
穿叶眼子菜茎叶结构及其通气组织发育解剖学研究
2018-09-05 06:18:06倪细炉
西北植物学报 2018年7期
王 晨,李 龙,倪细炉*,李 健
(1 宁夏大学 生命科学学院,银川 750021;2 宁夏林业研究院 种苗生物工程国家重点实验室,银川 750004)
通气组织(aerenchyma)是由位于内外皮层之间的一类特异的皮层细胞通过细胞分裂或溶解后而形成的腔隙,因形成方式的不同可分为溶生性(lysigenous)与裂生性(schizogenous)2种[1]。溶生性通气组织的形成过程已经有大量的文献报道,在水稻的叶鞘中,通气组织通过细胞死亡形成于维管束之间的薄壁细胞[2]。水稻根部通过皮层细胞降解而形成在横切面上呈放射状的通气组织[3],且缺氧处理会使其根部通气组织的形成起点更靠近根尖[4]。Typhaangustifolia叶通气组织形成前期开始出现部分晶状体细胞,随后晶状体细胞及周围细胞降解形成腔隙,腔隙的形成由靠近上下表皮处开始逐渐向叶中央扩展,最终腔隙间仅存在3~5层细胞及一排维管束[5]。在Trapapseudoincisa叶柄通气组织的形成过程中,于纵切面的腔隙内观察到了质体降解、液泡破坏、次级液泡出现的异常细胞,通过凝胶电泳及TUNEL检测也证明了其叶柄膨大涉及PCD过程[6]。对于一种沼泽植物Sagittarialancifolia的研究发现,其根内溶生性通气组织形成于分生组织并扩展到成熟区[7]。即使非水生或湿地植物在缺氧胁迫下也会产生溶生性通气组织以维持生长[8],如小麦根在水淹条件下皮层细胞死亡,形成发达的通气组织[9],在玉米[10]及向日葵[11]等作物中同样发现溶生性通气组织。裂生性通气组织的报道较少,且常见于湿地植物[12]。前面提到S.lancifolia的根部通气组织为溶生性,而其叶柄中仅通过细胞的分裂形成了贯穿于叶脉的腔隙[7]。Purnobasuki等对Sonneratiaalba的营养根研究发现,距根尖0~500 μm以内的皮层细胞排列紧密,基本无细胞间隙的形成;距根尖超过500 μm之后,皮层细胞开始相互分离并在根纵向上形成间隙,这标志着由细胞间隙的扩大而导致的裂生性通气组织开始形成[13]。除了湿地植物,Rumex在缺氧条件下也会加速新生跟生长,且新根内孔隙率更高[14]。通气组织的形成过程有时同时涉及裂生与溶生[15],这种形成方式为裂溶性通气组织。水稻根在靠近根与根冠结合处的中部皮层出现裂生性细胞间隙,而根冠、中柱与外皮层没有,但在距根尖3~4 cm根段的皮层出现了溶生性通气组织[16]。Bartoli等[17]通过对Egeriadensa茎通气组织的研究发现其早期在茎尖近端的皮层细胞间形成裂生腔隙,且在腔隙的膨大过程中检测到了细胞凋亡,最终形成蜂窝状的成熟通气组织。除了形成方式的不同,通气组织在形状上也有差异;通过对110种水生及湿地植物通气组织的研究[18],发现茎中存在5种形状的通气组织:蜂窝状、车轮状、中空状、叶状、溶生气道,其中蜂窝状与车轮状在形态上有一定相似性,水生植物大多数器官中发育出比湿地植物更发达的通气组织,且通气组织的形成模式在属水平上是稳定的。水生和湿地植物的根茎内均形成通气组织,从而在水淹情况下能够向根部运输氧气[19],保证其代谢需要。
穿叶眼子菜(Potamogetonperfoliatus)为眼子菜科眼子菜属中分布广泛的一种多年生沉水植物[20],其茎叶发达,节处生有须根。对于眼子菜属植物的通气组织已有部分报道,其根、茎、叶内均形成通气组织,Bona等[21]对Potamogetonpolygonus根部通气组织的研究,发现其先是形成小的裂生腔隙,随后的扩大过程涉及细胞坍缩,最终形成成熟的车轮状通气组织。丝叶眼子菜茎的横切面观察到了2~3轮发达的车轮状通气组织[22],且其花梗和叶内同样形成较发达的蜂窝状通气组织。另一种浮叶眼子菜Potamogetondistinctus的根、茎皮层薄壁细胞内形成网状通气组织,且茎内存在4束维管束,其叶由上下表皮、栅栏组织及位于栅栏组织下方的薄壁细胞组成,薄壁细胞内形成网状通气组织[23],叶脉分布于网状通气组织中。马毓泉等发现眼子菜属植物的叶片由上下表皮及1~2层叶肉细胞组成,整个叶片内充满气腔或仅主脉周围的叶肉内具气腔[20]。
综上所述,目前对通气组织的形成模式及形态特点已有较多的研究,但对于眼子菜通气组织的研究以及通气组织在时间、空间上的形成规律及特点研究较少。因此,本实验以穿叶眼子菜为材料,以其茎和叶为对象,通过连续切片在显微结构水平上研究通气组织的发育过程,以及通气组织发生的时空变化,以期为水生植物通气组织发育调控以及水生植物适应水环境的生存机制提供依据。
1 材料和方法
1.1 材料的培养
实验材料为穿叶眼子菜,通过植物组织培养的方法培养其无菌苗。培养条件为MS基础培养基及无菌水,室温光照12 h/d。培养方法为:将消毒处理过的穿叶眼子菜茎段或转接的无菌茎段接入培养基内,待转接茎段生根后向培养基内加入适量的灭菌水,以淹没植物顶端为宜;当新生茎长度达到5 cm左右即可取材,穿叶眼子菜的茎由节和节间两部分组成,其中,不同生长时期的茎节间处的结构不同,可分为实心期、形成期、膨大期和成熟期等4个时期,而不同生长时期的茎节处的结构相同。
1.2 石蜡切片
1.2.1茎横切石蜡切片的制作取长度为1.5 cm左右的新生茎尖,除去叶片后迅速固定于FAA固定液24 h以上;乙醇梯度脱水(70%、85%、95%、100%);二甲苯梯度透明(50%、100%),透明时可加少许番红对材料染色,便于后面操作步骤对材料的辨别;梯度浸蜡(50%、100%);纯蜡包埋。
用石蜡切片机(Leica RM2125 RTS)从茎尖开始进行连续横切,切片厚度为10 μm。粘片时每张载玻片粘4条蜡带并按一定顺序排好,连续切1 cm的茎尖为25张片子。将粘好的片子于烘片机(Leica HI1220)上38 ℃烘干后放40 ℃烘箱内过夜,然后染片。
染片时在染色缸内进行,首先是复水,梯度二甲苯(100%、50%);梯度酒精(100%、95%、85%、70%);番红染色(过夜);梯度酒精脱水(70%、85%、95%);固绿染色(数秒);100%酒精;梯度二甲苯(50%、100%);封片,显微镜观察。
由于所做的是从茎尖长度为1 cm的连续切片,所以最终可以观察到各个节处的横切结构,以及节间通气组织不同发育阶段的横切结构。
1.2.2茎纵切石蜡切片的制作取长度为1.5 cm左右的新生茎尖,除去叶片后迅速固定于FAA固定液24 h以上;乙醇梯度脱水(70%、85%、95%、100%);二甲苯梯度透明(50%、100%);梯度浸蜡(50%、100%);纯蜡包埋。
用石蜡切片机(LEICA RM2125 RTS)对包埋好的材料进行纵切,切片厚度为10 μm。粘片、烘片,放40 ℃烘箱内过夜;之后的染片步骤同上,最后封片,观察。
通过茎尖的纵切能够整体观察到各个节处的纵切结构,以及节间通气组织不同发育阶段的纵切结构。
1.2.3叶横切石蜡切片的制作取长度约0.5 cm左右的茎尖及距茎尖约5 cm处的成熟叶片,分别按照以上方法固定、包埋。然后将材料连续横切,厚度为10 μm。从茎尖的横切中选取具有不同发育程度叶片的片子进行染色,成熟叶片的横切直接染色,染色方法同上,最后观察。
通过茎尖叶以及成熟叶片的横切能够观察到叶通气组织的不同发育阶段。
1.3 茎横切面通气组织百分率(孔隙度)的测定
选取所做茎横切切片中的节处横切图片及节间通气组织不同发育时期显微图片各6张,用软件Motic image plus 2.0 ML测量每张横切图片中腔隙的总面积S1、茎横切总面积S2,则:
通气组织百分率(孔隙度)=(S1/S2)×100%
其中S1为茎的节处或节间横切面中所有通气组织腔隙的总面积,S2为节或节间横切面中茎的总面积。孔隙度的计算能够以数据的形式反映出茎的节处、和节间不同时期通气组织的发达程度,其值的大小与发达程度呈正相关。
1.4 数据处理
得到全部的计算结果,通过IBM SPSS statistics 22.0分析软件进行试验数据的统计计算,进行方差分析前对所有数据进行了正态性及方差齐性检验,用LSD单因素方差分析 (One-Way ANOVA)对茎中节及不同时期通气组织的差异性进行检验,并用Excel 2003作图。
2 结果与分析
2.1 穿叶眼子菜茎的结构
通过对穿叶眼子菜茎纵切(图1,a)的观察,其茎的结构包括表皮、皮层及维管柱,直径随距茎尖距离的增加而变大。通气组织发达,存在于内皮层与表皮之间,且主要存在于节间(图1,c、e),而节处(图1,b、d)的皮层细胞排列紧密仅形成少量且腔隙小的通气组织,孔隙度约为3.62%(图2)。通气组织在距茎尖约0.6 mm处开始形成,并随着距茎尖距离的增加其通气组织腔逐渐增大最终在约2.4 mm处形成成熟的通气组织。通过节间(图1,c、e)通气组织腔内的残留细胞可确定其通气组织腔的膨大过程伴随着皮层细胞的降解。
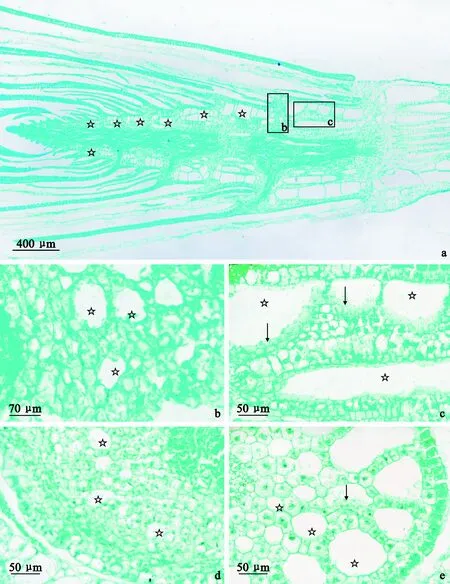
a.茎纵切面(示茎整体结构);b.节纵切(示节纵向通气组织);c.节间纵切(示节间纵向通气组织);d.节横切(示节横向通气组织);e.节间横切(示节间横向通气组织);b、c图为a图的局部放大。“☆”指示通气组织部位,箭头指示残留细胞图1 穿叶眼子菜茎纵切面及节和节间横、纵切a. Stem longitudinal section (indicating the whole structure of the stem); b. Node longitudinal section (indicating longitudinal aerenchyma of the node); c. Internode longitudinal section (indicating longitudinal aerenchyma of the internode); d. Node transverse section (indicating transverse aerenchyma of the node); e. Internode transverse section (indicating transverse aerenchyma of the internode); Fig.b and c are partial enlarged views of Fig.a. “☆” indicates the location of the aerenchyma, and the arrows indicate the residuary cellsFig.1 Stem longitudinal section and node, internode longitudinal section and transverse section of P. perfoliatus

图中不同字母表示差异显著(P<0.05),n=6图2 节及各时期茎通气组织占横切面比例Different letters in the figure indicate significant differences (P<0.05),n=6Fig.2 Stem transverse section aerenchyma area ratio of each phase and node
2.2 穿叶眼子菜茎通气组织的发育过程
在图3中展示了通气组织形成的4个时期的纵切与横切的对应图片。处于实心期(距茎尖0~0.6 mm)的细胞较小且排列紧密,此时的细胞体积处于增长阶段且细胞间没有明显的间隙,因此,现阶段的茎尚未形成通气组织。虽然实心期前期(图3,b)的茎仅为一团尚未明显分化的细胞构成,但处于后期(图3,c)的茎已分化出表皮、皮层及维管束,且细胞体积明显增大,细胞间开始出现小的间隙,间隙约占横截面的0.54%(图2),为下阶段通气组织的形成做准备。
形成期(距茎尖0.6~1.2 mm)细胞体积继续增大,于实心期后期形成的间隙处细胞分离形成小的腔隙(图3,e),通气组织开始形成,此时的腔隙大小仅相当于1~3个细胞。后期(图3,f)通气组织腔的数量较前期明显增加,茎为椭圆形且在两极形成约3轮腔隙,靠近中间部位形成约2轮较小的腔隙。后期腔隙已开始膨大,且部分腔隙间残留有正在降解的细胞,表明腔隙的膨大伴随着细胞降解,每个独立形成的腔隙之间存在一层或数层细胞;整个形成期的平均孔隙度为10.90%(图2)。从此时的纵切图(图3,d)可以看到处于上下相邻位置的通气组织之间也存在至少一层细胞将其相互隔开,所以此时的通气组织在空间上仅为独立的小腔室而并未上下联通。对于通气组织的形成部位,显示其主要存在于节间,而节的细胞紧密排列且形成的通气组织少。

a.实心期纵切(示实心期茎纵向结构);b、c.实心期横切(示实心期前期及后期横向茎结构);d.形成期纵切(示形成期茎纵向通气组织);e、f.形成期横切(示形成期前期及后期茎横向通气组织);g.膨大期纵切(示膨大期茎纵向通气组织);h、i.膨大期横切(示膨大期前期及后期茎横向通气组织);j.成熟期纵切(示成熟期茎纵向通气组织);k、l.成熟期横切(示成熟期茎横向通气组织)。“☆”指示通气组织部位,箭头指示残留细胞图3 穿叶眼子菜的茎纵切与横切a. Longitudinal section of solid phase (indicating the longitudinal stem structure); b, c. Transverse section of solid phase (indicating the transverse stem structure in the early and late phases of the solid phase); d. Longitudinal section of formation phase (indicating longitudinal stem aerenchyma of the formation phase); e, f. Transverse section of formation phase (indicating transverse stem aerenchymain the early and late stages of the formation phase); g. Longitudinal section of expansion phase (indicating longitudinal stem aerenchyma in the expansion phase); h, i. Transverse section of expansion phase (indicating transverse stem aerenchymain the early and late stages of the expansion phase); j. Longitudinal section of maturity phase (indicating longitudinal stem aerenchyma in the maturity phase); k, l. Transverse section of maturity phase (indicating transverse stem aerenchyma in maturity phase). “☆” indicates the location of the aerenchyma, and the arrows indicate the residuary cellsFig.3 Longitudinal section and transverse section of P. perfoliatus stems
从横切图(图3,f)可以看出靠近外皮层处的通气组织腔稍大,说明在横向上通气组织由外向内先后形成。
膨大期(距茎尖1.2~2.4 mm)的细胞进一步增长,腔数继续增加,且孔隙度达到了约27.61%(图2),腔内仍有降解残留的细胞。通过此时的纵切图(图3,g)可看到在纵向上节间通气组织腔开始融合扩大并形成网状结构,但之间仍有较多横向细胞层将其隔开。膨大期前期(图3,h)除了通气组织腔的扩大,维管束中间也开始形成髓腔,内皮层细胞、表皮细胞及外皮层细胞排列整齐且致密,细胞结构保持完整;后期(图3,i)茎中间形成3个髓腔,包括中央维管束包围的大髓腔以及两边由较小维管束包围的小髓腔,此时通气组织腔的数量已基本稳定,腔内仍存在降解的细胞;后期的外皮层细胞及最外面一轮通气组织腔周围的细胞开始降解坍缩。对比膨大期前期与后期茎的横切面积与通气组织腔的大小,后期相较于前期都明显增大,此时通气组织为蜂窝状且腔隙排列较为规律,腔的体积由外向内依次减小。
成熟期即距茎尖约2.4 mm之后为通气组织发育的最后一个时期,此时的通气组织发达且已完全成熟为茎横切面的57.58%(图2)。纵向(图3,j)上此时的通气组织基本融合为长的管道且直径更大,偶尔存在降解残留的细胞结构组成的线状结构将其隔开。横向(图3,k、l)上也表现为通气组织腔的扩大,且最外面一轮通气组织腔周围的细胞层已坍缩为线状结构。内皮层细胞较为完整,外皮层细胞存在部分降解,表皮细胞基本完整但排列不再规则。
2.3 穿叶眼子菜叶通气组织的发育过程
穿叶眼子菜不仅在茎中存在发达的通气组织,其叶中同样存在。通过整体的横切图(图4,a)可直观的观察到其通气组织的发育过程,且和茎中一样将其发育过程划分为同样的四个时期(图4,b~e)。叶中的通气组织仅在主脉中形成,主脉处由表皮、皮层薄壁细胞及维管柱构成,其叶片仅由上下2层表皮细胞及中间的1层叶肉细胞紧密结合而成且与侧脉一样并不形成通气组织。
新生的叶片(图4,b)短小,由3层细胞组成且仅在主脉处细胞团开始分化形成维管束,此时的细胞较小且排列紧密,仅在部分薄壁细胞间有很小的空隙,通气组织尚未开始形成,因此处于实心期。
随着叶片细胞的持续生长分化,其宽度与厚度增加,在主叶脉处位于上下表皮细胞之间的单个或多个薄壁细胞凋亡降解形成小的腔隙(图4,c),这标志着叶片通气组织的发育进入形成期。此时期发生于靠近茎尖的第2~3片新生叶,形成特征与茎通气组织类似,即腔隙的大小相当于1~3个细胞且独立的腔隙之间存在一层或多层细胞。主脉的横切也近似椭圆形,通气组织腔先形成于两极。此时的表皮细胞细胞质浓厚且排列规则、紧密,维管束细胞致密无腔隙形成。
叶片继续发育并开始形成不明显的侧脉(图4,a),但侧脉处并不形成通气组织。膨大期(图4,d)的细胞体积及通气组织腔进一步增大,腔数量有所增加,成因与茎通气组织的膨大期相似,部分腔隙周围存在正在降解的细胞。通气组织腔之间的细胞层在膨大期后期基本减少为一层且开始坍缩降解。此时的表皮细胞排列规则紧密,保持完整;维管束中间开始形成髓腔。
进入成熟期(图4,e)通气组织腔达到最大,腔隙之间的线状结构为之前细胞层降解残留的细胞结构。此时维管束中间已形成明显的髓腔,上下2层表皮细胞基本完整且排列较为规则、紧密。
3 讨 论
生存环境的不同必然导致植物发育与基本结构的差异,如陆生植物茎内皮层和维管束发达,皮层内一般无发达的通气组织,叶片上有大量气孔分布[24];高寒地区植物茎内机械组织发达,叶内有2层以上栅栏细胞而机械组织不发达[25]。湿地挺水植物机械组织较陆生植物退化,但维管组织发达,叶结构与陆生植物类似且存在气孔[26-27],通气组织较发达。沉水植物茎、叶机械组织退化,维管组织不发达,叶内少有栅栏组织及海绵组织的分化,叶表面无气孔,茎叶通气组织发达[23,28-29]。本研究的穿叶眼子菜是一种沉水植物,其茎、叶中的通气组织发达,而体内维管组织不发达,因此茎的机械强度不大,这有利于其随水流摆动;眼子菜属植物的整个叶片内充满气腔或仅主脉周围的叶肉内具气腔[20],穿叶眼子菜属于后者,其叶片薄,仅3层细胞,叶表面无气孔且仅在主脉形成通气组织,表皮细胞具叶绿体,这有利于对光照的吸收,增加光合能力。谢伟东等[28]对3种海草的茎叶解剖结构的研究中,矮大叶藻与二药藻的叶横切面无明显主脉,通气组织均匀形成于叶片内,与本实验中穿叶眼子菜仅在叶主脉形成通气组织存在差异,而喜盐草的叶横切面与穿叶眼子菜类似;矮大叶藻与二药藻的茎横切面为椭圆形,通气组织为较发达的蜂窝状,这与穿叶眼子菜相似。另一种植物海神草的茎存在数束外环维管束[29],且其叶存在数条维管束,维管束之间形成发达的通气组织,而穿叶眼子菜存在1束中央维管束和对称的2束外环维管束,且叶仅在主脉维管束周围形成通气组织。高晨光等[23]对鸭舌草等4种水生植物营养器官结构的研究中,发现其茎、叶均具有发达的通气组织,其中一种俗称水案板的浮叶眼子菜茎横切面为圆形,有小而排列紧密的表皮细胞,中柱处形成4个髓腔,皮层通气组织为网状,本实验中的穿叶眼子菜仅形成3个髓腔且皮层通气组织为较规则的2~4轮,但这2种眼子菜茎通气组织均为蜂窝状,符合属间模式的稳定[18];而这种浮叶眼子菜叶横切面上下表皮为排列紧密的小细胞,上表皮下为2~3层细胞组成的栅栏组织,栅栏组织下为网状通气组织,明显其叶的结构及通气组织与穿叶眼子菜存在差异。

a.叶的横切(示各时期叶横切及通气组织);b.实心期横切(示实心期叶横向结构);c.形成期横切(示形成期主脉横向通气组织);d.膨大期横切(示膨大期主脉横向通气组织);e.成熟期横切(示成熟期主脉横向通气组织)。“☆”指示通气组织部位,箭头指示残留细胞图4 穿叶眼子菜的叶横切a. Transverse section of leaves (indicating leaves transverse section and aerenchyma in each phase); b. Transverse section of solid phase (indicating transverseleaf structure in solid phase); c. Transverse section of formation phase (indicating transverse aerenchyma in main vein of formation phase); d. Transverse section of expansion phase (indicating transverse aerenchyma in main vein of expansion phase); e. Transverse section of maturity phase (indicating transverse aerenchyma in main vein of maturity phase)“☆” indicates the location of the aerenchyma, and the arrows indicate residuary cellsFig.4 Transverse section of P. perfoliatus leaves
Hydrocharismorsus-ranae的不定根中在距根尖约0.3 mm处皮层细胞开始相互分离产生间隙[30],而穿叶眼子菜茎通气组织形成的实心期后期部分细胞间出现裂隙,且随着细胞进一步生长、分离,于形成期前期(距茎尖约0.6 mm)在靠近外皮层处的皮层细胞间形成小的腔隙;H.morsusranae根在距根尖1 mm内由于中皮层细胞裂解而进行平周与垂周分裂,形成气室,而穿叶眼子菜在形成期后期及膨大期(距茎尖约1.2~2.4 mm)的横切及纵切面腔隙中也存在降解残留的细胞,说明腔隙的扩大涉及细胞降解的过程。穿叶眼子菜叶通气组织的形成过程与茎相似,靠近茎尖的第1~2片新生叶细胞排列紧密,无空隙,类似于Typhaangustifolia处于实心期的第1~2片新生叶[5],T.angustifolia于约第3片新叶出现细胞分离产生的小腔隙,且出现晶状体细胞,之后的腔隙膨大过程这些晶状体细胞发挥重大作用;穿叶眼子菜也在第2~3片新叶主脉形成裂生腔隙,而无晶状体细胞出现,说明与T.angustifolia叶腔隙的膨大存在差异,但随后腔隙的膨大也涉及细胞的降解。通过细胞降解而形成的通气组织被认为是细胞程序性死亡(programmed cell death, PCD)的结果,比如水稻茎叶[2,31]、格菱叶柄[6]、水淹的玉米[8,10]、小麦[9]、向日葵[11]等植物通气组织通过降解而形成,这些细胞在逐渐死亡的过程中表现出核DNA片段化特点,以及液泡膜破裂、囊泡的出现、细胞器和核降解、液泡膜破裂、细胞壁最终降解等过程,符合细胞程序性死亡的特征。穿叶眼子菜茎、叶通气组织自形成期后,开始出现细胞降解的方式来进一步扩大通气组织的腔隙,推测这个过程是PCD作用的结果。
不同的植物其通气组织发达程度与形成特点同样存在差异[32],水生植物Egeriadensa茎的结构及通气组织形成模式与穿叶眼子菜相似,其先是早期在距茎尖1~2 mm处的皮层细胞间形成裂生腔隙,在距茎尖2~3 mm处形成蜂窝状的成熟通气组织,腔隙面积为截面的27%[17]。而穿叶眼子菜茎通气组织的形成期早于E.densa,起始于约距茎尖0.6 mm处,成熟于距茎尖约2.4 mm处,与E.densa相似,但穿叶眼子菜茎成熟腔隙面积为截面的约57.58%,远高于E.densa的27%,这也与穿叶眼子菜的沉水生活方式相适应,更为发达的通气组织对呼吸作用产生的CO2及光合作用产生的O2具有一定的存储与扩散作用,这对于提高植物的光合作用效率及营养生长存在积极意义,从而降低了其对外界的O2及无机碳源的需求;发达的通气组织增加其在水中的浮力以获取更多的光照,光照强度对沉水植物的光合作用具有一定的影响作用[33]。另外,穿叶眼子菜茎的节处通气组织形成较少,甚至部分节处完全不形成,这与一种湿地植物Meionectesbrownii的解剖结构类似,这种植物平均节间横截面通气组织孔隙率为17.7%,而节处约为8.2%,甚至约23%的节处无通气组织形成[34]。这在一定程度上阻碍了穿叶眼子菜体内气体的扩散,但提高了茎节内的氧气利用效率,这对穿叶眼子菜的繁殖和分布来说是一种非常高明的适应机制,因为穿叶眼子菜的茎段在自然环境下可以进行无性繁殖,茎段上节为节间的气体扩散提供了一个屏障,为茎段不定根的生长提供氧气保证;同时,节的存在提高了植物体的机械力,弥补了其维管组织不发达,维持了其在水环境中的结构稳定。对于穿叶眼子菜茎、叶通气组织膨大期后期腔隙之间线状结构的形成,类似于水稻根部通气组织腔之间由坍缩细胞的细胞壁聚集而成的结构[3],Jeffree等[35]在Phaseolusvulgaris的叶腔隙形成的最后阶段也观察到细胞壁的降解。在T.pseudoincisa叶柄通气组织成熟期腔隙周围存在不完全降解的细胞,残留的细胞壁维持了其叶柄形态结构[6],因此,穿叶眼子菜茎、叶成熟通气组织腔之间的细胞坍缩一方面是为了进一步增加腔体积,另一方面其残留细胞壁得以维持茎叶形态结构,这都是其对沉水生活的适应机制。
猜你喜欢
护理研究(2022年14期)2022-08-06 08:59:48
中华养生保健(2020年9期)2021-01-18 03:11:50
中国临床医学影像杂志(2019年4期)2019-06-18 10:54:50
中学课程辅导·教师教育(上、下)(2018年15期)2018-10-10 07:56:54
益寿宝典(2018年32期)2018-01-26 19:36:36
中国马铃薯(2015年3期)2015-12-19 08:03:54
微型小说选刊(2015年16期)2015-11-17 17:03:05
参花(下)(2015年8期)2015-06-11 10:50:05
今传媒(2015年12期)2015-05-30 05:12:32
小小说月刊(2015年11期)2015-05-14 14:55:32
