
Effects of N:P ratio of Artemisia ordosica on growth influenced by soil calcium carbonate
2018-09-07 06:46:32YuTingLiangXingDongHeJianTanGuoHongJuanJing
YuTing Liang, XingDong He , JianTan Guo, HongJuan Jing
College of Life Sciences, Nankai University, Tianjin 300071, China
ABSTRACT Soil calcium carbonate (CaCO3) has a strong solid phosphorus effect, and high content of CaCO3 can significantly reduce the effectiveness of soil phosphorus. To reveal the limiting effect of soil CaCO3 on the growth of plants on sand land and its mechanism of plant physiology, we performed pot experiments with a two-factor randomized block design and a three-factor orthogonal design for different soil CaCO3 content treatments using Artemisia ordosica seedlings. In the experiments, we surveyed plant height, aboveground biomass, root length and root weight and analyzed N, P concentrations and RNA content of the seedlings, and discussed the relationships between relative growth rate (RGR) of the seedlings and N:P ratio as well as RNA. Results show that, the RGRs of plant height and above-ground biomass of the seedlings decreased significantly with the increase of soil CaCO3 content, and those for root length and root weight decreased. The RGRs of plant height and above-ground biomass of the seedlings were significantly negatively correlated with leaf N:P ratios, but significantly positively correlated with leaf RNA content and leaf P concentrations. It can be seen that soil CaCO3 is a stress factor for the growth of A. ordosica seedlings, and the growth response of the seedlings under the influence of soil CaCO3 is in line with the Growth Rate Hypothesis.
Keywords: Artemisia ordosica; relative growth rate; N:P ratio; RNA; soil CaCO3
1 Introduction
Plant nitrogen to phosphorus ratio (N:P ratio) is the link between different research scales from molecular, cell, individual, population to the community and ecosystem (Elser et al., 2000; Zeng and Chen,2005; He and Han, 2010). The plant N:P ratio can affect plant growth rate (Elser et al., 2000; He et al.,2016), thus influencing the phenotypic plasticity of plants (Pigliucci, 2001). The plant N:P ratio rather than phenotypic plasticity affects the dynamics of plant population or community (Reader et al., 1993;Schlichting and Smith, 2002). The variations in plant N:P ratio depend on elements such as soil nutrients,moisture, salts and air temperature, hence control changes in plant growth rate and affect plant population and community.
In arid and semi-arid regions, land desertification is very serious (Li et al., 2016; Luo et al., 2016; Yu,et al., 2017). During the process of sand dune fixation, the soil calcium carbonate (CaCO3) content increases significantly (Xiao et al., 2003; Wang et al.,2007). The increasing CaCO3in sand soil can limit the absorption of P and produce adverse effects on plants (Tunesi et al., 1999; Arnesen et al., 2007), thus the effect of soil CaCO3on the growth of plants on sand land needs to be explored. On the other hand,our previous research results show that the ratio of soil CaCO3to available P can significantly affect the density of Artemisia ordosica in arid and semi-arid areas (Zhao et al., 2012), which means that soil CaCO3can affect the growth rate of A. ordosica.Thus, the intrinsic mechanism of why soil CaCO3can affect the growth rate of A. ordosica needs to be further revealed.
In this study, we chose A. ordosica (an important dominant species of plant community on sandy soils of arid and semiarid areas) as the research object, performed pot experiments of a two-factor randomized block design and a three-factor orthogonal design for the different soil CaCO3content treatments. We also analyzed the influences of soil CaCO3, leaf N:P ratio,leaf P and RNA on the relative growth rate (RGR) of A. ordosica seedlings. Our aims are: (1) to reveal how soil CaCO3affects the growth of A. ordosica; (2) to illustrate the physiological basis of soil CaCO3affecting the growth of A. ordosica.
2 Materials and methods
2.1 Pot experiment setup
Pot experiments were performed in a greenhouse at the College of Life Sciences, Nankai University,using A. ordosica. Seeds of A. ordosica and the aeolian sand soil in the pots were collected from the Ningxia Habahu National Nature Reserve.
In this study, we designed a two-factor randomized block experiment and a three-factor orthogonal experiment. The two-factor experimental design (4×4)was composed of four soil CaCO3contents (10, 50,75 and 100 g/kg chemically pure) and four watering regimes (240, 260, 280 and 300 mL distilled water per week). The orthogonal experimental design (L16(45))was with 3 factors and 4 levels, soil CaCO3contents were 1, 25, 50 and 100 g/kg, available P in soil were 0.1, 3, 6, 12 mg/kg dry soil (KH2PO4was converted to P, i.e., 136.09 mg KH2PO4contains 30.97 mg P,corresponding KH2PO4was mixed with soil, respectively), and watering regimes were 240, 260, 280 and 300 mL per week. Each treatment was replicated three times in both experiments.
A. ordosica seeds were germinated in a potting mix in the greenhouse. The substrates in the mix consisted of perlite and turf soil (1:1). Three or four seeds per hole were sowed on the surface, and covered with thin paper films, then watered. After germination, the films were removed, and the mix was kept moist until they were transferred to the pots.
According to experimental designs, we mixed the aeolian sand, available P and CaCO3and put them into PVC pots (195 mm tall and 165 mm diameter) with corresponding numbers. Each pot was filled with 2 kg of the mixture to a depth of 12 cm. A. ordosica seedlings were divided into large, medium and small groups by growth conditions, and the seedlings in each group were adjusted with the same growth status. After transferred to the pot, the root neck part of A. ordosica seedling in every pot was marked with red paint for the determination of plant height after pot experiment. At the same time, the residual 102 seedlings were dug out and cleaned, to measure plant height, root length, shoot biomass and root biomass to construct regression equations.
After transplanted, seedlings were watered according to experimental designs every week. During the experiment, the average temperature in the greenhouse was around 25 °C during the day and 18 °C at night. After 54 days for the experiment, the seedlings were dug out and cleaned. Plant height, root length,shoot and root biomass of each seedling were measured and weighed. The plant samples were then placed in marked sample bags and stored in a -80 °C refrigerator.
2.2 Calculation of plant growth rate
The residual 102 A. ordosica seedlings were utilized to perform regression analysis, using plant height as the independent variable and shoot biomass, root length and root biomass as dependent variable, respectively. The analyses of three regression equations were all significant, so they were all valid. Then, we used the plant height of every seedling before the experiment to calculate responsive aboveground biomass, root length and root biomass through the three regression equations (Li et al., 2010; Wu et al., 2010).
Then, we calculated relative growth rate (RGR) of every index of A. ordosica seedlings utilizing the measured values and calculated values before the experiment as well as the measured values after the experiment according to the formula of (Yu et al., 2012).

where μ is RGR; M0is plant height, shoot biomass,root length or root biomass before the experiment; Mtfor after the experiment; and t for growth time (54 days).
2.3 Sample analyses
Plant N was determined using the Kjeldah method (Bao, 2000), while plant P was determined using the H2SO4-H2O2-Mo-Sb-Vc-colormetry method (Bao,2000). RNA was extracted using improved CTAB method, and the extract was measured for the content of RNA through Thermo Scientific NanoDrop2000(Xiao et al., 2010).
Because A. ordosica seedlings were small in the experiment and biomass was insufficient, we determined plant N and P using the seedlings in the twofactor experiment and RNA using seedlings in the three-factor orthogonal experiment.
2.4 Statistical analyses
Duncan test for significance test of difference was performed among plant height, or aboveground biomass, root length and root biomass of the seedlings under different soil CaCO3contents in both experiments using SPSS 19.0 software. Simple linear regression relations were performed between RGRs of plant height or aboveground biomass and the N:P ratios, and between RGRs of plant height or aboveground biomass and RNA contents through SPSS 19.0. Figures were made by Excel 2010.
3 Results
3.1 Influence of soil CaCO3 content on the growth rate of A. ordosica seedlings
As presented in Figure 1, the results of the twofactor and the orthogonal experiments indicated that with an increase of soil CaCO3contents, the RGRs of plant height and aboveground biomass of A. ordosica seedlings were significantly reduced, while the RGRs of root length and root biomass had no significant change. In the two-factor experiment, the root/shoot ratios (0.17, 0.19, 0.24, 0.38) increased with the increase of soil CaCO3contents, i.e., it caused redistribution of biomass. The decrease rate of RGRs of the aboveground biomass was higher than that of root biomass. Compared with the control group, the RGR of above-ground biomass decreased 89% under 100 g/kg soil CaCO3. The RGRs of root length were negative, and the negative values were increasing.Thus, soil CaCO3limited the growth of the seedlings.In the two-factor and orthogonal experiments, the water factor and its interactions with soil CaCO3had no significant influence on the RGRs of A. ordosica seedlings.
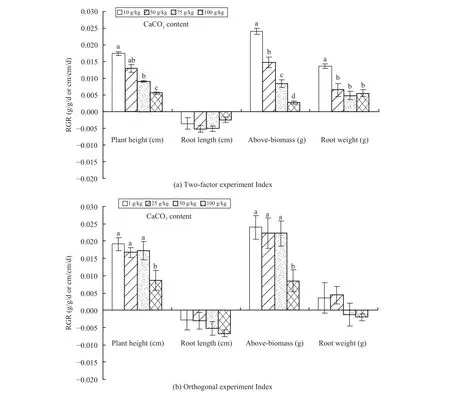
Figure 1 Relative growth rate of Artemisia ordosica seedlings under different soil CaCO3 contents
3.2 Influence of leaf N:P ratio of A. ordosica seedlings on growth
In the two-factor experiment, when soil CaCO3contents were at levels of control group, mild, moderate and severe, the average concentrations of leaf N in A. ordosica seedlings were 6.071, 5.596, 5.466 and 4.935 g/kg, respectively, but there was no significant difference among them. Alternatively, the average concentrations of leaf P were 1.251, 1.196, 0.626 and 0.303 g/kg, respectively, in which significant difference was observed among treatments (p<0.001). The average N:P ratios were 4.874, 6.217, 9.523 and 10.662, respectively, in which significant difference was found among treatments (p<0.05). Regression results show that with an increase of leaf N:P ratios of A. ordosica seedlings, both RGRs of plant height and above-ground biomass presented significantly negative power correlations (Figure 2), and the increase of plant height and aboveground biomass significantly slowed down. With the increase of leaf P of A. ordosica seedlings increasing, both RGRs of plant height and of above-ground biomass took on significantly positive straight line correlations (Figure 3).
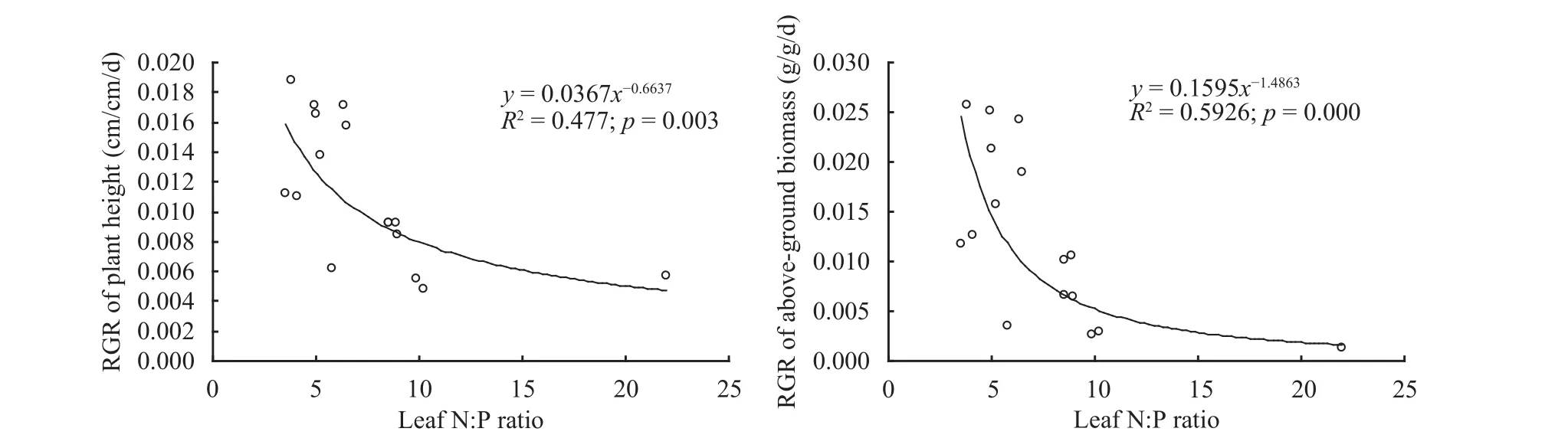
Figure 2 Influences of leaf N:P ratio of A. ordosica seedlings on growth affected by soil CaCO3
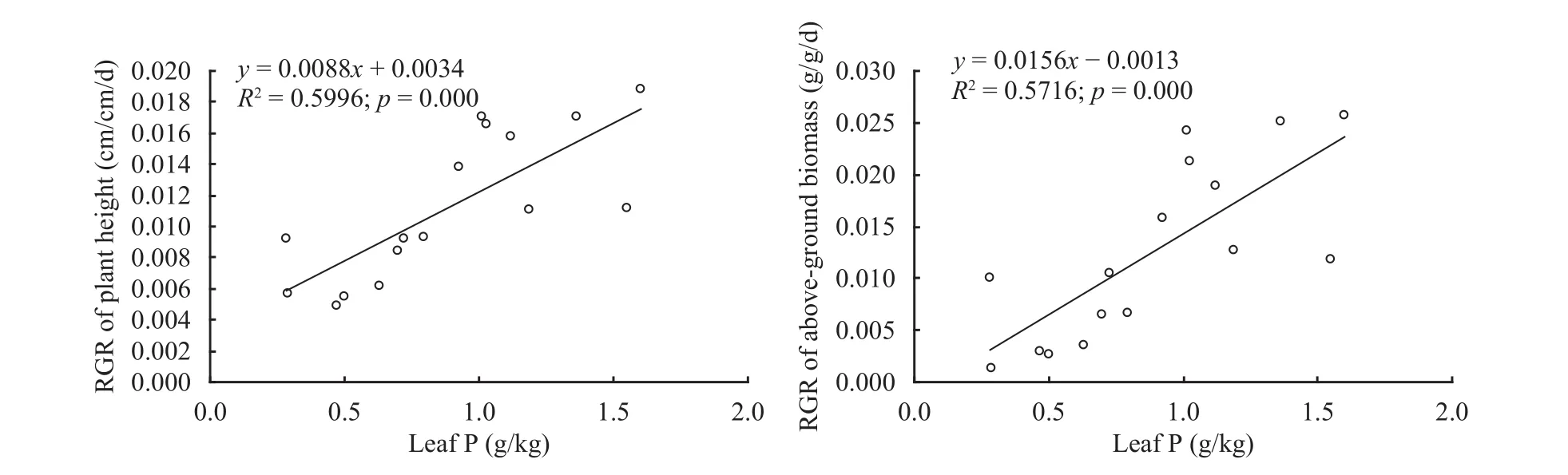
Figure 3 Influences of leaf P concentration of A. ordosica seedlings on growth affected by soil CaCO3
3.3 Influence of leaf RNA on growth of A.ordosica seedlings
In the orthogonal experiment, with the soil CaCO3contents increasing, no significant difference was observed in RNA contents of A. ordosica seedlings.However, compared with the control group, with an increase of soil CaCO3contents, RNA contents of the seedlings show a trend of decreasing firstly and then increasing. The results of regression indicated that with the increase of leaf RNA of A. ordosica seedlings, both RGR of plant height and shoot biomass presented significantly positive power correlations(Figure 4), and the growth of plant height and aboveground biomass rose significantly.
4 Discussion
Pedogenic calcium carbonate is of great significance to plant nutrition, carbon budget, paleoclimatology, paleoecology, and soil science itself (Monger,2002). A large amount of active liquid calcium ions in soil directly leads to the formation of Ca-P precipitate phases (Tunesi et al., 1999; Carreira et al., 2006),such as dicalcium phosphate and octacalcium phosphate (Ma et al., 2009), resulting in phosphorous restriction. Moreover, the higher levels of soil CaCO3increase soil pH, which further promotes the precipitation of Ca ions and P (Tunesi et al., 1999; Zhao et al., 2007), thereby reducing the availability of P in soil (Arnesen et al., 2007). In the present study, for both the two-factor and orthogonal experiments, with an increase of CaCO3contents in sandy soil, the RGRs of plant height, above-ground biomass, root length and root weight of A. ordosica seedlings were significantly reduced. The RGRs of root length show a "negative growth" phenomenon, and the negative growth rate show an increasing trend, the soil CaCO3curtailed the root length of A. ordosica seedlings. Obviously, the more CaCO3content in the soil, the more severe effect on the growth of A. ordosica seedlings,indicating that soil CaCO3was a stress factor for the growth of A. ordosica seedlings. On the one hand, this was due to the temporary soil drought caused by the existence of soil CaCO3in sandy soil (Jin et al.,2005); the effective reduction of soil moisture allowed the survival of A. ordosica seedlings in an unfavorable environment, that is, an adversity. On the other hand, the increase of CaCO3in soil reduced the concentrations of available P in soil and thus affected the seedlings. Our results found that leaf P of A. ordosica seedlings reduced significantly with increasing soil CaCO3contents. Lajtha and Schlesinger(1988) studied Larrea tridentata and Parthenium incanum and found that for L. tridentata seedlings, soil CaCO3directly inhibits P uptake or the availability of P in the roots. Also, with a gradient rising of CaCO3,plant N:P ratios increased, and the rate of P absorption decreased significantly. But the response of P. incanum seedlings to soil CaCO3mainly reflects the decrease of growth, the P concentrations in all parts of the plant decrease with an increase of soil CaCO3, and the difference in the root is greater than that in the aboveground. Thus, the increase of soil CaCO3can reduce the absorption and utilization of P by plants,i.e., the decrease of available P content in soil limits the normal growth of plants.
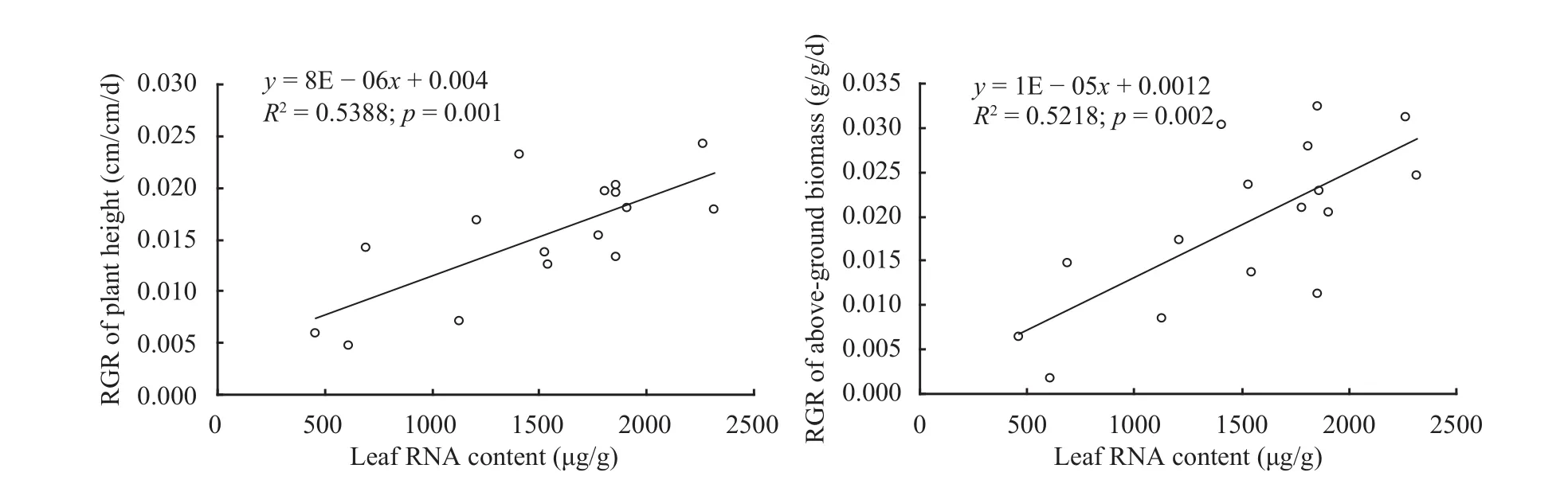
Figure 4 Influences of leaf RNA in A. ordosica seedlings on growth affected by soil CaCO3
The Growth Rate Hypothesis of Elser et al. (2000)indicates that organisms have smaller N:P ratios for rapid growth. Ågren (2004) found that when the RGR of plants is relatively low, N:P ratios in the body gradually increases up to the maximum value, then the N:P ratios gradually decreases with the increase of the RGRs. In the present study, under the existence of soil CaCO3, the RGRs of plant height and aboveground biomass of A. ordosica seedlings were significantly reduced with increasing leaf N:P ratios, i.e.,the larger the N:P ratio in the A. ordosica seedlings,the slower the growth of A. ordosica seedlings, which was in line with Elser's Growth Rate Hypothesis. This study found that N:P ratios of A. ordosica seedlings was linearly positively correlated with soil CaCO3content. With the increase of soil CaCO3contents, the RGRs of A. ordosica seedlings decreased but the N:P ratios increased. When the soil CaCO3contents increased from the control group (10 g/kg) to the light(50 g/kg) and moderate (75 g/kg), the P concentrations in A. ordosica seedlings decreased with the increase of soil CaCO3content, which is consistent with the changes in the RGRs of A. ordosica seedlings.This is because A. ordosica had a greater demand for P during its growth, the P effectiveness was limited as the soil CaCO3content increased, and thus P became a limiting factor for the growth of A. ordosica seedlings. The P reduction in A. ordosica seedlings resulted in an increase in N:P ratios, therefore, the N:P ratios in A.ordosica seedlings increased and the growth of the seedlings became slower.
The Growth Rate Hypothesis of Elser et al. (2000)shows that, to grow rapidly, organisms invest more in P-rich ribosomes and rRNA to facilitate the rapid synthesis of proteins, that is, more RNA in the body facilitates the synthesis of more proteins, which allows individuals to have greater growth rates. In the present study, the RGRs of plant height and aboveground increased significantly with an increase of RNA contents and leaf P concentrations in A. ordosica seedlings, which is consistent with the Growth Rate Hypothesis. Theoretical analysis shows that the synthesis of P-rich ribosomal RNA (rRNA) in A. ordosica seedlings increased, and the amount of protein synthesis under the guidance of rRNA increased. The large quantity of proteins was synthesized to repair and up-date tissue and new tissue proliferation, the synthesis rate was positively correlated with the plant growth rate. Therefore, plant height and aboveground biomass of A. ordosica seedlings grew faster with the increase of RNA contents.
Our previous research shows that when the magnitude of soil CaCO3:available P ratio is the same and less than 2.5, the density of A. ordosica in the community gradually increases in semi-arid areas and decreases gradually in arid areas with an increase of soil CaCO3:available P ratios. In semi-arid regions, when the magnitude of soil CaCO3:available P ratio is unchanged and the increase amplitude of soil CaCO3is greater than that of available P, the density of A. ordosica increases gradually with the increase of soil CaCO3:available P ratios. In arid regions, when the magnitude of soil CaCO3:available P ratio increases as well as soil CaCO3content increases significantly and available P significantly decreases, the density of A. ordosica increases gradually with the increase of soil CaCO3:available P ratios (Zhao et al., 2012). The results of this study suggests that the soil CaCO3:available P ratio could significantly influence the density of A. ordosica in arid and semi-arid regions. At the same time, the results of this study also verified the Growth Rate Hypothesis of Elser et al.(2000).
5 Conclusions
Soil CaCO3can limit the growth of A. ordosica seedlings. Under the existence of soil CaCO3, changes of N and P concentrations in A. ordosica seedlings lead to changes of the N:P ratios and the RNA and protein contents in the body, which in turn regulates the redistribution of their aboveground and underground biomass. Thus, the change of RGRs of A. ordosica seedlings is the basis for its survival and resistance to stress. With the soil CaCO3contents increasing, the RGRs of plant height, above-ground biomass,root length and root weight of A. ordosica seedlings all reduce. The RGRs of plant height and aboveground biomass of A. ordosica seedlings are negatively correlated with the leaf N:P ratio and positively correlated with the leaf P concentration and leaf RNA content.
Acknowledgment:
This research was supported by the National Key Research and Development Program of China(2016YFC0500706).
 Sciences in Cold and Arid Regions2018年4期
Sciences in Cold and Arid Regions2018年4期
- Sciences in Cold and Arid Regions的其它文章
- International Workshop on Cryospheric Changes and Their Regional& Global Impacts Successfully Held in Gansu Province
- Cultivated-land change in Mu Us Sandy Land of China before and after the first-stage grain-for-green policy
- Comparisons of plant calcium fraction between two different vegetation zones in semi-arid region
- Seasonal dynamics of N:P ratio stoichiometry and Ca fraction for four dominant plants in the Alxa Desert
- Change in summer daily precipitation and its relation with air temperature in Northwest China during 1957-2016
- Analysis of water vapour flux between alpine wetlands underlying surface and atmosphere in the source region of the Yellow River