
不同竹类人工林生态系统生物量空间分配格局
2018-09-10 05:36:30鹿士杨潘丽萍彭晚霞宋同清杜虎刘永贤
生态科学 2018年4期
鹿士杨, 潘丽萍, 彭晚霞, 宋同清, 杜虎, 刘永贤 *
不同竹类人工林生态系统生物量空间分配格局
鹿士杨1, 潘丽萍1, 彭晚霞2, 宋同清2, 杜虎2, 刘永贤1 *
1. 广西壮族自治区农业科学院农业资源与环境研究所, 南宁 530000 2. 中国科学院亚热带农业生态研究所亚热带农业生态过程重点实验室, 长沙 410125 3. 中国科学院环江喀斯特生态系统观测研究站, 广西, 环江 547100
以胸径为单变量, 利用幂函数、指数函数和多项式函数方程模拟麻竹()、毛竹()和粉单竹()林各类器官的生物量, 研究比较了3种不同种类竹林的生物量分配特征。结果表明, 所选模型可以很好的估测竹林及各器官生物量。麻竹、毛竹和粉单竹单株平均生物量为18.160、13.736和4.372 kg, 其林分生物量分别为15.124、28.598和5.102 t·ha-1。竹林单株和林分器官分配一致, 均以竹秆最大, 其次为竹根, 再者为竹枝、竹叶, 但麻竹竹叶>竹枝例外。不同竹林生态系统总生物量与灌木层、草本层、凋落物变化规律一致, 粉单竹>毛竹>麻竹; 但各层生物量分配不同, 麻竹和毛竹表现为乔木层>凋落物层>灌木层、草本层, 乔木层占绝对优势; 而粉单竹表现为草本层>凋落物层>灌木层>乔木层, 且各层次优势不明显。根据单株和林分生物量分配特征, 地上部分和地下部分生物量不均, 应适当调整竹林林分密度和制定合理的采伐措施, 提高竹林的生产力水平, 从而增强其碳储存能力。
生物量; 空间; 分配格局; 生态系统; 竹林
1 前言
森林是陆地上面积最大的生态系统, 森林植被生物量约占陆地生物量的85%[1], 是全球碳循环的重要组成部分[2, 3]。准确估计森林植被的生物量是反映其碳储量[4, 5]及二氧化碳吸收量的研究基础[6]。各国在20世纪60—70年代对包括森林在内的植被生物量进行了大量的调查, 为后来的全球变化研究提供了重要的基本数据。我国森林植被生物量和生产力的研究始于20世纪70年代末80年代初, 冯宗炜等[7]、李文华等[8]率先对中国森林植被生物量进行了测定, 随后, 各地对我国主要森林类型的生物量都有测定, 并就某些森林类型的全国尺度的生物量也分别进行了估算[9-11], 取得了国际上瞩目的成果。然而, 区域尺度某个树种的森林生物量仍有待于精确估算。目前森林生物量的研究更加注重乔木树种, 而竹林生物量的研究较少。
竹是禾本科的竹亚科的总称, 全球有1225种, 主要分布于46°N与47°S之间的热带、亚热带和暖温带地区。竹是中国南方重要的森林资源, 经济用途非常广泛, 是当地农民经济收入的重要来源。竹不仅生命力强、耐旱瘠、成林快、成林后每年均可间伐等特点, 而且能笋材兼用, 出笋多, 同时具有产量高、造纸性能好、无性繁殖力强、适应范围广等优点, 因而竹林二氧化碳固定能力巨大, 对平衡大气二氧化碳具有重要作用。近年来, 我国南方竹林种植面积有不断增大的趋势, 在各国寻找控制或减轻碳释放途径的背景下, 本研究选取麻竹()、毛竹()和粉单竹()3种常见竹林作为研究对象, 通过对这3种竹林生态系统生物量的实测调查, 探讨了不同竹类人工林生态系统生物量分配格局, 以期为进一步开展竹林碳循环、竹林低产林的改造及高效率碳吸存经营措施提供基础数据, 也为正确评价森林在全球碳平衡中的作用提供理论依据, 同时为我国森林生态系统碳平衡的估算和动态模拟提供基础数据, 从而为政府部门制定森林发展规划和环境保护政策提供理论支撑, 提升我国在应对气候变化的科学话语权。
2 研究区概况与研究方法
2.1 研究区概况
研究区位于广西北部、西部和东部竹林主产区, 该区域属中亚热带季风气候区, 具有山地气候一般特征, 气候温暖, 热量丰富, 年平均气温16.5—22 ℃,极端最低气温–8.4至2.9 ℃, 极端最高气温33.7—42.5 ℃, ≥10℃年总积温5000—8300 ℃。年降雨量1300—1800 mm, 主要集中在4—9月, 占全年降雨量的80%, 年总日照1600—1800 h, 年蒸发量1600 mm, 相对湿度74. 8%。
2.2 样方选择与建立
参照《IPCC优良做法指南》对系统随机抽样的建议和广西竹林的分布特征, 基于第八次森林资源清查数据, 计算竹林在广西各县(市)的面积、蓄积综合权重, 选择权重最大的3个县(市) —田林县、兴安县和岑溪县, 相应建立麻竹、毛竹和粉单竹人工林样地, 样地大小为1000 m2(50 m2×20 m2), 3个重复, 共计9个样地。将每块样地进一步划分为10个10 m2× 10 m2的样方, 对样方内测量胸径()≥2 cm竹的胸径、树高、冠幅和坐标进行每木调查, 并挂牌标号, 便于5年后复查。各样地的基本信息见表1。
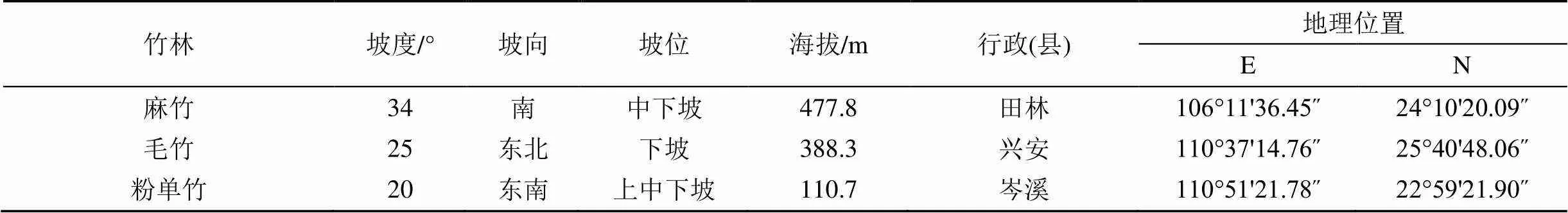
表1 不同竹林人工林样地基本特征
2.3 生物量获取
2.3.1 乔木层
在50 m2×20 m2的样方内, 于每木调查的基础上, 以2 cm为径阶, 根据正态分布原则, 在样地外选择不同径阶的竹共5株作为样木, 进行生物量的测定。测定其基径、胸径、树高和冠幅并按2 m区分段锯断称量, 在树干基部、胸径、中部及顶端分别锯取一个圆盘测定含水量; 并分别称取枝条和叶片的鲜质量; 根的生物量采用全挖法, 并分小根(<2 cm)、中根(2—5 cm)、大根(>5 cm)三组分别称重, 对于丛生的麻竹和粉单竹采取整丛全挖, 根据径阶权重来计算根部重量; 然后对各部分器官分别取样300 g带回实验室, 将样品在105 ℃烘箱内杀青2 h, 调至75 ℃下烘干至恒重, 求出各器官干鲜质量之比, 由此换算出样木各器官的干质量及总干质量, 得出分株各构件生物量分配=构件生物量/分株生物量×100%。利用样木各器官(干、枝、叶、根)的干重和总干重建立各器官及总生物量与样木胸径()的幂回归方程(W=aD)、多项式方程(W=a+b1D+ b2D+b3D)或指数方程(W=aeD)。根据样地每木调查的结果及相应的回归方程计算乔木层各个体的生物量, 并由此获得乔木层的总生物量。
2.3.2 灌木层、草本层和地上凋落物
按“品”字型在样地1、5、8号10 m2×10 m2的小样方各设置1个2 m2×2 m2的样方, 调查所有灌木种类、株丛数、高度、地径、覆盖度, 在每个2 m2×2 m2的样方内取1个1 m2×1 m2的小样框, 调查草本种类、株丛数、平均高度、覆盖度。为了不破坏复查样地, 在相距各样框5 m的范围内选择1个与灌木调查样框植被类型、立地条件等基本一致的样框按全收获法收获样框内所有灌木的枝、叶、花果、根, 测定各器官生物量, 并以3个样框的平均值来推算样地中灌木层的总生物量。按同样方法在1 m2×1 m2小样框内获取草本层植物各器官、凋落物生物量及总生物量。
2.4 数据处理
所有数据及多重比较分析、回归方程模型建立均在Excel—2003和SPSS 16.0中进行处理。
3 结果与分析
3.1 乔木层生物量回归模型的建立
虽然同时利用胸径()和树高()2个变量模拟单木生物量优于以单因子为自变量的模拟结果[12]。但鉴于树高估测误差较大, 本文采用以树木胸径()为单变量的幂函数(=aD)、指数函数(=ae)或多项式(=+1+22+33)函数方程, 不同器官生物量估算模型不同, 不同竹类同一器官生物量估算模型也不同。根据相关系数和显著性检验, 麻竹的整株生物量、粉单竹叶、枝和整株生物量的回归模型为=ae, 麻竹叶、毛竹秆、根、整株和粉单竹秆生物量的回归模型为=aD, 其他的则为=+1+22+33(表2)。经统计学检验除毛竹叶和粉单竹根外其他生物器官生物量数学模型的相关系数在0.407—0.954之间, 经t检验均达到显著(p<0.05)或极显著水平(p<0.01), 表明所选模型可以对竹林生物量进行估测。
3.2 单株生物量和分株各构件生物量分配
不同竹类单株生物量和分株构件生物量分配不同(图1)。单株生物量表现为麻竹(18.16 kg) >毛竹(13.736 kg)>粉单竹(4.372 kg)(<0.01, 图1a); 各构件生物量在麻竹中最大、粉单竹最小(<0.01, 图1b); 叶生物量分配在麻竹最高(20.339%)(<0.01), 枝生物量分配为粉单竹(9.184%)>麻竹(7.843%)>毛竹(3.726%)(<0.01), 秆生物量分配为粉单竹(68.899%)、毛竹(67.818%)>麻竹(49.453%) (< 0.01), 根生物量分配以毛竹(26.607%)最高, 显著高于粉单竹(16.556%)(图1c)。
3.3 不同竹林乔木层生物量及其分配
不同竹林不同器官生物量分配不同(表3)。3类竹林林分生物量5.102—28.598 t·ha-1, 其中毛竹最大, 显著高于麻竹, 极显著高于粉单竹; 3类竹林各器官生物量基本表现为竹秆>竹根>竹枝>竹叶。不同竹林竹秆生物量最大, 所占乔木层生物量比例也最大, 为50.99%—70.65%, 可见竹类器官生物量主要集中在竹秆中; 毛竹竹秆生物量最高19.303 t·ha-1, 极显著高于粉单竹 (4.324 t·ha-1), 显著高于麻竹(7.663 t·ha-1)。竹根生物量次之, 毛竹竹根生物量达6.434 t·ha-1, 显著高于麻竹和粉单竹, 后两者差异不显著; 粉单竹竹根生物量占竹林生物量比例最高, 达29.06%。竹叶和竹枝生物量较小, 毛竹和粉单竹均显著低于麻竹, 与麻竹不同的是, 毛竹和粉单竹竹枝生物量高于竹叶。不同竹林地上部分生物量占林分生物量70.94%—78.11%, 地下部分生物量占21.89%—29.06%。

表2 不同竹类各器官生物量估算模型
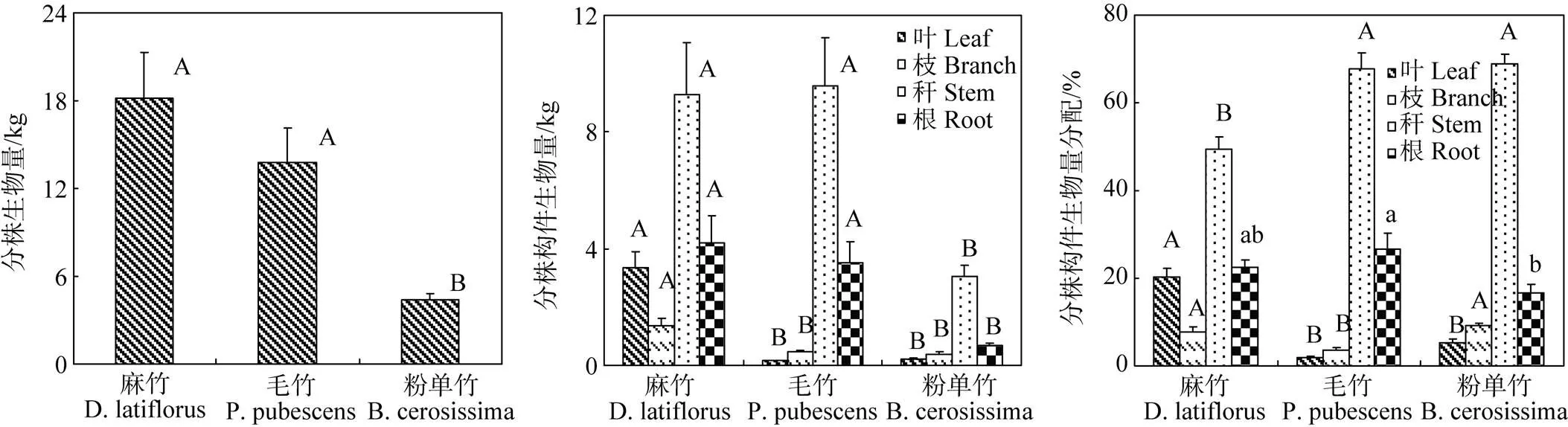
#不同大小写字母标记的值之间差异极显著(p<0.01)或者显著(p<0.05)。
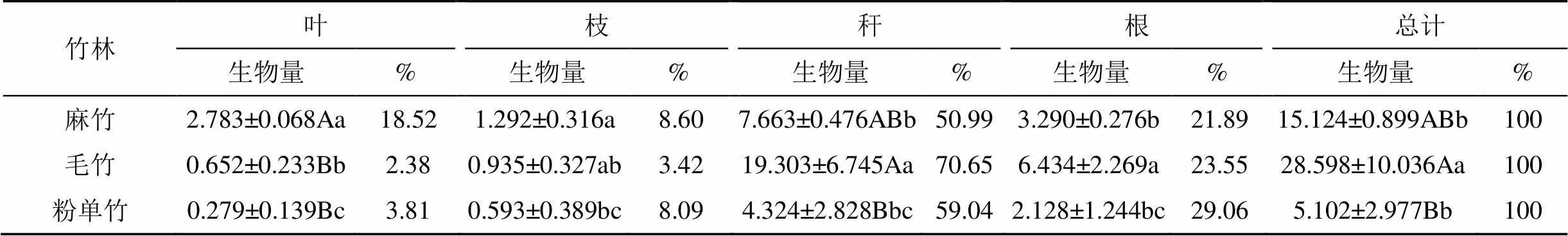
表3 不同竹林乔木层生物量器官分配
3.4 不同竹林灌木层生物量及其分配
不同竹林灌木层生长发育与竹的种类生物特性密切相关, 因此, 不同竹林灌木层生物量分配格局不同(表4)。麻竹中灌木层基本没有, 也就没有生物量; 灌木层各器官生物量的分配比例在毛竹和粉单竹林有所不同, 规律性不强。粉单竹灌木层生物量较大, 达11.995 t·ha-1, 灌木枝生物量所占灌木层生物量比例最大, 为43.77%, 其次为灌木根(32.67%)和灌木叶(23.65%); 毛竹灌木枝生物量为3.995 t·ha-1, 根所占比例最大, 达56.85%, 其次为灌木枝, 再者为灌木叶。
3.5 不同竹林草本层及凋落物生物量
不同竹林草本层和凋落物生物量分配不同, 其大小顺序一致, 依次为粉单竹、毛竹和麻竹(表5)。粉单竹林生物量较小, 可利用空间相对较大, 草本层生物量最大, 达16.19 t·ha-1, 其凋落物也最大; 尽管毛竹林分生物量最大, 其草本层和凋落物生物量也不小, 介于粉单竹和麻竹之间; 麻竹林草本层生物量最小, 因为麻竹林叶片比较繁密, 遮住了大量阳光, 导致林下发育不好。草本层除麻竹地上生物量<地下生物量外, 毛竹和粉单竹均表现为地上生物量>地下生物量。
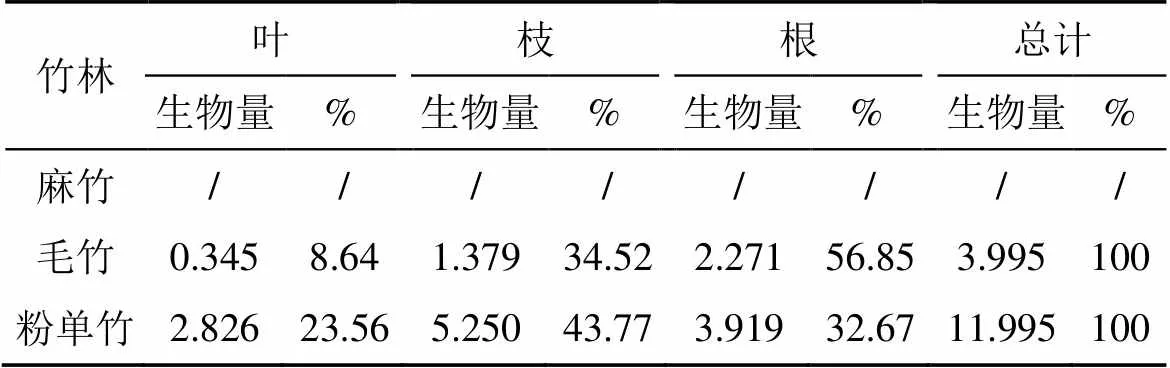
表4 不同竹林灌木层生物量器官分配
3.6 不同竹林总生物量组成及其分配
不同竹林总生物量变化规律与灌木层、草本层和凋落物一致(表6、表5、表4), 粉单竹>毛竹>麻竹, 其中活体贡献率为73.72%—79.27%, 凋落物贡献率为21.73%—26.28%。不同林分各层生物量分配不同。麻竹和毛竹表现为乔木层>凋落物层>灌木层、草本层, 乔木层占绝对优势; 而粉单竹表现为草本层>凋落物层>灌木层>乔木层, 且各层次优势不明显。
4 讨论
4.1 竹林生物量回归方程
生物量模型估算法是目前广泛采用的研究森林生物量的一种有效方法。它利用林木易测因子(如胸径和树高)来推算林木生物量, 从而减少野外工作量。目前, 采用的生物量模型很多[13, 14], 包括线性模型、幂函数模型、多项式和指数模型等, 常用的自变量有胸径()、2、树高()、和2等[15-17]。因树高估测误差较大, 本文仅采用以树木胸径()为变量的模拟竹林生物量, 很多研究者认为竹林地上部分生物量及各构件生物量与径级的回归程度明显高于高度[18, 19]。竹类是不同年龄立竹构成的异龄林, 由于生长年限的不同, 各年龄的竹株光合作用时间和能力有所差异, 从而, 其生物量及其分配存在差异[20], 不同年龄竹生物量的分配是不同的[21], 杨春花等[19]研究验证了这一点。本研究中竹林不同器官生物量估算模型不同, 3种竹类同一器官生物量估算模型也不同。麻竹的整株生物量、粉单竹叶、枝和整株生物量的回归模型指数函数, 麻竹叶、毛竹秆、根、整株和粉单竹秆生物量的回归模型为幂函数, 其他的则为多项式。经统计学检验除毛竹叶和粉单竹根外其他生物器官生物量数学模型的相关系数均达到显著或极显著水平, 说明所选模型可以较准确地预测竹林生物量。
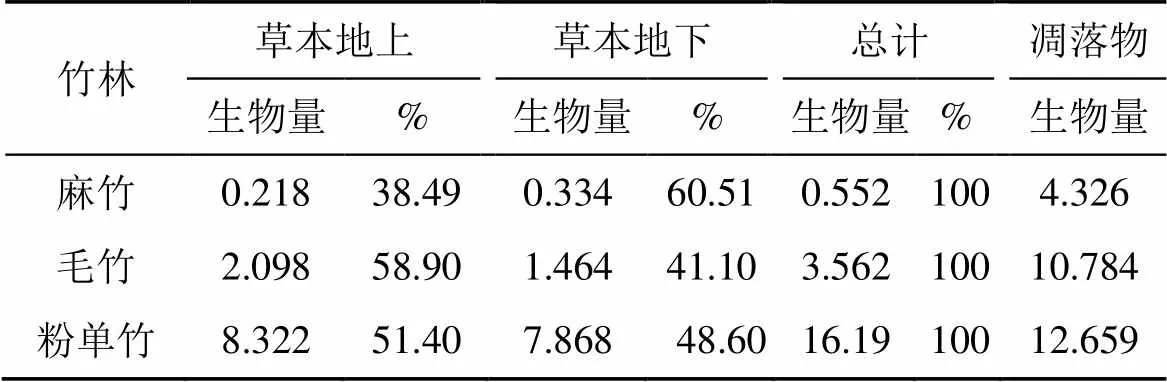
表5 不同竹林草本层、凋落物层生物量分配
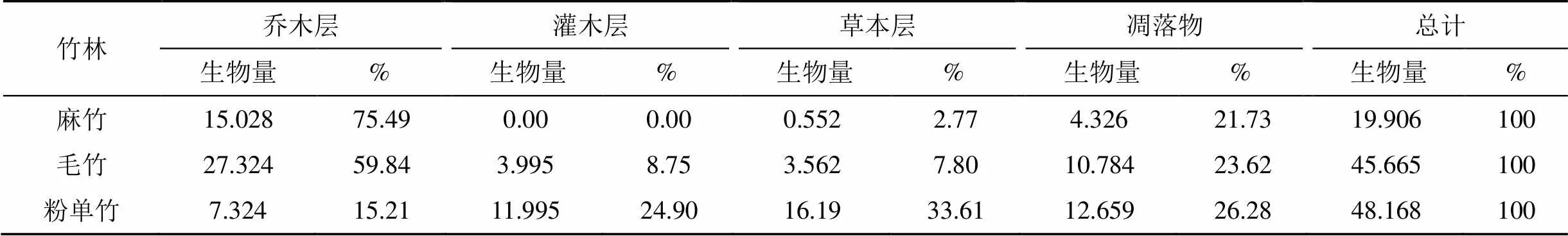
表6 不同竹林各层生物量分配
4.2 单株生物量分配格局
生物量分配格局是植物应对异质性生境的重要途径, 同时也能反应环境中可利用资源的变化[22]。生物量在各器官的分配比例因树种的不同而有差异, 反映了植物对环境资源的不同的利用方式[23]。竹枝、竹秆、竹叶为竹子地上部分主要的营养器官, 各器官生物量的合理分配, 对提高林分的生物量及出材量具有重要意义。本研究表明, 不同竹类单株各器官生物量在空间上的配置不同, 麻竹为竹秆>竹根>竹叶>竹枝, 毛竹为竹秆>竹根>竹枝>竹叶, 粉单竹竹秆>竹根>竹枝>竹叶, 与竹种不同有关。秆是乔木层生物量比例最大的器官, 本研究中3类竹秆比例达到50.99%—70.65%, 从而影响分株地上部分生物量和单位面积上地上部分总生物量。叶片是植物光合作用的主要器官, 对大多数植物而言, 叶片越多, 其光合作用产生的有机物质越多, 植物生长越旺盛。麻竹叶的生物量分配最高, 达20.339%, 其单株生物量最高。与麻竹、粉单竹不同, 毛竹竹叶和竹枝的比例为5.58%, 竹根比例高达26.61%, 两者所占比例极不平衡, 可能是人为采伐和栽植密度有关。当地上部分(枝、叶)和地下部分生物量比例基本一致, 有利于植物的生长, 因此, 应调整竹林林分密度和制定合理的采伐措施, 以调整各器官生物量的分配比例。
4.3 竹林不同层次生物量分配格局
不同竹林生物量不同, 其生物量分配格局也不一样。麻竹、毛竹和粉单竹生物量分别为19.906、45.665和48.168 t·ha-1, 表现为粉单竹>毛竹>麻竹, 与草本层和凋落物生物量的变化规律一致(表6、表5), 其中活体贡献率为73.72%—79.27%, 凋落物贡献率为21.73%—26.28%。麻竹和毛竹林生物量分配表现为乔木层>凋落物层>灌木层、草本层, 乔木层占绝对优势; 而粉单竹表现为草本层>凋落物层>灌木层>乔木层, 且各层次优势不明显。虽然麻竹乔木层生物量远高于粉单竹, 由于麻竹竹叶和竹枝生长太旺盛, 覆盖度较高, 吸收了绝大部分光能, 以致林下灌木、草本发育不好, 凋落物也不多, 从而使得整个竹林生物量最少; 虽然粉单竹其乔木层生物量显著小于麻竹和毛竹, 但其灌木层和草本发育良好, 凋落物多, 后三者的生物量均高于乔木层生物量, 竹林整个生物量达到最高; 毛竹介于其中。3类竹林同一器官生物量分配不同, 可能与竹种类及人类无规律的掠夺式采笋干扰等有关。
4.4 人为干扰对竹林生物量的影响
现存毛竹林的生物量(45.665 t·ha-1)小于浙江临安青山和三口2个乡镇的毛竹林生物量(60.65 t·ha-1)[24]和湖南会同林区毛竹林生物量(57.51 t·ha-1)[25]。现存麻竹生物量远小于毛竹和粉单竹, 这可能因为麻竹能笋材兼用, 农民每年对麻竹进行笋材的采伐, 导致生物量降低; 虽然毛竹也笋材兼用, 但生物量远高于麻竹, 这与人为干扰较少、竹林管理有关; 虽然粉单竹林乔木层生物量最小, 干扰较小, 其灌木层和草本层发育较好, 枯枝凋落物也较多, 从而使得整个竹林生物量最高。竹林是异龄林分, 通常采取择伐作业, 隔年伐去3年以上竹。因此, 从生态系统的角度来看, 竹林永远处于生长动态平衡之中, 并可以近似认为每次采去竹林的生物量相当于现存生物的1/4[26]。由此换算, 麻竹、毛竹和粉单竹分别被砍伐掉3.757、6.831和1.831 t·ha-1, 总的生物量分别为23.663、52.496和49.999 t·ha-1。可见, 人为干扰对竹林的生产力产生重要影响。
5 结论
(1)本研究中麻竹的整株生物量、粉单竹叶、枝和整株生物量的回归模型为指数函数, 麻竹叶、毛竹秆、根、整株和粉单竹秆生物量的回归模型为幂函数, 其他的则为多项式。经统计学检验除毛竹叶和粉单竹根外其他生物器官生物量数学模型的相关系数均达到显著或极显著水平, 说明所选模型可以较准确地预测竹林生物量, 对今后的竹林调查及研究能提供数据支撑。
(2)不同竹类单株各器官生物量在空间上的配置不同, 麻竹为竹秆>竹根>竹叶>竹枝, 毛竹为竹秆>竹根>竹枝>竹叶, 粉单竹竹秆>竹根>竹枝>竹叶。地上部分(枝、叶)和地下部分生物量比例相差较大, 应调整竹林林分密度和制定合理的采伐措施, 以调整各器官生物量的分配比例。
(3)不同竹林生物量不同, 其生物量分配格局也不一样。3类竹林同一器官生物量分配不同, 可能与竹种类及人类无规律的掠夺式采笋干扰等有关。通过对竹林的合理经营和管理, 可以提高竹林的生产力水平, 从而增强其系统的碳储存能力。
[1] LIETH H, WHITTAKER R H. Primary Productivity of the Biosphere[M]. New York: Springer-Verlag, 1975: 89–95.
[2] SCURLOCK J M O, CRAMER W, OLSON R J, et al. Terrestrial npp: toward a consistent data set forglobal model evaluation[J]. Ecological Applications, 1999, 9(3): 913– 919.
[3] FANG Jingyun, CHEN Anping, PENG Changhui, et al. Changes in forest biomass carbon storage in China between 1949 and 1998[J]. Science, 2001, 292(5525): 2320–2322.
[4] 杨昆, 管东生. 珠江三角洲地区森林生物量及其动态[J]. 应用生态学报, 2007, 18(4): 705–712.
[5] 黄从德, 张健, 杨万勤, 等. 四川省及重庆地区森林植被碳储量动态[J]. 生态学报, 2008, 28(3): 966–975.
[6] 于贵瑞. 全球变化与陆地生态系统碳循环和碳蓄积[M]. 北京: 气象出版社, 2003: 56–66.
[7] 林生明, 徐土根, 周国模. 杉木人工林生物量的研究[J]. 浙江农林大学学报, 1991, 26(3): 288–294.
[8] 李文华, 邓坤枚, 李飞. 长白山主要生态系统生物量生产量的研究[J]. 森林生态系统研究, 1981, 2(34): 22.
[9] 刘世荣, 柴一新, 蔡体久, 等. 兴安落叶松人工群落生量物与净初级生产力的研究[J]. 东北林业大学学报, 1990, (2): 40–46.
[10] 周玉荣, 于振良, 赵士洞. 我国主要森林生态系统碳贮量和碳平衡[J]. Journal of Plant Ecology(植物生态学报(英文版)), 2000, 24(5): 518–522.
[11] WANG Xiaoke, FENG Zongwei, OUYANG Zhiyun. The impact of human disturbance on vegetative carbon storage in forest ecosystems in China[J]. Forest Ecology & Management, 2001, 148(1–3): 117–123.
[12] 李轩然, 刘琪璟, 陈永瑞, 等. 千烟洲人工林主要树种地上生物量的估算[J]. 应用生态学报, 2006, 17(8): 1382– 1388.
[13] 冯宗炜, 陈楚莹, 张家武, 等. 湖南会同地区马尾松林生物量的测定[J]. 林业科学, 1982, 18(2): 127–134.
[14] 吕晓涛, 唐建维, 何有才, 等. 西双版纳热带季节雨林的生物量及其分配特征[J]. 植物生态学报, 2007, 31(1): 11– 22.
[15] MONSI M. Mathematical models of plant communities[M]. Paris: Univ. , Fac. of Science, Department of Botany, 1968: 107–120.
[16] 王维枫, 雷渊才, 王雪峰, 等. 森林生物量模型综述[J]. 西北林学院学报, 2008, 23(2): 58–63.
[17] 李燕, 张建国, 段爱国, 等. 杉木人工林生物量估算模型的选择[J]. 应用生态学报, 2010, 21(12): 3036–3046.
[18] 董文渊, 黄宝龙, 谢泽轩, 等. 筇竹无性系种群生物量结构与动态研究[J]. 林业科学研究, 2002, 15(4): 416–420.
[19] 杨春花, 周小平, 王小明. 卧龙自然保护区华西箭竹地上生物量回归模型[J]. 林业科学, 2008, 44(3): 113–123.
[20] 林益明, 林鹏. 绿竹种群生物量结构研究[J]. 竹子研究汇刊, 1998(2): 9–13.
[21] 薛萍, 汪晓萍, 李锡泉. 湖南衡阳紫色页岩地区芦竹生物量的动态规律[J]. 中南林业科技大学学报, 2006, 26(3): 11–14.
[22] HILBERT D W, CANADELL J. Biomass Partitioning and Resource Allocation of Plants from Mediterranean-Type Ecosystems: Possible Responses to Elevated Atmospheric CO2[M]. New York: Springer, 1995: 1495–1502.
[23] TILMAN D. Plant strategies & the dynamics & structure of plant communities[M]. Princeton, New Jersey: Princeton University press, 1988: 152–160.
[24] 周国模, 姜培坤. 毛竹林的碳密度和碳贮量及其空间分布[J]. 林业科学, 2004, 40(6): 20–24.
[25] 肖复明, 范少辉, 汪思龙, 等. 毛竹()、杉木()人工林生态系统碳贮量及其分配特征[J]. 生态学报, 2007, 27(7): 2794– 2801.
[26] 肖复明, 范少辉, 汪思龙, 等. 毛竹、杉木人工林生态系统碳平衡估算[J]. 林业科学, 2010, 46(11): 59–65.
Characteristics of biomass allocation in different bamboo plantations
LU Shiyang1, PAN Liping1, PENG Wanxia2, SONG Tongqing2, DU Hu2, LIU Yongxian1*
1. Agricultural Resource and Environment Research Institute, Guangxi Academy of Agricultural Sciences, Nanning 530000, China 2. Key Laboratory of Agro-ecological Processes in Subtropical Region, Institute of Subtropical Agriculture, Chinese Academy of Sciences, Changsha 410125, China 3. Huanjiang Observation and Research Station of Karst Ecosystem, Chinese Academy of Sciences, Huanjiang 547100, Guangxi, China
Based on the data of diameter at the breast height (DBH), characteristics of biomass allocation in different bamboo plantations were studied using power function, exponential function, and polynomial equation, respectively. Three types of bamboo plantations were selected, i.e.,, and. The results indicated that biomass allocation in bamboo organs could be precisely predicted by the models. Individual plant biomass of,, andwere on average 18.160, 13.736, and 4.372 kg, respectively. While the total biomass of three bamboo plantations was 15.124, 28.598, and 5.102 t·ha-1, respectively. Biomass allocation in individual bamboo plant for the three bamboo typesfollowed the order of stem > root > branch > leaf, except forin which the biomass of leaf was larger than branch. The total biomass of the three bamboo plantation ecosystems was in the order of>>, and the same pattern was observed in shrub layer, herbaceous layer, and litter layer. However, the biomass allocation in each layer differed among the three bamboo plantations. For example, biomass allocation followed the order of canopy > litter layer > shrub layer > herbaceous layer inandplantations, while followed the order of herbaceous layer > litter layer > shrub layer > canopy inplantations. The results suggested that, to enhance the productivity of bamboo plantation and increase carbon sequestration, it should adjust bamboo stand density and develop rational harvest regimes according to biomass allocation.
biomass; spatial pattern; allocation; ecosystems; bamboo plantation
10.14108/j.cnki.1008-8873.2018.04.015
Q948
A
1008-8873(2018)04-123-07
2017-03-17;
2017-12-29
国家自然科学基金项目地区科学基金项目(31660141); 国家重点研发计划(2016YFC0502405); 广西重点研发计划(桂科AB16380255, 桂科AB17129009)
鹿士杨(1985—), 男, 硕士, 实习研究员, 主要从事土壤微生物以及土壤修复研究, Email:lusyang@yeah.net
刘永贤, 男, 硕士, 副研究员, 主要从事土壤修复以及富硒农产品研究, E-mail: liuyx27@163.com
鹿士杨, 潘丽萍, 彭晚霞, 等. 不同竹类人工林生态系统生物量空间分配格局[J]. 生态科学, 2018, 37(4): 123-129.
LU Shiyang, PAN Liping, PENG Wanxia, et al. Characteristics of biomass allocation in different bamboo plantations[J]. Ecological Science, 2018, 37(4): 123-129.
猜你喜欢
中华建设(2024年3期)2024-03-16 08:04:52
乡村科技(2023年20期)2024-01-23 16:39:18
中华建设(2023年10期)2023-10-07 05:23:04
少儿美术(2020年8期)2020-11-06 05:21:38
意林·少年版(2020年13期)2020-08-02 11:02:50
现代园艺(2017年14期)2017-02-05 00:38:45
广西林业科学(2016年4期)2016-03-16 05:44:55
中国农业信息(2016年10期)2016-02-06 14:54:04
福建农业科技(2015年1期)2015-02-27 10:20:39
福建农业科技(2015年1期)2015-02-27 10:20:38
