
黄土高原北部日本弓背蚁巢穴结构特征及其影响因素*
2018-08-27 03:29:16邵明安李同川贾玉华贾小旭黄来明
土壤学报 2018年4期
杨 析 邵明安,2,,4† 李同川 贾玉华 贾小旭 黄来明
(1 西北农林科技大学资源环境学院,陕西杨凌 712100)
(2 中国科学院水利部水土保持研究所,陕西杨凌 712100)
(3 中国科学院地理科学与资源研究所,北京 100101)
(4 中国科学院大学资源与环境学院,北京 100190)
(5 沈阳农业大学水利学院,沈阳 110866)
自1999年国家实施退耕还林还草措施以来,黄土高原植被覆盖度由20世纪70年代的6.5%上升至2010年的50%左右[1],生态环境有了较大改善。植被的大量恢复给土壤动物提供了大量的食物和适宜的生存空间[2]。植被枯枝落叶、根系分泌物和微生物代谢形成的有机质、根冠脱落下来的细胞、死亡的根毛以及较老的根区上濒于死亡的外皮细胞等均为土壤动物的食物来源[3],植被的枯枝落叶还为土壤动物提供隐蔽的生存空间。
蚯蚓、地鼠和蚂蚁等土壤动物活动是影响土壤大孔隙形成和演化的一个重要因素[4-6]。研究土壤动物巢穴结构及其影响因素,有助于加深对黄土高原土壤大孔隙结构类型的理解。土壤的孔隙结构对降水的转化、储存、利用以及污染物在土壤中迁移有重要影响[7-8]。此外,土壤大孔隙发育特征对水和溶质输移也有重要影响[9]。土壤动物通过进出土体获得生存空间和食物,在其生命活动过程中改变土体结构,形成具有水稳性的管状大孔隙通道[3]。有研究表明,在植被生长茂盛的土壤中,树鼩、鼹鼠、地松鼠和小鼠等大型土壤动物常常挖掘直径较大、连通性较好的土壤通道,鼠类形成的通道直径可达数十厘米[10-11]。很多研究证实,在潮湿气候区,蚯蚓的生命活动过程中常产生土壤大孔隙[12],蚯蚓形成的大孔隙直径在2~11 mm之间,通道深度最大70 cm左右[13]。一方面,蚯蚓通过挖掘洞穴改变了土壤孔隙结构,改善了土壤通气透水能力;另一方面,蚯蚓在生命活动过程中将地表的枯落物带入洞内增加了土壤有机质来源,而且蚯蚓排泄物的有机质含量较高,因此,可增加其洞穴内部的总有机质含量,进而促进形成更多的次生孔隙[3]。在土壤较为干旱的黄土高原北部地区,蚂蚁是形成土壤大孔隙的重要土壤动物,蚂蚁通过挖洞来储存食物和生存繁殖,从而形成大孔隙[14]。据Benckiser[15]研究,蚂蚁几乎在所有的陆地生态系统中均有发现,且它们的数量占全部昆虫数量的50%以上,大部分蚂蚁在地下筑巢,结构复杂,其巢穴在一定程度上可改变土壤结构和土壤孔隙。调查发现,黄土高原北部神木县六道沟流域有日本弓背蚁(Camponotus japonicus)、针毛收获蚁(Messor aciculatus)、红林蚁(Formica sinensis)、草地铺道蚁(Tetramorium caespitum)和小家蚁(Monamorium pharaonis)等大量不同种类的土栖性蚂蚁,其中,日本弓背蚁体型最大,数量最多,属于优势种。日本弓背蚁在中国大部分地区均有分布[16],属于蚁亚科,工蚁体长7.4~13.8 mm。张家明[3]研究了拟黑多刺蚁(Polyrhzchis vicina Roger)的巢穴结构,表明拟黑多刺蚁的巢穴结构复杂,通道间互相联通,但是巢穴深度仅为30 cm左右。总体而言,关于黄土高原蚂蚁巢穴结构的研究并不多见。
土壤动物的活动及群落规模易受土壤含水量、土壤利用方式以及土壤质地等因素的影响,进而间接影响土壤大孔隙的形成。免耕和少耕均能够改善土壤保水、保土等物理性状[17],有利于土壤动物的生存繁殖,进而有利于土壤动物形成土壤大孔隙。与蚯蚓相比,蚂蚁不能在水分过大的土壤中生存。调查表明,蚂蚁在含水量适宜的黄土、砂土、粉砂土和砾石中均有发现。不同类型的土壤中蚂蚁巢穴结构的基本组成相似[18],由纵横的通道、“生育室”及“垃圾堆放室”等组成[19]。但是,不同种类蚂蚁巢穴的结构有一定的差别,这主要与蚂蚁的体型大小有关。同时,蚂蚁的活动和巢穴分布受捕食者、植被、孔隙水盐度和颗粒尺寸等生物、非生物因素的影响[20]。本文将土壤物理性质的影响作为主要研究目标,探索了土壤含水量、土壤质地和土壤容重对日本弓背蚁巢穴结构的影响。
研究土壤动物巢穴结构最常见最简单的传统方法是直接挖掘法[6,21],此法可直接观察巢穴内部结构并记录蚂蚁数量,但该法对土壤结构破环较大,挖掘、记录过程具有不完整性。Tschinkel[22]用稀薄的石膏浆灌入蚂蚁巢穴,石膏硬化后挖掘得到巢穴的石膏模型。这种方法获取的巢穴结构更为完整,立体感更强。Bastardie等[23]用电子计算机断层扫描法(CT)获取蚯蚓的三维孔隙结构。此法操作简单、省时省力,更重要的是可以保持土样的原状结构,且能通过三维手段对土壤孔隙网络结构进行定量分析。本文采用较为简单实用的石膏浇注法获取蚂蚁巢穴结构。本研究的主要目标是:(1)对自然条件下的日本弓背蚁巢穴结构进行实地调查,探索不同群落规模下日本弓背蚁巢穴的结构特征;(2)对比分析神木县六道沟小流域两种典型土栖性蚂蚁(日本弓背蚁和针毛收获蚁)成熟巢穴的结构特征;(3)土壤质地、含水量和容重对日本弓背蚁巢穴结构的影响。
1 材料与方法
1.1 蚂蚁个体野外采集方法
在黄土高原北部神木县六道沟流域选择土质疏松利于挖掘的砂壤土采集蚂蚁(日本弓背蚁)个体。发现蚂蚁后在附近寻找其巢穴,找到巢穴后首先将巢口的枯枝落叶等杂物清理干净,然后顺着巢穴入口用铁铲沿着其通道挖掘,巢穴挖开后大量的工蚁从巢穴内出来,此时用改装过的吸尘器收集蚂蚁个体,将蚂蚁转移至塑料瓶内并在瓶内放入适量的巢内原土,进行遮光处理后带回实验室备用。
1.2 野外蚂蚁巢穴模型的获取——石膏浇筑法
蚂蚁(日本弓背蚁和针毛收获蚁)巢穴结构获取的前期处理与收集蚂蚁个体前期处理相同,找到巢穴并清理杂物后,沿着巢口向巢穴内间歇性缓慢注入事先准备的自来水(尽量减少水流对土壤结构的破坏),迫使蚂蚁从洞内爬至地面,根据蚁后和工蚁数量来判断蚁群规模。当蚁后出现并过一段时间无工蚁出来时停止向巢穴内注水,用自来水和石膏粉按一定比例(比例不定,以石膏不沉淀为基本标准,通常水与石膏粉的比例大于10)配成的石膏浆通过巢口注入巢穴内,直至石膏浆充满整个巢穴为止。等待24 h,石膏浆完全凝固后,先用铁铲从巢穴旁边挖一个剖面,然后用小刀和毛刷将巢穴的石膏模型取出,带回实验室对其进行清洗和测量。
用游标卡尺测量蚂蚁巢穴石膏模型的巢穴入口、通道直径、巢穴长度等结构特征。本实验用体积置换法测量巢穴的总体积,首先将清洗后的蚂蚁巢穴石膏模型浸泡在自来水中12 h,确保石膏模型内部完全湿润,然后将巢穴石膏模型放入装满水的容器中,用量筒量取溢出水的体积,将溢出水的体积视为石膏模型的体积。
1.3 室内试验设计
采集神木县六道沟小流域的砂土、壤砂土和壤土(土壤质地和有机质含量见表1),经风干、磨碎、过筛(2 mm)后备用。室内试验所用蚂蚁均为日本弓背蚁。
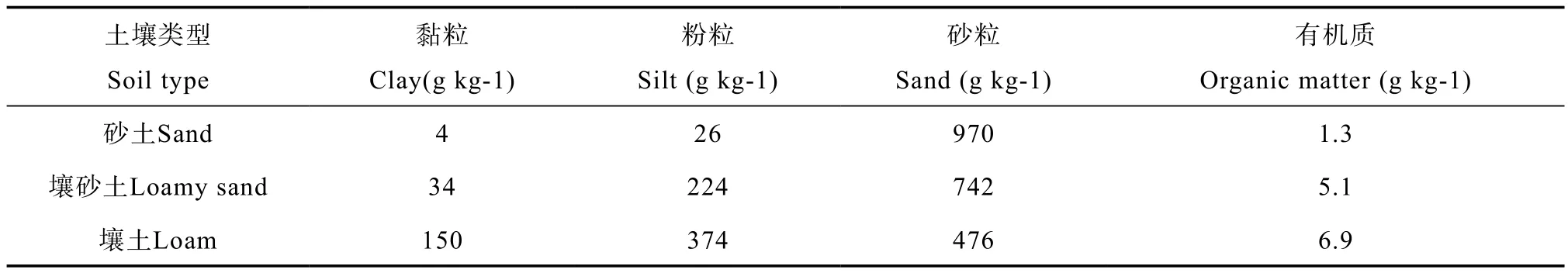
表1 六道沟流域不同土壤质地组成及有机质含量Table 1 Soil textures and organic matter contents in the Liudaogou Catchment
工蚁数量对巢穴体积的影响试验:将处理后的壤土置于直径20 cm、高20 cm的圆柱形铁桶中,容重控制在1.4 g cm-3,含水量控制在100 g kg-1左右,桶内分别放入0、10、30、50、100、150、200只工蚁,每处理3个重复,并在铁桶内壁喷涂防逃液。蚂蚁在土柱中筑巢并将土壤颗粒搬运至土壤表层,当蚂蚁进入土壤内部后,用改装的吸尘器收集地表的土壤团粒并风干称重。最后对蚂蚁数量和搬运至地表的土壤质量进行回归分析。
土壤质地对蚂蚁巢穴结构的影响试验:将壤砂土、壤土和砂土放入直径30 cm、高40 cm的塑料桶,填充高度为36 cm,桶底部密封、上部敞开,给土柱加水使土壤含水量在100 g kg-1左右,每处理3个重复。将野外获取含有蚁后的蚁群200只放入土柱内,并在桶内边缘喷抹防逃液防止蚂蚁逃出土柱。当蚂蚁停止向地表搬运土粒时,实验结束并利用石膏浇筑法获取巢穴结构模型,并测量体积、直径和长度等巢穴结构信息。
土壤含水量对蚂蚁巢穴结构的影响试验:将处理过的砂土装入带有小孔盖子的透明塑料盒中,将塑料盒内砂土的含水量(质量含水量)分别控制在5、30、60、100、150、200和250 g kg-1,每处理4个重复。每天18:00 称重,并补加蒸发的水量保持不同处理的含水量不变,观察巢穴结构并记录繁殖结果(工蚁和蚁卵数量),分析土壤含水量对蚂蚁巢穴结构的影响。
土壤容重对蚂蚁巢穴结构的影响试验:将处理后的砂土放入直径30 cm、高40 cm的塑料桶,填充高度为36 cm,桶底部密封、上部敞开,给土柱加水使土壤含水量在100 g kg-1左右,每处理3个重复。填装的砂土容重分别为1.2 g cm-3和1.5 g cm-3,然后将带有蚁后的蚂蚁群落(1只蚁后和100只工蚁)置于土柱内,蚂蚁停止挖掘后,用石膏浇筑法获取巢穴结构模型,并测量巢穴总体积、分支、节点等巢穴结构信息。
1.4 数据处理
本文采用描述性统计方法对比不同群落规模以及不同种类蚂蚁巢穴特征。用SPSS18.0软件对蚂蚁数量和搬运至地表的土壤重量进行回归分析。用Origin8.0 和Photoshop CS 6.0软件进行制图和巢穴结构照片的处理。
2 结果与讨论
2.1 日本弓背蚁巢穴结构特征
不同群落规模的日本弓背蚁巢穴结构如图1所示。日本弓背蚁的巢穴结构是由相连的通道和巢室组成,这与Karlen等[19]的研究结果相同。野外条件下,日本弓背蚁巢穴的通道总体趋势是垂直向下的,巢室是倒置的漏斗状或立体的椭圆形。当蚁群规模仅有十几只时,其巢穴结构简单且体积较小(图1a),巢穴深度约为10 cm,通道呈圆柱形,仅有一个巢室供蚁后产卵并哺育后代,巢穴总体积仅有3~4 cm3。随着蚂蚁数量的增加,蚂蚁巢穴深度逐渐增加,通道和巢室依次相连,形成多级巢室且通道有横向延伸的分支(图1e和图1f)。
在六道沟流域内对日本弓背蚁巢穴的挖掘,统计蚁群数量和相关数据测量表明:日本弓背蚁最大种群数量在520只左右,巢穴最深可达63cm左右,通道直径4.1~6.6 mm,横截面面积606~2117 mm2。虽然日本弓背蚁巢穴规模大小随着蚁群数量的增加而增加,但这并未改变巢穴的基本组成结构,依然是由通道和巢室构成。随着日本弓背蚁个体数量的增加,其巢穴深度、巢室数量、巢穴的分支以及巢穴的复杂程度均在增加,但蚁群数量的增加不会显著改变通道直径和巢室的横截面面积。
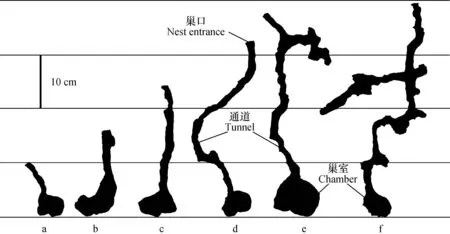
图1 不同群落规模日本弓背蚁巢穴结构特征(a-f群落规模分别是:15、32、60、100、165和248只)Fig.1 Characteristics of ant (Camponotus japonicus) nests relative to community size (a-f stands for community size, a: 15, b: 32, c:60, d: 100, e: 165 and f: 248)
在室内模拟日本弓背蚁工蚁数量对巢穴体积的影响时,由于工蚁沿着桶壁向下挖掘巢穴,影响石膏浆的灌注,这种情况下以蚂蚁搬运至地表土壤颗粒的量视为其巢穴的体积。如图2所示,对工蚁数量和搬运至地表的土壤质量分析,结果显示,土壤表层堆积的土壤质量随着工蚁群落规模的增加而增加,两者之间有显著的回归关系(p<0.001),工蚁搬运至地表的土壤质量和引入土壤的工蚁数量高度相关(R2=0.892 7)。蚂蚁的数量或者群落规模决定了其在土壤中的生存空间,继而影响了挖掘至土壤表面的土壤质量。由于蚂蚁数量越多时需要更大的生存空间,蚂蚁搬运至地表的土壤就越多,导致其巢穴规模越大,这与自然条件下日本弓背蚁巢穴规模大小随着蚁群数量的增加而增加的结果相符合。蚂蚁属于群居性昆虫,依靠工蚁群体挖掘洞穴,洞穴的规模随群体的发展而增大。而独居性昆虫挖掘的洞穴则随个体生长阶段的变化而变化,Bailey等[24]研究了美国3种蝼蛄在不同生长阶段和不同质地土壤中的巢穴结构,结果表明:蝼蛄制造的孔隙直径为其身体宽度的2.5倍,成熟阶段的蝼蛄挖掘的洞穴体积为未成熟阶段的3倍左右。
自然条件下大部分日本弓背蚁巢穴属于单巢口类型(图3a),单巢口巢穴横向发展较小,纵向发展较大。图3b和图3c,此类蚁巢有2~3个巢穴入口,其巢口直径与单巢口直径大小相同。与单巢口类型巢穴相比,双巢口或多巢口巢穴有更长的横向延伸。对六道沟流域内日本弓背蚁巢穴的调查结果表明:日本弓背蚁巢穴的巢口在1~3个之间变化,大多数巢穴仅有一个巢口,仅有个别巢穴有2个或者3个巢口。根据王常禄和吴坚[25]研究,蚂蚁在春季开始活动时常修建新的巢穴入口。巢口增加有利于调节巢室内的温度、湿度及便于蚂蚁出入巢穴搬运食物等行为[26]。因此,巢口的增加不仅有利于蚂蚁的生存发展,而且还可能会对降水、土壤溶质和氧气等物质的迁移起到促进作用。
室内饲养条件下日本弓背蚁巢穴结构和自然条件下巢穴结构特征不同。室内试验将200只工蚁和蚁后放置于聚氯乙烯(PVC)管内的土壤表面,工蚁沿PVC管内壁的多个位置挖掘洞穴,随着挖掘的深入,地下空间逐渐变大。当地下空间足够大时,蚁后被移入巢内,此时,其他工蚁停止挖掘并进入蚁后所在巢穴继续挖掘。工蚁沿着PVC管壁旋转向下挖掘,到达底部后沿着PVC管底部环形挖掘。饲养条件下日本弓背蚁巢穴巢口在1~3个之间,巢穴基本结构依然是由通道和巢室构成,通道直径在4.1~6.6 mm之间,这些特征和自然条件下相同;但自然蚁群巢穴主要在垂直和水平两个方向延伸,而饲养条件下仅在垂直方向上旋转延伸。根据Villani等[27]对蝼蛄的室内研究表明:室内用PVC管或树脂玻璃等材料做土柱试验时,会限制蝼蛄的横向发展,使蝼蛄沿着管壁旋转向下挖掘洞穴,并且洞穴的深度和体积受到试验土柱大小的影响。因此,室内饲养条件下所用PVC管的大小限制了日本弓背蚁巢穴发展,使其巢穴的深度以及通道的延伸趋势与自然条件下不同。

图2 工蚁搬运至地表的土壤质量随工蚁数量的变化Fig. 2 Volume of the soil moved up onto the surface by worker ants relative to number of worker ants
2.2 自然条件下日本弓背蚁和针毛收获蚁巢穴结构对比
日本弓背蚁和针毛收获蚁是神木县六道沟流域两种常见的土栖性蚂蚁,自然条件下其成熟巢穴结构如图4所示。
两种蚂蚁巢穴结构均由通道和巢室组成且逐渐发展形成多级巢室,通道的总趋势均是垂直向下发展,但日本弓背蚁有横向发展的分支。两种蚂蚁的巢室形状有所不同:日本弓背蚁巢室为倒置的漏斗形或立体的椭圆形;针毛收获蚁巢室为扁平形,巢穴体积主要集中在巢穴的上半部分且巢穴体积随着深度的加深而减少,巢室间的间隔也逐渐增大。此外,两者巢穴深度明显不同(表2),日本弓背蚁巢穴深度达600 mm,而针毛收获蚁巢穴深度可达1 500 mm。自然条件下日本弓背蚁巢穴巢口在1~3个之间,巢口直径在4.1~6.6 mm之间;而针毛收获蚁仅有1个巢口,巢口直径在2.3~3.2 mm之间。日本弓背蚁巢室横截面面积、巢室数量和巢室间距均小于针毛收获蚁。针毛收获蚁相比于日本弓背蚁有更深的巢穴和更多的巢室,这可能与针毛收获蚁的生活习性有关,一方面,较深的巢穴有助于针毛收获蚁抵御巨大的昼夜温差并保证群落越一。在许多其他土壤动物巢穴的研究中也得到了类似结果:食草性蝼蛄的洞穴主要存在于土壤的表层,食肉性蝼蛄趋于将巢穴建造至较深的土层[27]。土白蚁的地下巢穴深度仅有几毫米,而高等白蚁的巢穴可达地下5~15 cm[29]。根据它们自身的尺寸,蜣螂制造的洞穴直径在1~50 mm 之间变化,巢穴深度在10~103 cm之间[30]。六道沟流域日本弓背蚁和针毛收获蚁在生活习性、群落大小及体型等方面存在差异,所以两种蚂蚁的巢穴结构在巢口数量、巢口直径、巢穴深度、巢室数量、巢室面积和巢室间距等方面均有显著差异。
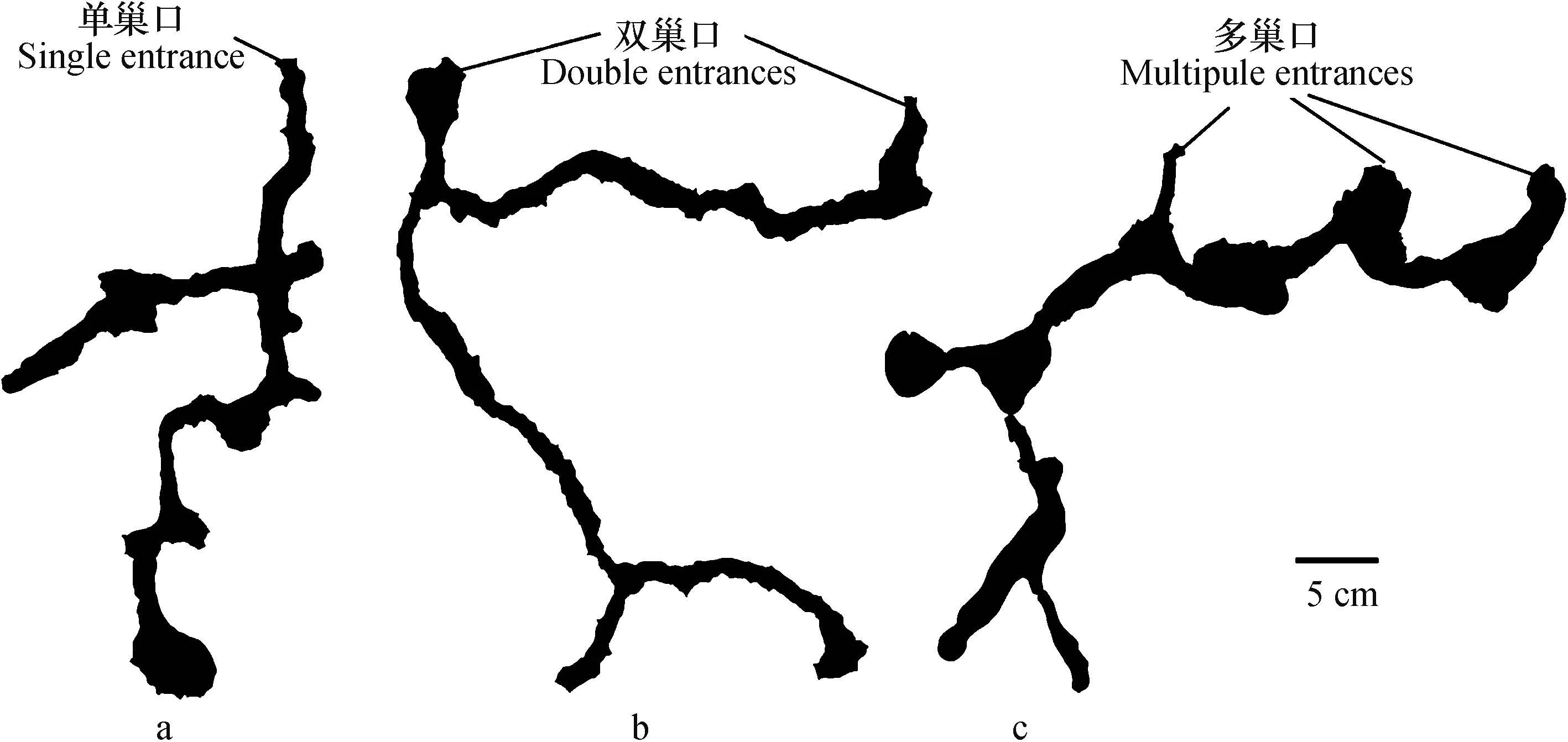
图3 不同数量巢口的日本弓背蚁巢穴(a:单巢口,b:双巢口,c:多巢口)Fig. 3 Varying numberof nest entrances of Camponotus japonicus (a: Single entrance, b: Double entrances, c: Multipule entrances)

图4 日本弓背蚁(a)和针毛收获蚁(b)巢穴对比Fig. 4 Comparison of ant nest between Camponotus japonicas(a) and Messor aciculatus (b)
2.3 土壤质地、含水量和容重对日本弓背蚁巢穴结构的影响
在六道沟流域调查表明:日本弓背蚁在壤土和壤砂土有较多分布,而在颗粒较大的砂土中分布较少。通过室内试验在一定容重和适宜的土壤含水量下研究日本弓背蚁在壤土、壤砂土、砂土三种土壤中的巢穴特征。日本弓背蚁在三种土壤中巢穴巢口均仅有一个且巢口直径在4.1~6.6 mm之间,由于巢穴深度受土柱长度的限制导致巢穴的深度仅有36 cm左右,巢室平均为1~2个。此外,三种不同土壤的巢穴巢室的横截面面积、巢室间距等均无明显差异,因此,在一定容重和适宜的含水量条件下,不同质地土壤对日本弓背蚁巢穴结构特征无明显影响。冬;另一方面,针毛收获蚁属于食种子类动物,在针毛收获蚁巢穴中,种子以种类和形态的不同被分类、分层保存于不同深处的巢室[26]。根据陈元瑶等[28]研究敏捷扁头猛蚁(Pachycondyla astuta)的巢穴结构,其巢穴深度在5~25 cm之间,通道更贴近地面,巢口一个至多个不等,巢穴较为简单。因此,蚂蚁种类是决定巢穴结构差异的重要因素之

表2 日本弓背蚁和针毛收获蚁巢穴结构特征Table 2 Characteristics of nest structure (Camponotus japonicus and Messor aciculatus)
通过室内试验在一定容重的砂土条件下,研究日本弓背蚁在不同含水量下的巢穴结构特征。砂土含水量在30~200 g kg-1之间时,日本弓背蚁可挖掘巢穴且巢穴的基本结构特征与野外条件相同;当含水量低于30 g kg-1或大于200 g kg-1时,蚁群无法在土柱内筑巢。由于过干或过湿的砂土容易坍塌,导致蚂蚁无法筑巢,仅有在含水量适宜时砂土结构稳定才能支撑蚂蚁巢穴。Espinoza和Santamarina[31]认为,在颗粒较大的砂土中,蚂蚁巢穴的通道大部分是垂直分布,主要是因为砂土中蚂蚁洞穴内部的土壤颗粒大多处于承重状态,不允许巢穴通道的横向发展,但是本研究并未发现类似现象。此外,含水量对蚂蚁的生长繁殖有更大的影响,表3显示了日本弓背蚁蚁后在不同含水量条件下的生存和繁殖能力。
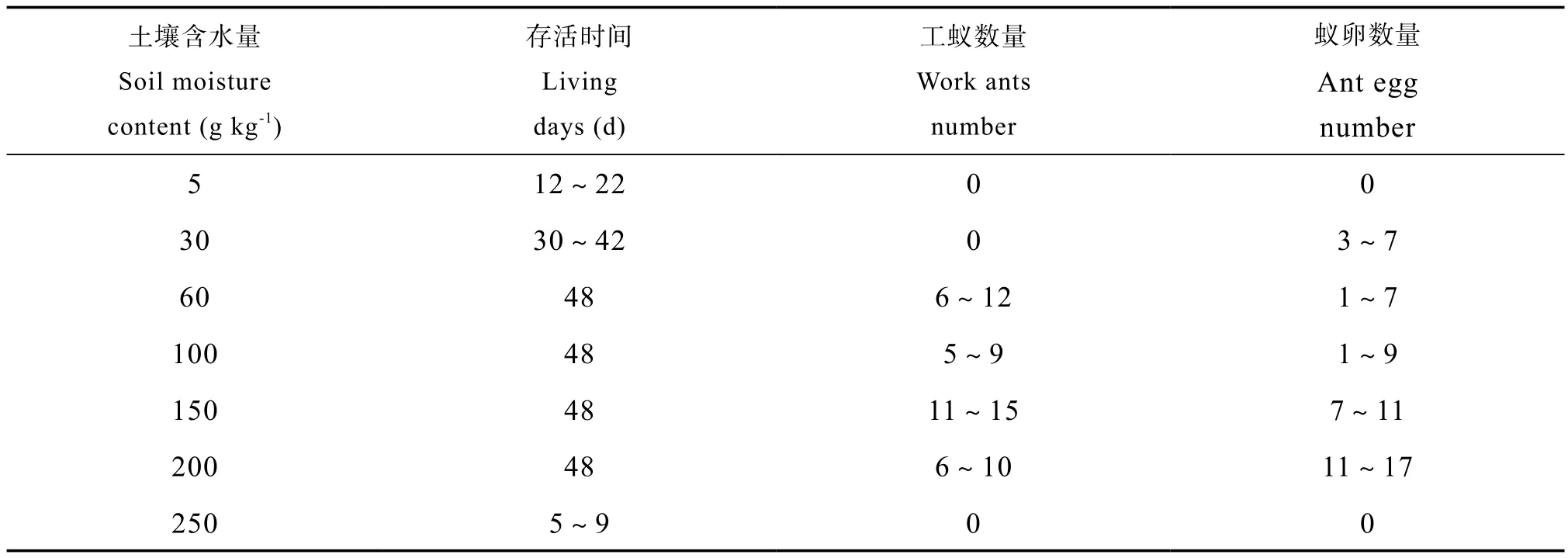
表3 不同含水量条件下日本弓背蚁蚁后的生存及繁殖能力Table 3 Surviving and propagating capabilities of queens ant (Camponotus japonicus) relative to soil moisture content
日本弓背蚁蚁后在含水量5 g kg-1的砂土中未挖掘巢穴,也未繁殖,在17 d左右因缺水死亡;在含水量30 g kg-1的砂土中,蚁后可产卵并挖掘巢穴,但未繁殖出工蚁;在含水量60~200 g kg-1的砂土中,蚁后产卵、筑巢、繁殖出工蚁;在含水量250 g kg-1的砂土中,蚁后不能产卵、筑巢、繁殖工蚁,并在7 d左右死亡。由此可见,含水量对蚂蚁的生存、筑巢、繁殖均有较大影响,含水量的变化可直接影响蚁群的规模从而引起巢穴规模的变化。但是在含水量30~200 g kg-1之间时,其巢穴通道直径、巢室横截面面积、巢口数目等巢穴结构特征无明显变化。因此,不同含水量条件对蚂蚁的生存和繁殖能力影响较大,从而影响蚁群的规模再而引起巢穴规模的变化,但不会对巢穴的结构产生明显影响。
野外调查显示,土壤容重较大时日本弓背蚁巢穴深度较浅,更趋向于纵向发展。室内模拟试验在其他条件一致的情况下,仅改变容重,探究不同容重下的日本弓背蚁巢穴特征,结果表明:不同容重下巢穴结构有明显的差异(图5)。土壤容重在1.2 g cm-3时,巢穴通道的长度、分支、巢穴体积等均大于土壤容重在1.5g cm-3时的巢穴,土壤容重为1.2 g cm-3时巢穴更趋向于横向发展且整体结构较为复杂。
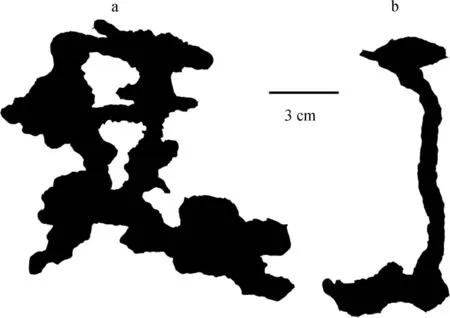
图5 不同容重下(a:1.2 g cm-3和b:1.5 g cm-3)日本弓背蚁巢穴结构Fig. 5 Sturcture of typical ant (Camponotus japonicus) nests relative to soil bulk density (a: 1.2 g cm-3 and b: 1.5 g cm-3)
土壤容重为1.2 g cm-3时巢穴通道分支有9个,节点为5个,巢室总体积为13.6cm3,这些均大于土壤容重为1.5g cm-3时的巢穴(表4)。但土壤容重的变化未显著改变巢穴的巢口直径。根据Espinoza和Santamarina[31]研究:在中低含水量条件下的低容重壤土中,蚂蚁巢穴朝各个方向充分发展,洞穴通道分支较多。与本研究结果类似,在低容重下蚂蚁巢穴结构更加复杂。由于土壤容重会影响土壤的密度,土壤容重越大土壤密度越大,导致土体结构较硬,蚂蚁不易挖掘,因此,在土壤容重较大时,蚂蚁巢穴结构简单,通道单一,总体积、节点和分支数量等结构特征指标均较低。

表4 不同容重下日本弓背蚁巢穴总体积、分支数量和节点数量Table 4 Total volume, number of branches, and number of node number of ant (Camponotus japonicus) nests relative to soil bulk density
3 结 论
自然条件下日本弓背蚁巢穴结构由垂直向下的通道和倒置漏斗形的巢室组成,巢口直径4.1~6.6 mm,巢口1~3个,巢穴结构随着蚂蚁群落增加而不断发展,巢穴最大深度可达63 cm左右,巢室横截面面积606~2117 mm2。随着日本弓背蚁工蚁数量的增加,土壤表层堆积的土壤质量也逐渐增加,两者之间有显著的回归关系。室内试验条件下,由于PVC管的限制,使日本弓背蚁巢穴深度在30 cm左右,巢穴通道旋转向下且无横向发展的分支。日本弓背蚁和针毛收获蚁在巢室形状、巢穴深度、巢口直径、巢口数量、巢室间距等有明显的差异。不同土壤质地和不同含水量条件下,日本弓背蚁巢穴结构特征无明显差异。土壤容重较大时,蚂蚁巢穴结构简单,通道单一,总体积、节点和分支数量均较少。但是,工蚁数量、土壤质地、土壤含水量和土壤容重均不会显著影响日本弓背蚁巢穴的通道直径,通道直径主要与蚂蚁的体型相关。
猜你喜欢
植物保护(2024年3期)2024-06-08 07:17:41
农技服务(2020年7期)2020-12-17 20:45:50
农技服务(2020年6期)2020-12-17 13:38:06
科学之谜(2018年10期)2018-01-02 10:08:28
创新作文(3-4年级)(2017年3期)2017-09-18 11:03:23
环境昆虫学报(2017年4期)2017-09-16 05:29:31
幼儿园(2017年2期)2017-03-11 20:34:21
创新作文(小学版)(2017年8期)2017-02-25 03:25:46
少年博览·小学高年级(2016年4期)2016-04-16 00:13:14
党的生活(2016年2期)2016-03-12 01:02:46
