
中亚热带不同母质和森林类型土壤生态酶化学计量特征
2018-09-28 08:45张星星杨柳明李一清林燕语郑宪志楚海燕杨玉盛
生态学报 2018年16期
张星星,杨柳明,*,陈 忠,李一清,3,林燕语,郑宪志,楚海燕,杨玉盛
1 福建师范大学地理科学学院,福州 350007 2 湿润亚热带山地生态国家重点实验室培育基地,福州 350007 3 夏威夷大学希罗分校农林和自然资源管理学院,希罗 96720
土壤微生物和植物分泌的胞外酶是土壤有机质分解的关键媒介,是细胞代谢的产物,其活性与产量和碳底物及氮、磷养分有效性密切相关[1- 2]。通常认为,微生物用于获取碳、氮、磷的主要胞外酶比值(βG∶NAG∶AP),即生态酶化学计量比,能够反映微生物群落的新陈代谢及养分需求与环境中养分有效性之间的生物地球化学平衡模式,这可以用来衡量土壤微生物能量和养分资源限制状况[3- 4]。
现有研究表明,多种生物因子和非生物因子都能够影响土壤生态酶化学计量比。气候通过调节土壤微生物的生长速率和养分有效性从而改变土壤生态酶化学计量比[5- 6],并且土壤生态酶化学计量比亦受土壤养分状态、pH[7]、土壤C∶N∶P以及土壤质地[8]等非生物因子的影响;例如Peng等在中国北方温带草原研究发现土壤养分有效性主要影响了土壤生态酶化学计量[2]。此外,其亦可能受生物因子的影响,如地上生物量[9]、根生物量[10]以及微生物生物量化学计量比[11]的影响,Allison等研究亦表明微生物量C∶N∶P比值不同的微生物群落能够通过分泌特定的胞外酶从复杂的基质中获取限制性养分,从而改变土壤生态酶化学计量比[12]。虽然,生物和非生物因子对土壤生态酶化学计量比的影响已经受到关注,但是各种因子对土壤生态酶化学计量比影响的贡献比例仍不明确。
土壤母质和森林类型都是影响森林土壤养分有效性的重要因素。土壤母质决定土壤养分的初始供应,并且影响土壤对养分的保持能力[13],如赵汝东等[14]研究发现花岗岩发育的马尾松人工林土壤养分、生化强度、磷酸酶活性等较砂岩发育的土壤高。而森林类型则通过不同凋落物输入改变土壤养分的有效性和循环。如杨玉盛[15]研究表明天然常绿阔叶林转变为杉木人工林,土壤C、N、P等养分流失及生产力下降,这主要是由于天然林转变成人工林后,凋落物数量显著降低的结果。关于土壤母质和森林类型对土壤养分的影响已有大量报道,然而这两种因素将如何通过调控土壤生物和非生物因子,从而对土壤生态酶化学计量特征产生影响仍需要深入研究。因此,本研究分别选取了砂岩和花岗岩土壤的米槠林和杉木林为研究对象,测定了土壤的物理化学性质、微生物生物量碳、氮和磷以及酶活性,研究母岩和森林类型对土壤酶活性及生态酶化学计量特征的影响,有助于完善土壤生态酶化学计量比的理论研究体系,为认识区域土壤微生物能量和养分资源限制状况提供新的视角。
1 材料与方法
1.1 研究区域概况
试验区位于格氏栲自然保护区(26°11′N,117°28′E)和金丝湾森林公园陈大林业采育场(26°19′N,117°36′E),两者在直线距离上不超过30 km,在行政区划上二者都属于福建省三明市。三明市区域内,平均海波300 m,平均坡度25°—35°,其属中亚热带季风气候,具有冬冷夏热、雨热同季的特点,多年平均气温20.1℃,年降水量1670 mm,年均蒸发量1585 mm,相对湿度81%,其中格氏栲自然保护区土壤以砂岩发育的红壤为主,金丝湾公园陈大林业采育场土壤以花岗岩发育的红壤为主。
本研究分别选取了格氏栲自然保护区和金丝湾森林公园陈大林业采育场的米槠林和杉木林。其中格氏栲自然保护区的米槠林约200年没有受到人为的干扰,建群树种为米槠,而米槠(Castanopsiscarlesii),格氏栲(Castanopsiskawakamii),杜英(Elaeocarpusdecipiens),木荷(Schimasuperba)等为主要树种。杉木林是经过米槠林皆伐、火烧等形成的人工纯林,而杜茎山(Maesajaponica),毛冬青(Ilexpubescens),华山姜(Alpiniachinensis)等为主要林下植被。金丝湾森林公园陈大林业采育场的米槠林约50年没有受到人为干扰,建群树种为米槠,其中米槠(Castanopsiscarlesii)、闽粤栲(Castanopsisfissa)、黄丹木姜子(Litseaelongata)、新木姜子(Neolitseaaurata)、木荚红豆(Ormosiaxylocarpa)等为主要树种。杉木林是经过米槠林皆伐、火烧等形成的人工纯林,狗骨柴(Tricalysiadubia)、毛冬青(Ilexpubescens)、芒萁(Dicranopterisdichotoma)等为主要林下植被。
1.2 研究方法
1.2.1 土壤采集
土壤样品于2016年3月分别在三明格氏栲自然保护区和金丝湾公园陈大林业采育场的米槠林和杉木林4个样地中采集(每个样地有3个20 m×20 m的样方)。在每个样方中用土钻按样方“S”型随机取15—20个点,并在0—10 cm土层取样。每个样方的土壤混合均匀后放入便携式冰盒中并立即带回实验室。去除土壤中可见的根及石砾等,分为两部分,一部分储存在4℃冰箱中,用于土壤酶活性的检测,另外一部分土样过2 mm的筛后再分为2部分,一部分储存在4℃冰箱中,用于土壤微生物量的检测,其余土样风干后,用于其他基本理化性质的分析。
1.2.2 土壤理化性质的测定
土壤有机碳和全氮采用碳氮元素分析仪(Elementar Vario EL Ⅲ)测定;土壤总磷采用HClO4-H2SO4消煮-钼锑抗比色法提取后,用连续流动分析仪(Skalar san++,Skalar,荷兰)测定;土壤质地采用激光粒度仪(MasterSizer 2000,Malvern,Englang)测定。
1.2.3 土壤微生物量的测定
土壤微生物量采用氯仿熏蒸-浸提法[16- 17],称取两份5 g过2 mm筛的鲜土,分别作为熏蒸和未熏蒸处理。微生物量碳、氮(MBC、MBN)用0.5 mol/L K2SO4溶液浸提,其中4∶1的水土比,震荡离心后,其上清液过0.45 μm滤膜抽滤。浸提液分别用总有机碳分析仪(TOC-VCPH/CPN,Shimadzu,日本)测定微生物量碳,使用连续流动分析仪(San++, Skalar,荷兰)测定微生物量氮。土壤微生物量磷(MBP)用0.5 mol/L NaHCO3溶液浸提,其中4∶1的水土比,震荡离心后,其上清液过滤纸后,用连续流动分析仪(San++, Skalar,荷兰)测定微生物量磷。微生物量碳、氮、磷的计算公式分别如下:
MBC=△EC/kC;MBN=△EN/kN;MBP=△EP/kP
式中,△EC为熏蒸和未熏蒸的土壤有机碳含量的差值,kC为转换系数0.38;△EN为熏蒸和未熏蒸土壤有机氮含量的差值,kN为转换系数0.45;△EP为熏蒸和未熏蒸为土壤磷含量的差值,kP为转换系数0.4。
1.2.4 土壤生态酶活性测定
参照Saiya-Cork等[18]研究方法测定3种水解酶的活性(AP、βG和NAG),用伞形酮(MUB)做标示底物,称取1 g土壤放于含有125 mL 50 mmol/L、pH值为5.0的醋酸缓冲液的广口瓶中,用磁力搅拌器搅拌5分钟使其均质化。搅拌后将悬浮液用移液器取200 μL到96孔微孔板。微孔板在黑暗中20℃孵育4 h后,向每个微孔板中加入10 μL、1 mol/L NaOH溶液使其反应停止,用多功能酶标仪(Synergy H4,美国)测定水解酶的荧光度。各种酶缩写及所用底物见下表1。
1.3 数据统计
因为亮氨酸氨基肽酶的含量(LAP)通常较低,本文根据Turner等[19]的研究方法,将分解几丁质的β-N-乙酰氨基葡萄糖苷酶(β-N-acetylglucosaminidase NAG)的氮相关酶作为氮循环的特征酶进行计算。C∶N酶活性的比值,通过βG:NAG进行计算,C∶P酶活性的比值,通过βG:AP进行计算,N∶P酶活性的比值,通过NAG:AP进行计算。用Excel 2013软件对数据进行处理后,用SPSS 19.0和Canoco Software 5.0软件对数据进行分析,主要由Origin 9.0进行绘图。不同母岩发育的土壤生物及非生物因子属性的差异使用单因素方差分析(One-way ANOVA),米槠林和杉木林土壤酶活性和生态酶化学计量比的差异采用独立样本T检验,母质、森林类型及母质和森林类型的交互作用对酶活性及生态酶化学计量比的影响采用双因素方差分析(two-way ANOVA),影响酶活性的环境因素均采用Pearson相关分析和冗余分析(RDA)来分析。
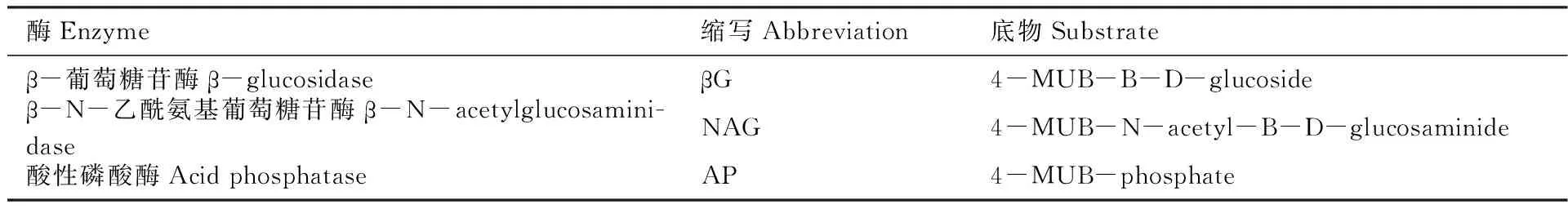
表1 土壤酶活性的种类、缩写和所用底物
2 结果与分析
2.1 不同森林类型土壤非生物因子和生物因子属性
土壤母质和森林类型均影响土壤非生物及生物因子(表2)。总体上,花岗岩发育的土壤pH、砂粒均高于砂岩发育的土壤,与此相反,TP、TN、粉粒和粘粒低于砂岩发育的土壤。其中,砂岩发育的土壤米槠林SOC、TN、TP、MBC、MBN和MBP的含量均高于杉木林,花岗岩发育的土壤则表现出相反的趋势,即米槠林SOC、TN、TP、MBC、MBN和MBP的含量均低于杉木林,结果表明土壤pH、TP及质地组成可能主要取决于土壤母质,而SOC、TN和其它生物因子主要是土壤母质和林分类型共同作用的结果。
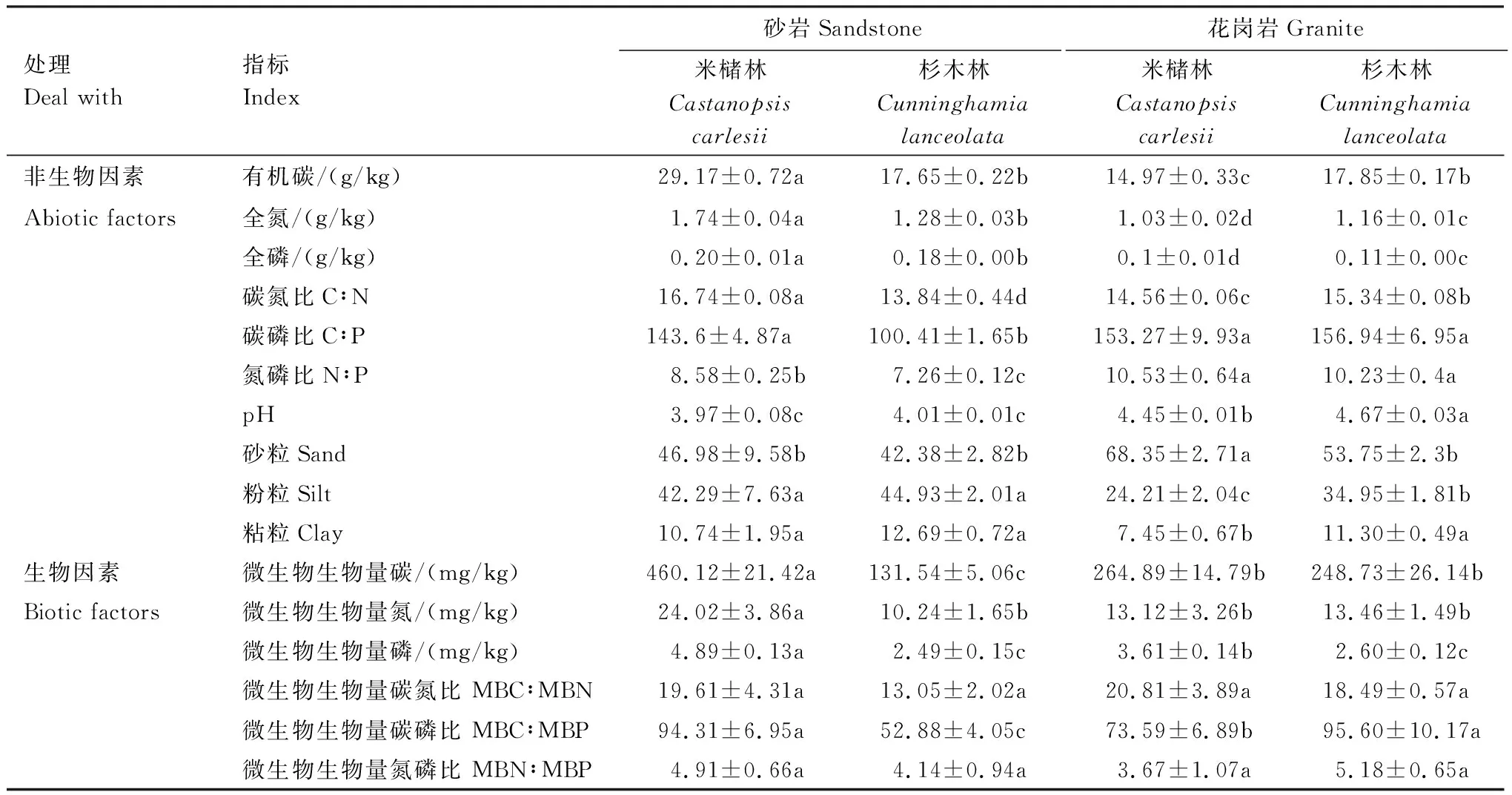
表2 不同森林类型土壤非生物因子和生物因子的含量
表中数值为平均值±标准差,同行不同小写字母表示相同指标不同林分间的差异显著(P< 0.05)
2.2 土壤母质和森林类型对土壤酶活性的影响
土壤母质和森林类型显著影响土壤AP、βG和NAG 3种土壤酶活性(图1),并且母质和森林类型存在交互作用(表3)。AP、βG和NAG 3种酶活性的变化范围分别为77.04—257.52、12.73—59.42、33.52—86.30 nmol g-1h-1,其中砂岩发育的土壤,米槠林βG和NAG活性显著低于杉木林,而AP的活性则高于杉木林,花岗岩发育的土壤βG、NAG和AP活性显著高于杉木林。此外,砂岩发育的土壤AP活性显著低于花岗岩发育的土壤,说明相对于砂岩发育的土壤,花岗岩发育的土壤中微生物受磷养分限制的强度更大,而土壤母质对βG和NAG活性影响则因林分类型的不同而不同。
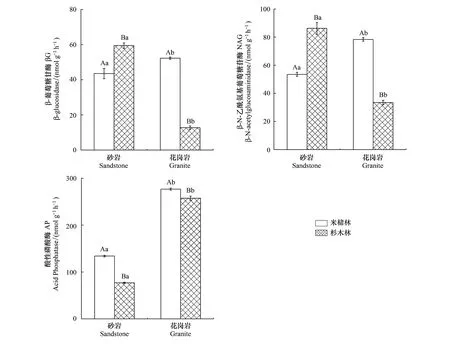
图1 不同森林类型土壤酶活性的含量
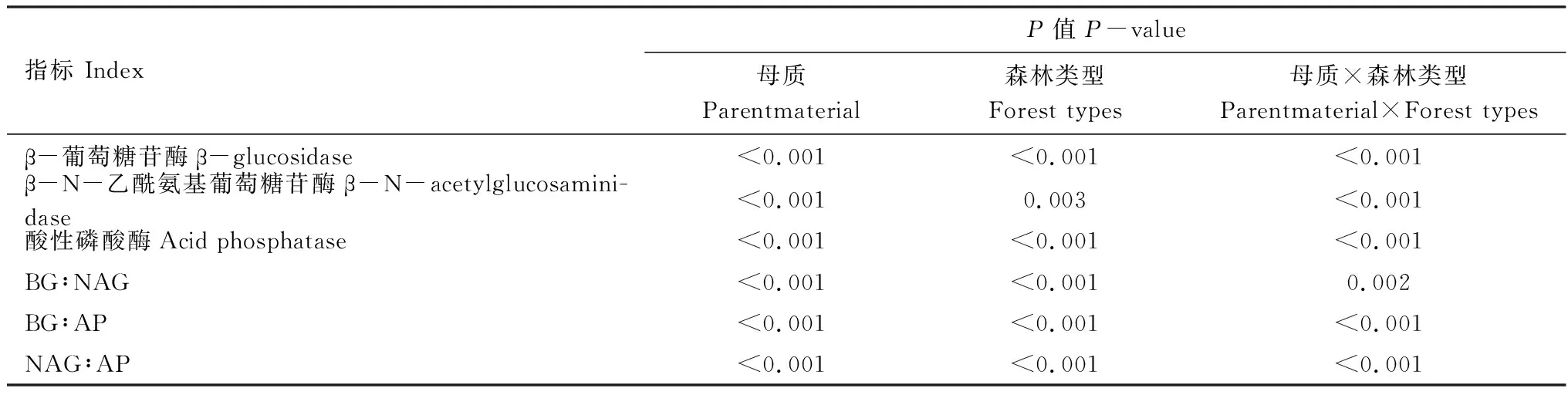
表3 母质、森林类型及其交互作用对土壤酶活性和生态酶的化学计量比影响的双因素方差分析
2.3 土壤母质和森林类型对土壤生态酶化学计量比的影响
土壤母质和森林类型亦显著影响土壤生态酶化学计量比(图2),并且母质和森林类型存在交互作用(表3)。βG:NAG、βG:AP和NAG:AP的变化范围分别为0.38—0.81、0.05—0.77、0.13—1.12,其中砂岩发育的土壤,米槠林βG:AP和NAG:AP显著低于杉木林,而βG:NAG则显著高于杉木林。花岗岩发育的土壤,米槠林βG:NAG、βG:AP 和NAG:AP都显著高于杉木林,并且砂岩发育的土壤βG:AP和NAG:AP都显著高于花岗岩发育的土壤。
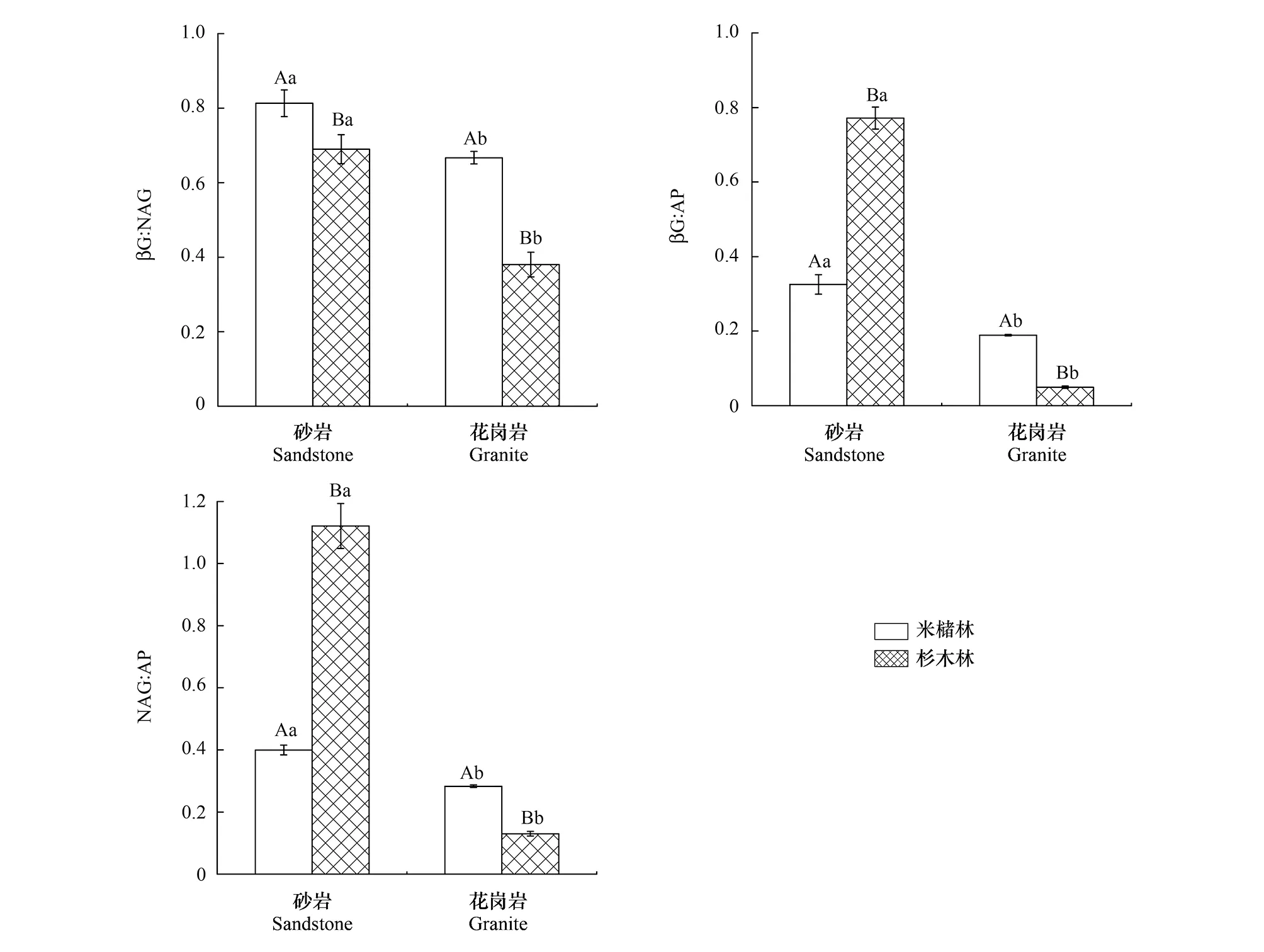
图2 不同森林类型土壤生态酶化学计量比
2.4 土壤生态酶化学计量比与土壤非生物和生物因子的相关性
以土壤酶活性和生态酶化学计量比为响应变量,以生物因子和非生物因子为解释变量进行RDA分析,第一轴解释了变量的77.87%,第二轴解释了变量的19.96%,并且土壤pH、TP和MBC分别解释土壤酶活性和生态酶化学计量比的56.9%、27.9%和12.3%(图3),此外,从RDA分析可知,土壤酶活性和生态酶化学计量比可以明显的将四种林分类型区分开。相关性分析结果表明,土壤βG:AP和NAG:AP与C∶P、N∶P、MBC∶MBP、MBC∶MBN以及砂粒呈显著的负相关关系,土壤βG:NAG、βG:AP和NAG:AP与pH呈显著的负相关关系,TP与土壤βG:NAG、βG:AP呈显著的正相关关系(表4)。
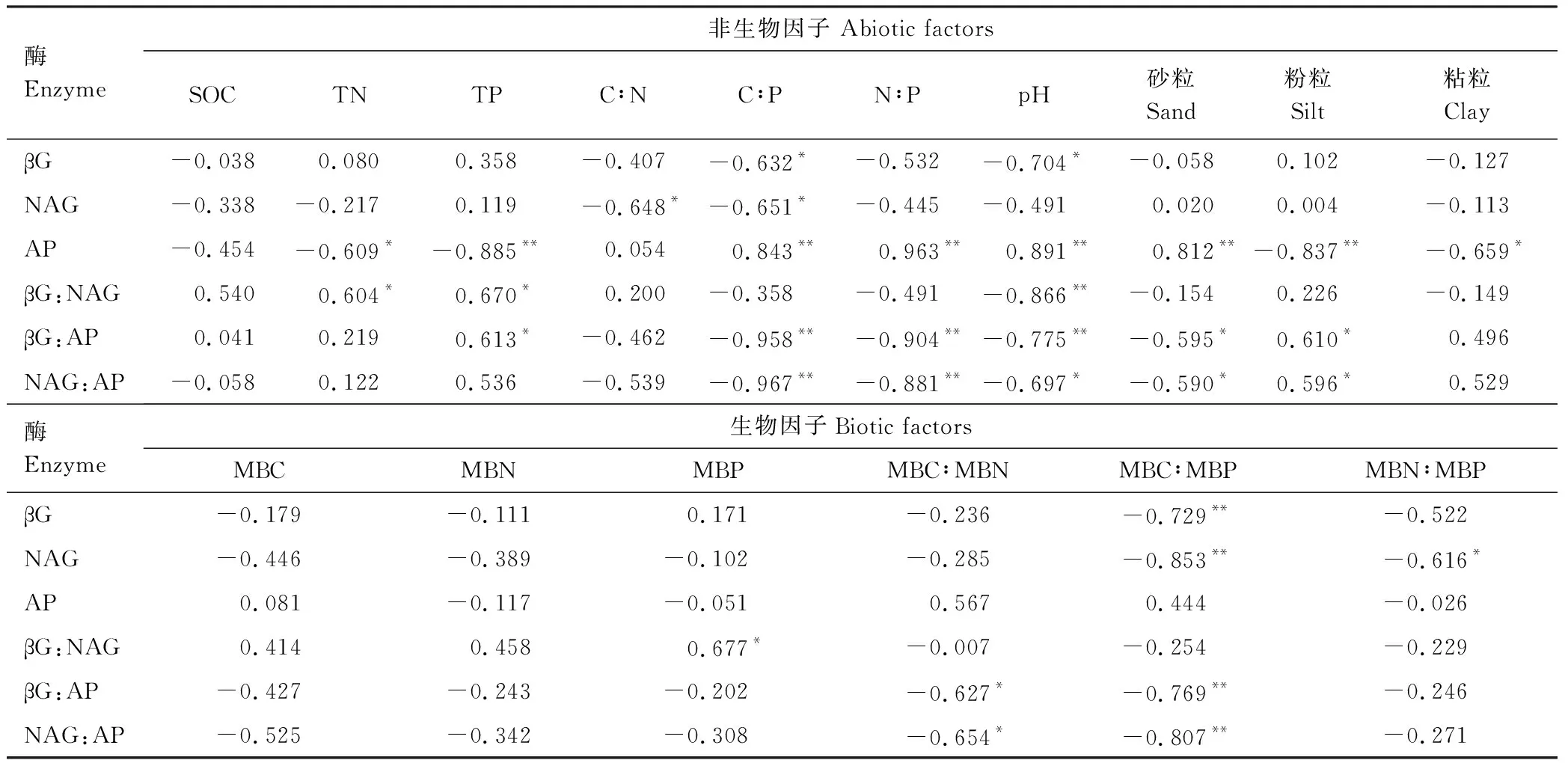
表4 土壤酶活性、生态酶化学计量比与非生物和生物因子的相关性系数
*:P<0.05;**:P<0.01
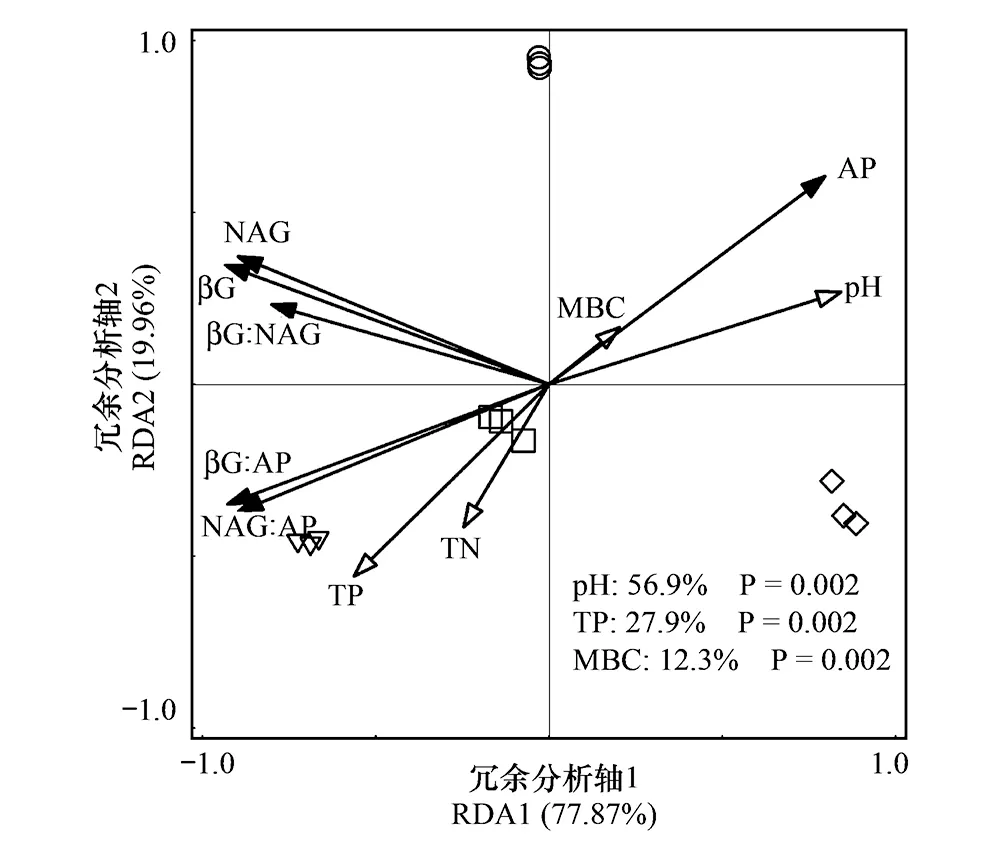
图3 土壤酶活性和生态酶化学计量比与非生物及生物因子的冗余分析
3 讨论
3.1 土壤生态酶化学计量比特征与养分限制
土壤βG∶NAG∶AP对数转换后的比值为1∶1.14∶1.42,与全球生态系统1∶1∶1的比值相偏离,其中βG∶AP平均值为0.33,低于全球水平0.62[7],而NAG∶AP平均值为0.48,与全球水平0.44[7]相接近,说明相对于土壤碳底物有效性来说,本区域土壤微生物可能主要存在一定的磷限制,这与大多数的研究一致,认为热带亚热带地区磷是生态系统生产力的限制因子[20]。Cleveland等[21]在热带施磷实验的结果发现,施磷显著提高土壤呼吸,并且大多数的研究表明酸性磷酸酶(AP)的活性与环境中磷的有效性呈负相关的关系[12]。相关研究发现,施用磷肥可以使土壤中磷的含量增高,而酸性磷酸酶的活性则降低[22]。一般来说,当微生物存在磷限制时,微生物就会分泌更多的磷酸酶促进土壤有机磷矿化,缓解自身的磷限制,微生物分泌的磷酸酶增多,土壤βG∶AP、NAG∶AP的值则降低。
由于土壤酶活性受土壤碳底物和养分的影响,且C、N、P相关的酶活性之间存在权衡关系,因此土壤生态酶化学计量比存在空间的异质性[23- 24]。本研究发现土壤AP的活性显著高于βG、NAG的活性,并且砂岩发育的土壤AP的活性显著低于花岗岩发育的土壤,而砂岩发育的土壤βG∶AP、NAG∶AP则显著高于花岗岩发育的土壤,说明相对于砂岩发育的土壤,花岗岩发育的土壤微生物受磷养分限制的程度更严重,不同母岩发育的土壤,其磷素含量存在一定的差异[25],因此,在一定程度上母质可能是影响土壤磷有效性的关键因子。然而土壤微生物磷养分限制的程度除受母质类型的影响,亦与林分类型密切相关。从花岗岩发育的土壤上来看,本研究发现米槠林βG∶AP和NAG∶AP的值都显著高于杉木林,同时米槠林βG和NAG的活性也显著高于杉木林,这表明在花岗岩发育的土壤上,相对于米槠林,杉木林土壤微生物可能存在磷限制更严重,在碳、磷含量差异不显著的情况下,这可能主要是由于花岗岩发育的杉木林土壤粘粒含量较米槠林高所导致(表2),一般认为土壤粘粒含量越高,土壤对磷吸附能力越强,土壤供磷能力则越弱。但是砂岩发育的土壤则存在相反的趋势,βG∶AP 和NAG∶AP的值都显著低于杉木林,说明相对于砂岩发育的杉木林土壤,米槠林土壤微生物可能存在磷限制更严重,这主要是由于砂岩发育的米槠林土壤微生物可利用的碳底物和氮养分含量显著高于杉木林(表2)。
3.2 非生物和生物因子对生态酶化学计量比的影响
土壤非生物和生物因子通过调节微生物代谢作用,影响土壤酶活性和生态酶化学计量比。冗余分析结果表明土壤pH、总磷(TP)和微生物量碳(MBC)分别解释土壤酶活性和生态酶化学计量比变异的56.9%、27.9%和12.3%,说明土壤非生物因子对土壤酶活性和生态酶化学计量比的影响强于生物因子,这与Peng等[2]在我国不同气候带温带草原及Bowles等[26]在美国加利福尼亚不同管理强度的农业土壤中的研究结果相一致。此外,本研究结果表明土壤pH值是影响土壤生态酶化学计量比的主要因素,并且Pearson相关分析结果发现土壤βG∶AP和NAG∶AP与pH呈显著的负相关关系,说明pH值较大时,βG∶AP和NAG∶AP的值越小,酸性磷酸酶活性相对增高,土壤受到磷养分的限制越严重,这与Xu等[24]对我国不同区域森林土壤的研究结果及Sinsabaugh等[7]全球尺度的Meta分析的结果相一致,其可能原因是土壤pH通过影响土壤微生物生物量和群落组成、酶的空间构象以及酶与土壤颗粒之间的结合状态,从而调控土壤酶活性和生态酶化学计量比[27]。
土壤酶活性及生态酶化学计量比还受土壤总磷(TP)、C∶P及N∶P的影响,相关关系分析结果表明βG∶AP和NAG∶AP分别与C∶P、N∶P呈极显著的负相关关系(表4),并且βG∶AP与土壤总磷呈显著正相关关系,说明土壤生态酶化学计量比亦受土壤养分化学计量比和土壤养分含量的影响,本结果与Xu等[24]和Peng等[2]的研究相一致,其可能原因是土壤酶主要来源于土壤微生物、植物根系分泌、植物降解的产物以及动物残体[28],而微生物生物量化学计量比随土壤和植物组织养分化学计量比的改变而改变,因此土壤生态酶计量比亦能够受土壤养分化学计量比的影响,可以反映土壤养分限制状况[24]。
综上所述,土壤母质可能主要通过影响土壤pH和TP含量而调控土壤酶活性和生态酶化学计量比。一般认为对于全磷含量相差不大的酸性土壤,pH值越高,有效磷含量就越高,而土壤酸性磷酸酶活性则相对更低,但是本研究中土壤pH较高的花岗岩发育土壤磷酸酶活性显著高于砂岩,其可能原因是来自母质风化的土壤总磷含量低于砂岩,微生物可利用磷含量相对较低。森林类型则通过调控土壤C∶P和N∶P调控土壤酶活性和生态化学计量比,其可能原因是不同林分类型的森林凋落物数量和质量的不同,导致土壤有机碳数量和质量、C∶P及N∶P不同,从而改变土壤微生物可以利用C、N、P等底物和养分的数量和质量,影响土壤酶活性和生态酶化学计量比。
4 结论
本研究发现中亚热带地区存在着一定的磷养分限制,花岗岩发育的土壤磷养分限制的强度较砂岩发育的土壤严重,并且砂岩发育的米槠林土壤磷养分限制比杉木林严重,而花岗岩发育的米槠林和杉木林土壤则表现出相反的趋势,因而磷养分限制强度因森林类型和土壤母质的不同而不同。统计分析表明土壤母质可能主要通过影响土壤pH、土壤全磷含量,调控土壤酶活性和生态酶化学计量比,而森林类型可能主要通过改变土壤C∶P和N∶P影响土壤酶活性和生态酶化学计量比,因而本研究建议,未来森林经营及管理应考虑土壤母质和森林类型差异对区域森林土壤养分循环的影响。
猜你喜欢
中国中西医结合皮肤性病学杂志(2022年6期)2023-01-13
落叶果树(2021年6期)2021-02-12
农民致富之友(2020年8期)2020-05-11
文苑(2020年12期)2020-04-13
土壤学报(2019年6期)2020-01-15
海峡姐妹(2019年1期)2019-03-23
现代园艺(2017年23期)2018-01-18
现代园艺(2017年23期)2018-01-18
浙江农业科学(2016年11期)2016-05-04
广西林业科学(2016年2期)2016-03-20
