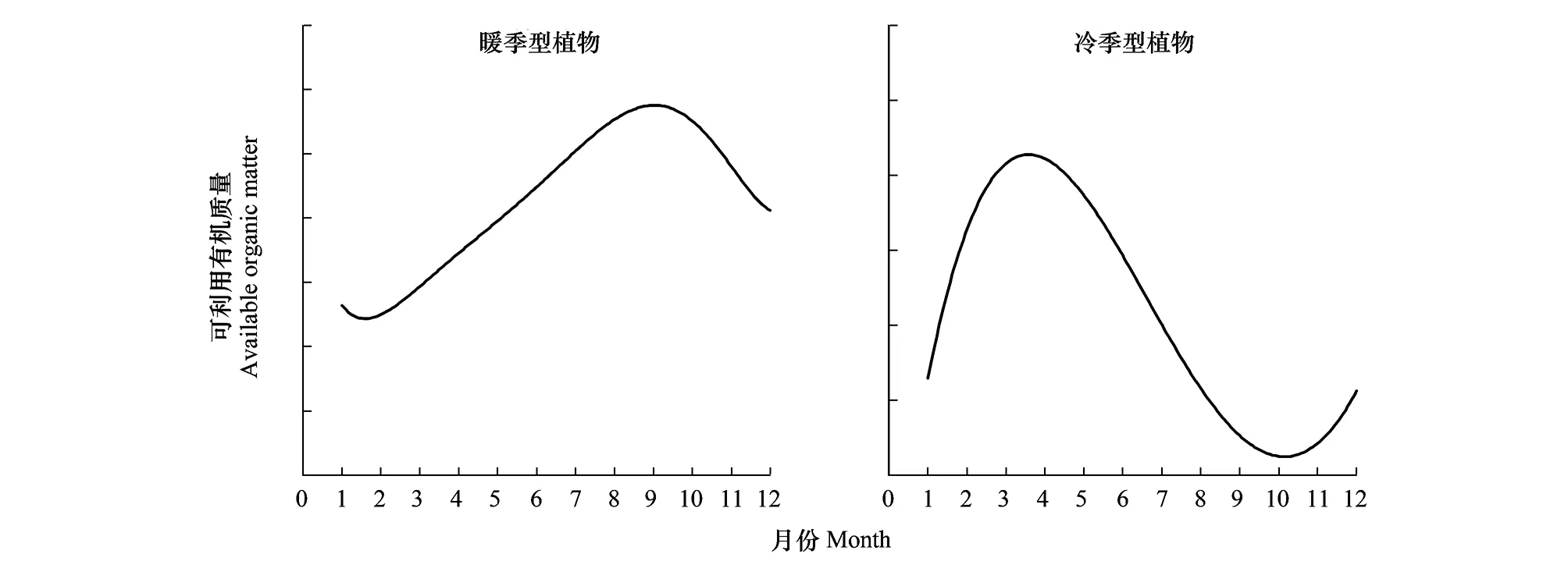
图2 华东地区典型暖季型和冷季型湿地植物可利用有机质季节变化动态
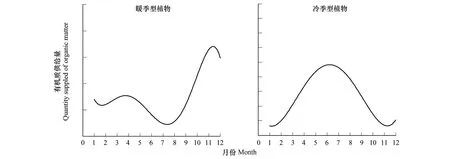
图3 华东地区典型暖季型和冷季型湿地植物有机质供给量季节变化动态
3.3 湿地植物供碳促反硝化功能调控
影响湿地植物供碳功能的因子很多,对于人工湿地而言,植物种类的选择是调控湿地植物供碳功能最为常用的措施[13, 55]。但是,湿地植物体内含有大量的矿质养分,管理不当容易引起二次污染[55]。例如,芦苇植株含氮量可在0.35%—2.69%范围内宽幅波动,平均值为1.0%—1.5%[63],也即植株重返生态系统氮量占其供碳促反硝化脱氮量的20%—30%。因此,按湿地系统反硝化需求定量供给湿地植物生物量,避免湿地植物产生二次污染就尤为重要。如前所述,湿地植物的潜在可利用有机质量和实际有机质供应量均存在显著的季节性波动,运用不当可能会造成一些时期二次污染的发生。如图2—3所示,暖季型和冷季型湿地植物间存在显著的季节互补作用。因此,在同一湿地生态系统将暖、冷季型湿地植物进行合理的配置,可为生态系统提供稳定的碳源,提升供碳促反硝化能力,降低植物残体集中分解引发的二次污染风险[64]。辅以收割和前处理后回用等措施将进一步调控湿地植物的供碳能力[8, 13]。此外,在湿地生态系统中暖、冷季型湿地植物配置,两个方面的问题需引起关注:一方面,由于暖、冷季型湿地植物物候方面的差异,可提高生态系统单位年生产量,从而进一步提升供碳促反硝化能力[65]; 另一方面,暖、冷季型湿地植物物候期存在部分重叠,而且两种类型植物可能产生复杂的相互影响[66]。
4 研究展望
(1)湿地植物供碳促反硝化定量化研究及其调控有待加强。当前,有关植物残体分解过程的研究较多,这些研究可为凋亡期的供碳规律提供参考; 而对于生长期的湿地植物供碳功能研究相对较少。如前所述,湿地反硝化作用与温度密切相关,因此碳氮失衡湿地的碳缺乏问题在春、夏温度较高时表现尤为突出; 而芦苇、菖蒲等夏季型植物在人工湿地中应用最为广泛,春、夏季为这类植物的主要生长期,其实际供碳能力却相对较弱,因此,湿地植物在生长期的供碳促反硝化作用尤为重要。当前,对于湿地植物生长期的供碳研究,如定量化研究生长期湿地植物通过枯枝落叶或老根系脱落分解供碳的定量化研究方面少见报道。此外,根系分泌有机质是湿地植物供碳的重要途径,而当前对根系有机质的分泌的研究主要侧重于其成分分析与生态学作用的研究,对其作为湿地碳源供应功能的研究也相对较少。而湿地植物的供碳过程和能力是由光合作用、碳水化合物分配和有机质分解等过程综合作用的结果,表现出很强的可调控性,有关湿地植物供碳功能的定量化研究,特别是生长期供碳功能的调控及其工程化应用方面有待于加强。
(2)湿地植物供碳定量化研究方法上存在较大的不确定性。当前,对于根系有机质分泌动态的研究多采用室内水培法,或者是通过分析根系在超纯水中短期分泌量来进行定量分析,研究的是水体中有机质严重缺乏条件下的分泌动态,这一研究结果与自然条件下湿地植物根系有机质的分泌动态必然差异较大; 有关湿地植物残体分解动态的研究多采用将烘干样品装袋后放置水中或埋于土中进行观测的方法,这与自然条件下的分解过程也必然差异极大[67]; 而有关湿地植物根系分解过程的研究,出于操作性考虑,样品多为生长期采集的活体根系,研究的周期也与自然条件下的分解周期差异较大[68- 69]。此外,湿地植物可利用有机质量和有机质供应量不仅与植物的光合过程和分解过程有关,还会受水流、水位等多因素影响。因此,有必要发展新的研究方法,分析近自然条件下的湿地植物的供碳动态,为定量化分析湿地植物供碳动态,构建自供碳人工湿地提供依据。
(3)稳定高效供碳促反硝化人工湿地构建技术有待于进一步优化。湿地植物供碳促反硝化能力与动态是多种因素综合影响下的结果,当前,与供碳动态密切相关的湿地植物光合作用、根系有机质分泌和有机质分解的影响因子的研究报道很多,如温度、光照、水分、重金属、物种、管理措施等,但是,人工湿地的运行过程中,有些因素的人为调控能力有限,其影响机制的实用性不足; 而有些因素,如物种、水分状况、收割等,人为调控相对便捷,如何调控湿地植物的供碳过程是相关领域的相对薄弱之处。本文研究表明,不同物种的供碳动态存在较大差异且存在季节性的互补特征,通过物候组合可实现稳定的碳源供应和构建自供碳人工湿地,从而解决一些湿地系统碳缺乏引起的氮去除问题。当前,虽然有关人工湿地中湿地植物的配置研究很多,但大多基于景观、群落稳定性等角度考虑,存在目标模糊、效益提升有限等问题。有必要加强湿地植物供碳与植物配置关系,以及不同物候类型湿地植物间的互作关系的研究。此外,进一步整合物种配置和其他调控因子,构建更加高效、稳定供碳促反硝化人工湿地有待于进一步研究。
猜你喜欢
环境工程技术学报(2022年3期)2022-06-05
中国土壤与肥料(2021年5期)2021-12-02
土壤学报(2021年4期)2021-09-10
昆钢科技(2021年6期)2021-03-09
生态学报(2019年8期)2019-05-31
环境保护与循环经济(2017年7期)2018-01-22
环境保护与循环经济(2017年10期)2017-03-16
电源技术(2016年9期)2016-02-27
中国环境科学(2016年3期)2016-02-08
西藏大学学报(自然科学版)(2015年1期)2015-12-25
