
梭梭气体交换特征对人工模拟增温的响应
2018-09-10 07:11:24李桂芳王仲科
江苏农业科学 2018年15期
张 维, 庄 丽, 李桂芳, 王仲科, 黄 刚
(石河子大学生命科学学院,新疆石河子 832000)
植物的光合作用是地球上最重要的生物化学反应之一[1],极易受到许多环境因子的影响,其中温度就是一个很重要的方面[2]。温度升高对植物的影响可分为直接影响和间接影响,直接影响主要包括温度升高将改变植物的光合特性及生长速率;间接影响则主要包括改变植物的土壤特性和对营养物质的利用[3-4]。迄今为止,大量的研究主要集中在低温下植物的光合机理方面,对于高温响应的研究比较少。温室效应引起的全球性气候变暖在很大程度上直接影响植物的生长发育及其生存状况。因此,研究模拟增温对植物的影响对于促进生态平衡发展具有重要的理论价值和生态学意义。
梭梭(Haloxylonammodendron)为藜科梭梭属灌木或小乔木,属亚洲荒漠成分,广泛分布于中亚荒漠地区,具有较强的抗旱、抗寒、耐盐碱、耐瘠薄等生态适应特征[5],新疆尤以准噶尔盆地为主[6]。由于其所具有的经济价值和生态价值而遭到大规模的开发利用,从而导致原生天然梭梭林很快被大面积破坏,物种多样性急剧降低,在一段时间内处于濒危状态,因此,在1984年该种被列为国家濒危三级保护植物。同时也正是因为梭梭这些巨大的经济价值和灭绝风险,才引起了政府、专家及学者的高度重视与关注。迄今为止,已从微观的分子水平[7-8]、生理水平[9-10]及宏观的梭梭种人工培育种植[7,11]、梭梭林的更新修复技术[12-13]等多个方面展开了较为系统的研究。所获得的研究成果也为今后如何更合理地开发利用及科学保护梭梭物种提供了丰富的科学依据,但有关梭梭光合生理特征方面,则更多的是围绕梭梭对水分需求及土壤营养状况方面展开,而对全球性气候变暖后梭梭光合特征变化及其未来生存状况方面的研究尚未见报道。
对此,为尽量避免人为破坏梭梭生境的前提下,本研究采用开顶箱(open top chamber,OTC)增温的方法来进行试验温度变量的控制,与此同时,对于试验全过程所需获取的各项数据都通过探头监测技术而获得,这就意味着所得数据与梭梭真实的各项生理反应参数更接近,也更具说服力和参考价值,且在具体的试验操作过程中,还模拟全球气候变化,对梭梭生长环境下的土壤进行了氮(N)、磷(P)等营养成分及水分含量等相关分析。本研究主要是在控制其他各处理因素相同的情况下,就温度变化对梭梭光合特征的影响方面而深入展开探讨,旨在为认识梭梭在全球气候变化下的生存策略的适应性变化及对梭梭未来的生存状况提供一定的科学依据。
1 材料与方法
1.1 研究区域概况
本试验选址为中国科学院新疆生态与地理研究所阜康荒漠生态研究站(87°45′ E、44°30′ N,海拔436 m)。地处天山北麓、准噶尔盆地南缘,该地区年平均降水量 117.2 mm,年平均蒸发量1 942.1 mm,年平均气温6.1 ℃,属温带大陆性干旱荒漠气候。植被属典型荒漠灌木、半灌木及半乔木的单生或混生稀疏植被,梭梭是该区主要的关键种或建群种稀疏植被之一。
1.2 试验样地设置
本试验样地设置于研究站试验基地的围栏沙地内,选取地势平坦、分布相对均一的梭梭植被地作为试验样地。在试验样地内随机设置OTC开顶箱增温样地(上下端直径均为 4 m、高为2 m、边长为1 m的正十二边形)和常温样地(边长为10 m的正方形)各8个,且通过温度探头监测开顶箱内外的温度差,计算平均增温值。为了便于定期地观察和采样,在开顶箱的南、北侧各开1个高2 m、宽1 m的小门。
1.3 光合参数日变化及环境因子测定
气体交换参数的测定于梭梭生长季(7月),选择晴天,北京时间08:00—20:00进行,每2 h测定1次。测定方法为随机选取常温组与增温组中生长良好的梭梭充分展开同化枝,利用美国Li-6400 XT便携式光合测定系统仪测定,且每次每株同化枝连续重复测取5个数值,共测取3组,取其平均值。整个观测全过程均采用光合仪系统自动记录同步的各项参数数值,主要包括梭梭同化枝净光合速率(Pn)、蒸腾速率(Tr)、胞间二氧化碳浓度(Ci)、气孔导度(Gs)、水气压亏缺(VPD)以及大气二氧化碳浓度(Ca)、光合有效辐射(PAR)、气温(Ta)和相对湿度(RH)。而其同化枝瞬时水分利用效率(WUE)与气孔限制值(Ls)则通过以下公式计算得出:WUE=Pn/Tr[14];Ls=1-Ci/Ca[15]。所取土样则采用烘干法测定其含水率,电位法(水土比为5 ∶1)测定其酸碱度(pH值)和电导率(EC)。
1.4 数据分析
利用SPSS 17.0统计分析软件对2种生境下的各环境因子及梭梭气体交换参数进行配对t检验;利用Pearson法对梭梭Pn、Tr与各环境因子间的关系进行相关性分析,若P<0.05,则认为两者间存在显著差异,同时结合Origin 8.0进行制图。
2 结果与分析
2.1 梭梭不同生境土壤特性及微气象因子日变化
由表1可以看出,梭梭生长地的土壤含水量极度缺乏,常温、增温处理分别为0.769%、0.656%,土壤电导率分别为140.2、136.9 μS/cm,土壤pH值分别为9.55、9.35,属于典型的荒漠干旱性碱性土类型。因此,只有极度耐干旱耐盐碱的植物种才能在此生境中生长。

表1 不同样地的土壤特性
注:同列数据后不同小写字母表示差异显著(P<0.05),不同大写字母表示差异极显著(P<0.01)。
从梭梭生长的环境因子日变化结果可以看出,在整个观测期内PAR和Ta的日变化均为“单峰”型,RH则为“W”形,Ca为“S”形(图1)。2种生境的环境因子在此过程中的变化规律基本相同,如2种生境中的PAR均随着太阳高度的变化而变化,从上午最初观测时开始上升,至14:00太阳高度最高,分别为1 922.6 、1 909.5 μmol/(m2·s),之后又缓慢降低,并使得增温处理于20:00左右降至整个观测期的最低值(图1-a)。Ta主要受PAR的影响,分别观测到的最低Ta为08:00时的24.7、31.1 ℃左右。此后,由于光照度的逐渐增加而迅速上升,使得常温与增温下Ta分别于18:00和16:00左右达到1 d内的最高温度,较PAR而言,其各自峰值分别滞后了4、2 h左右,之后则随着PAR的减弱而下降(图1-b)。RH则受PAR和Ta的双重影响,使得其日变化观测过程中的最高值分别出现在08:00和10:00,约为29.3%和27.9%,最低值出现在12:00,约为16.7%和11.4%,其中增温下RH在14:00有较小幅度的剧烈回升,后下降,之后才随着PAR和Ta的下降逐渐回升,且变化幅度较大(图1-c)。而Ca最初2 h则降低,于10:00降至峰谷值,其大小分别为265.0、240.0 μmol/mol,随后又逐渐升高,直至18:00,最后急剧下降(图1-d)。
2.2 不同生境下梭梭气体交换的日变化
通过图2-a可知,在整个试验观测期内,2种生境下梭梭同化枝净光合速率(Pn)的日变化趋势均呈典型的“双峰”型。其具体变化情况从最初的08:00迅速升高,至10:00左右出现第1个峰值,分别为17.1、16.8 μmol/(m2·s),随后出现急剧下降,约在14:00、16:00分别降至峰谷值,其值大小为4.5、3.6 μmol/(m2·s),之后均逐渐回升,约于18:00左右出现第2个峰值,分别为8.4、4.8 μmol/(m2·s);最后呈下降趋势,其明显的午间降低即所谓的“午休”现象。同时,从图2-a也可看到增温下梭梭同化枝Pn于10:00后随着温度的继续升高而逐渐低于常温组,虽分析结果并不显著(P>0.05),但却从一定程度上表明了增温对梭梭光合作用进行所造成的负效应。
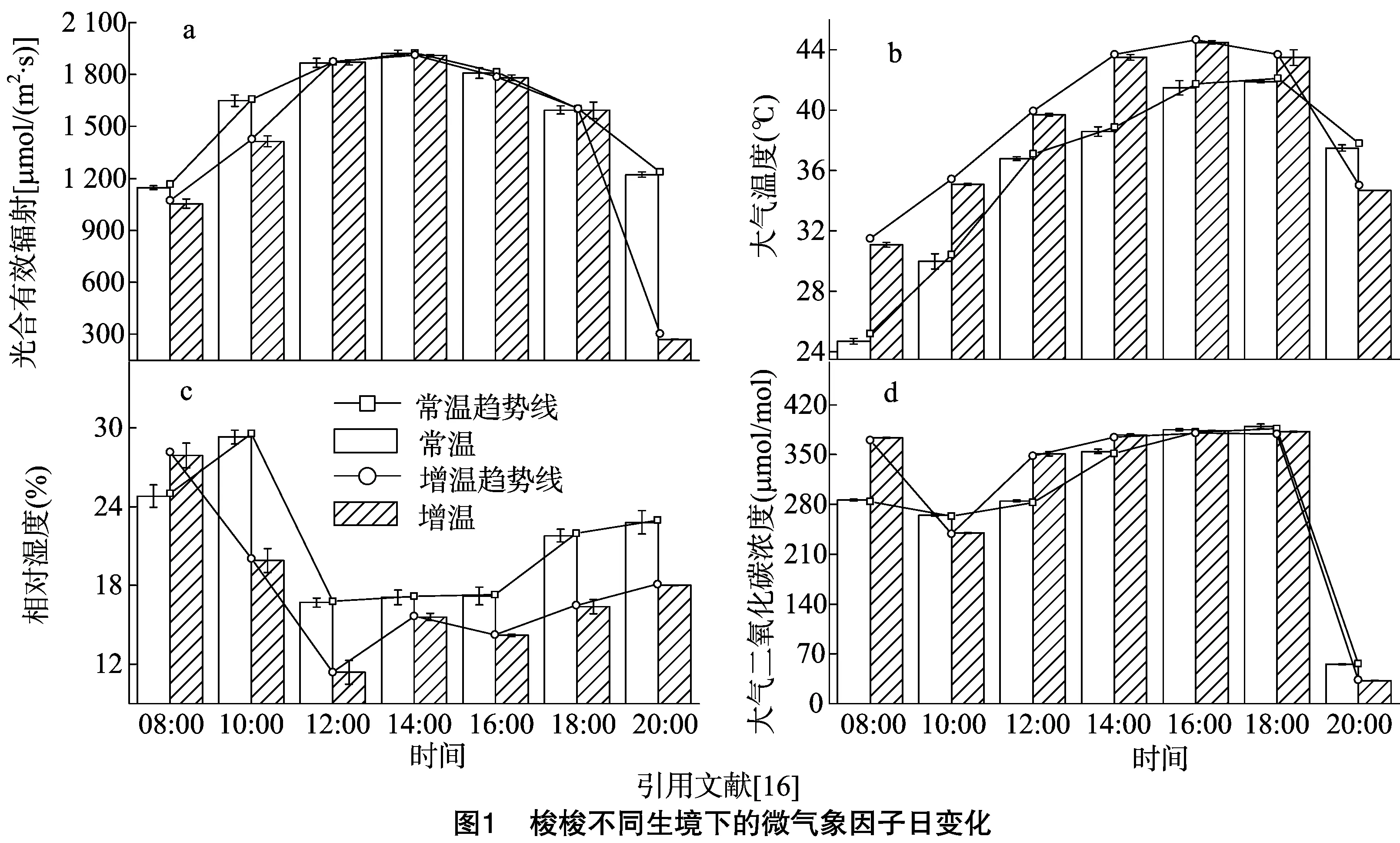
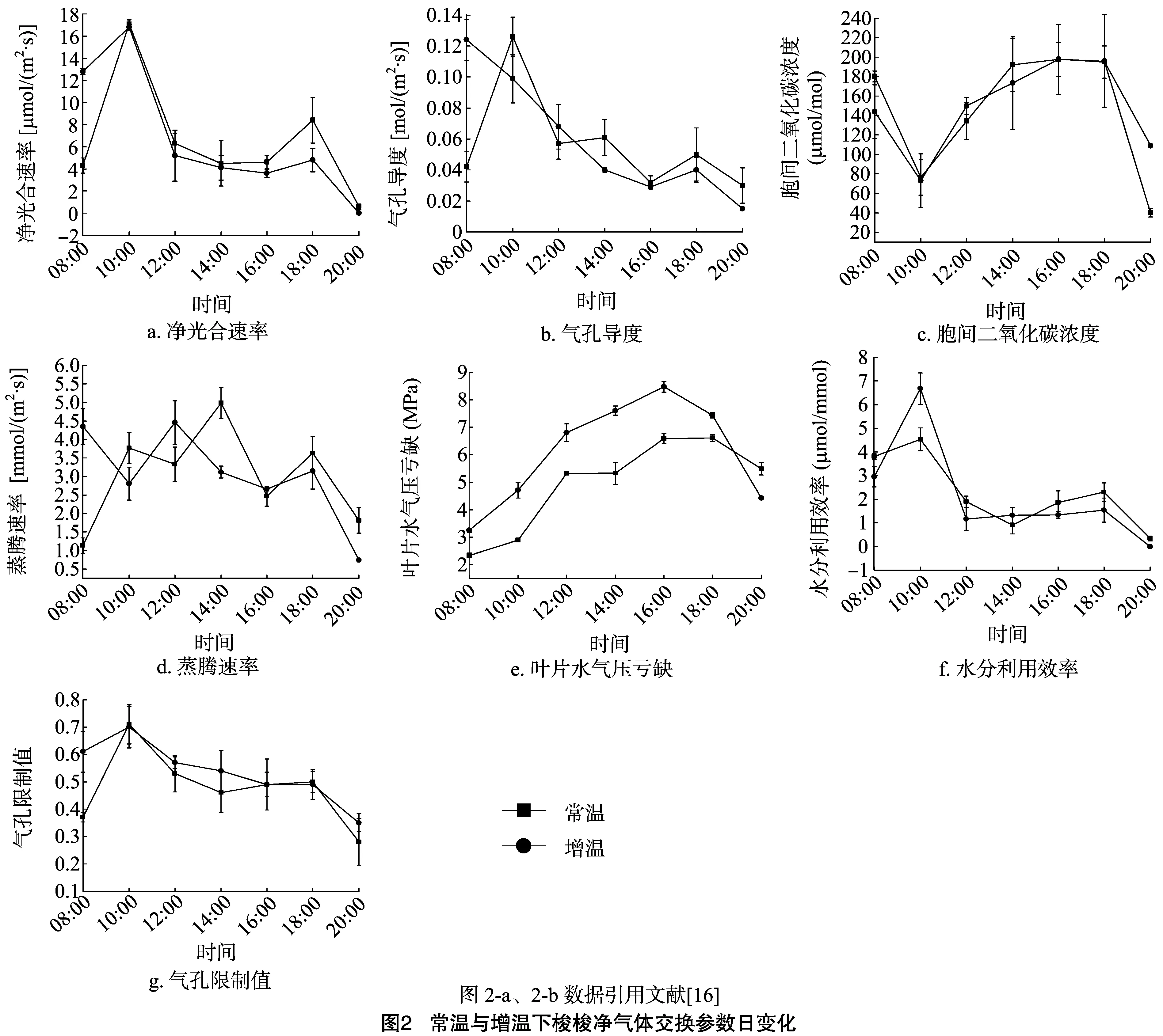
图2-b表明,在08:00—20:00常温与增温下梭梭Gs的变化趋势则表现出截然不同的情况。常温下梭梭Gs在 08:00—12:00变化剧烈,10:00达到峰值0.126 mol/(m2·s),之后下降至16:00达到最低值,为0.032 μmol/(m2·s),其间14:00时有小幅度的回升波动,16:00后才又逐渐回升,直至18:00左右,最后方才呈下降趋势,从而使其日变化曲线呈“多峰”型。而增温下梭梭Gs则在08:00—16:00期间呈持续下降变化,表明该环境下的温度一开始就已对梭梭气孔运动产生了一定的胁迫效应。最后,通过图2-b可看出增温下梭梭Gs也整体低于常温组,但不显著(P>0.05)。
图2-c表明,梭梭Ci的日变化趋势与其Pn日变化的趋势相反。在08:00—16:00梭梭Ci均变化剧烈,Ci均于 10:00 达到最小值,其值分别为76.6、73.1 μmol/mol,又在10:00—16:00迅速上升,之后呈持续下降趋势。其主要原因在于太阳辐射的增强和环境温度的逐渐升高,使得光合作用酶的活性增强,从而加快了光合作用速率,导致Ci降低,但是由于酶活性受温度的影响,导致在环境温度高于酶最大活性温度范围时,光合速率减慢,气孔逐渐关闭,Ci消耗达到最小值。与此同时,也使其呼吸作用加快,CO2释放量增加,最终在一定时间内表现出Ci的升高。说明随着温度的升高,增温下梭梭Ci逐渐高于常温下梭梭Ci,但差异性不显著(P>0.05)。
2种生境下梭梭Tr的日变化趋势也均截然不同,常温下梭梭Tr的日变化趋势却与Gs的变化趋势相似(图2-d、图 2-b),即在10:00、14:00、18:00相继出现3个峰值,其大小分别为3.77、4.99、3.63 mmol/(m2·s);同时也在12:00、16:00相继出现2个峰谷值,分别为3.33、2.47 mmol/(m2·s)。然而增温下梭梭Tr的变化趋势则呈现仅在14:00之后与常温下梭梭Tr的变化趋势相同,其余时间段则与之相反(图2-d)。
在08:00—20:00期间2种生境下梭梭VPD的日变化均呈“单峰”型曲线(图2-e),且最小值的出现均在08:00,其大小分别为2.34、3.25 MPa;最大峰值则出现在16:00,其值分别为6.59、8.47 MPa。在整个过程中,增温下梭梭VPD显著(P<0.05)高于常温下梭梭VPD。其主要原因可能在于随着温度和光辐射的增强,植物体为避免被高温与过强的太阳辐射灼伤,其幼嫩同化枝及其细胞组织结构就必须尽可能通过增加蒸腾作用使水分挥发带走大量热能,从而来降低体表温度,避免温度过高带来的伤害;但梭梭的生长环境本身又是一个白天温度极高加之降雨不均且蒸发量极大的沙漠或荒漠环境,最终导致水分缺乏,使其吸水速率低于蒸腾失水速率,而出现VPD升高、同化枝水势的降低及气孔的关闭。
WUE反映了植被光合生产过程与耗水特性之间的关系。经计算,常温和增温下梭梭WUE的日变化总体趋势相似,均表现出明显的“双峰”型(图2-f)。在08:00—12:00梭梭WUE变化剧烈,均于10:00达到第1个峰值,其大小分别为4.54、6.68 μmol/mmol,之后分别迅速下降,分别在14:00、12:00达最低值,其值大小分别为1.90、1.16 μmol/mmol,最后逐渐回升,均于18:00出现第2个峰值。总体可以看出,在增温下梭梭WUE明显高于常温组。Ls反映了由于气孔导度的降低,导致进入气孔的CO2的减少程度[17]。经计算,2种生境下梭梭Ls的日变化趋势并不完全同步(图2-g),常温下梭梭表现出较明显的“双峰”型,2个峰值分别出现于 10:00、18:00;而增温下梭梭则呈现出明显的“单峰”型,其峰值出现于10:00。
2.3 不同生境梭梭光合生理分析
植物Pn和Tr不仅受到内在生理因子的调节,同时还受到环境因子的影响[17-19]。从表2的分析结果可看出,2种生境下梭梭Pn与Gs之间均呈极显著正相关,而其Tr则仅在常温下与PAR呈显著正相关。其中,常温下梭梭Pn与Ta及VPD均呈负相关,与PAR、RH和Gs则呈正相关。而其Tr则仅与RH呈负相关;环境因子中对常温下梭梭Pn和Tr的影响依次分别为Gs>RH>VPD>PAR>Ta、PAR>Gs>Ta>VPD>RH。增温下梭梭Pn与Ta和VPD也呈负相关,与PAR、RH及Gs则呈正相关,而其Tr与环境因子之间的关系则均呈正相关;影响增温下梭梭Pn和Tr的环境因子大小依次分别为Gs>RH>Ta>VPD>PAR、Gs>PAR>RH>VPD>Ta。
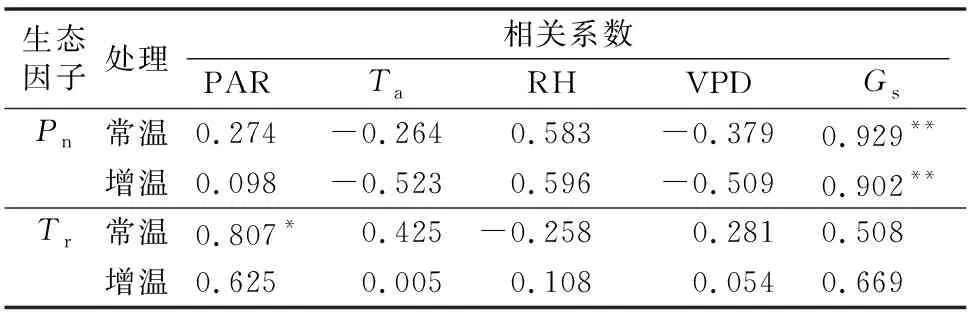
表2 梭梭同化枝Pn、Tr与生理生态因子的相关系数
注:*、**分别表示在0.05、0.01水平的双尾检验中差异显著。
3 讨论
植物光合作用日变化是在一定天气条件下,各种生理生态因子综合效应的最终反应[17-18]。在整个观测期内,梭梭2种生境中PAR和Ta的日变化均为“单峰”曲线,RH则为“W”形,Ca为“S”形。一般情况下,植物光合作用日变化均有规律可循,变化曲线呈“单峰”型或“双峰”型[17-20]。2种生境下梭梭同化枝Pn在试验观测期间的日变化曲线均呈“双峰”型,且峰值出现的时间一致。随着PAR的逐渐增强,Ta升高而RH减小,但由于生长室内的Ta上升较快且最初RH较大,从而导致梭梭Pn在前2 h内显著大于常温下梭梭Pn且均呈上升趋势;之后PAR和Ta继续上升,使得RH进一步减小,进而造成水分亏缺而迫使Pn降低,但对10:00的Ta来说,则为该环境下梭梭叶片光合作用的最适温度,在低于此温时,升温可促进梭梭的光合作用,反之,则产生抑制作用。这与胡将男等研究高温对杨梅叶片气体交换的影响结果[21]相似。同时,本研究结果也进一步表明了增温下梭梭受高温胁迫较常温更为严重。不同生境下梭梭Gs值日变化趋势则完全不同。相对Pn而言,仅常温下梭梭Gs值日变化趋势与其相似,而增温下梭梭Gs日变化则在整个过程中均呈整体持续下降趋势。
Tr是衡量植物水分状况的有效指标,同时WUE的大小也可反映植物对干旱环境适应能力的强弱[17],而高的VPD则可使植物叶片的光合速率下降[22]。在本研究中,利用生长室模拟增温效应,得到梭梭同化枝VPD明显增加,同时伴随着Pn的下降,其主要原因是高温对光合速率的影响是通过梭梭同化枝VPD的上升来实现的。
光合作用的降低一般可归于2个原因:气孔因素和非气孔因素,但判断其降低的主要原因是气孔因素还是非气孔因素的2个可靠判据则是Ci和Ls的变化。若Ci降低而Ls升高,则表明气孔导度降低是主要原因;反之,则表明主要原因是非气孔因素,即归因于叶肉细胞羧化能力的降低[19,23]。尽管本试验2种生境下梭梭Pn日变化均随着光合有效辐射的增加呈上升趋势,但当PAR和Ta的进一步升高,Pn则开始下降,且出现下降的时间同步,均从10:00开始;在此过程中Ci升高而Ls降低,表明2种生境下梭梭午间Pn下降均主要是由非气孔因素造成的。但从14:00之后则主要是由于气孔限制所致。植物这种所谓的“午休”现象,也是其适应中午高温高光强而免受伤害的一种普遍适应策略,也是提高WUE的途径之一[17-18]。此结果与赵长明等对梭梭同化枝Pn的研究结论[24]存在一定差异。
植物生长是对于各环境因子综合作用的反映[18,25-56],且已有不少研究发现,即使是同一环境因子,在不同生境下对同物种所发挥的主导作用也是不尽相同的[27-28]。如本研究中常温下梭梭Tr与RH呈负相关性,而在增温下则呈正相关性;这表明增温下梭梭的生理需水已出现亏缺现象,进而导致梭梭同化枝气孔的关闭和Tr的降低,同时也降低了叶肉细胞的羧化能力,致使Ci的升高。此外,在2种生境下RH是对梭梭Pn起最大主导作用的环境因子,而对梭梭Tr起最大主导作用的环境因子则为PAR,这些都充分表明了各环境因子相互作用时的复杂性。
4 结论
通过对2种生境下梭梭气体交换特征及相关环境因子的同期观测与对比分析得到:增温生境下梭梭受高温胁迫程度较常温严重,造成其适应干旱环境的能力减弱。因此,若未来全球气温进一步变暖,则将对梭梭种的生存适应能力与分布造成一定程度上的影响。在未来全球性气候进一步变暖的环境下,强辐射、高温及其所导致的水分缺乏等环境因子虽都将对梭梭Pn产生一定程度的影响,但水分缺乏对其Pn的影响将更加显著。
猜你喜欢
钻井液与完井液(2022年4期)2022-10-26 06:39:38
哈哈画报(2022年4期)2022-04-19 11:11:54
哈尔滨轴承(2021年1期)2021-07-21 05:43:14
落叶果树(2021年6期)2021-02-12 01:29:10
科学大众(2020年17期)2020-10-27 02:48:48
中学生天地(B版)(2019年4期)2019-05-08 03:22:54
电镀与环保(2017年3期)2017-06-23 08:24:51
体育科技(2016年2期)2016-02-28 17:06:07
绿色中国(2015年6期)2015-07-03 01:13:10
西华师范大学学报(自然科学版)(2015年3期)2015-02-27 15:31:17
