
富民枳种质资源的SSR遗传多样性研究*
2018-08-25 07:12:40张珊珊甘云浩杨文忠段宗亮丁红茜康红梅诺苏那玛
西部林业科学 2018年4期
张珊珊,甘云浩,杨文忠,段宗亮,丁红茜,康红梅,诺苏那玛
(1.云南省林业科学院,云南 昆明650201;2.国家林业局云南珍稀濒特森林植物保护和繁育重点实验室,云南省森林植物培育与开发利用重点实验室,云南 昆明650201;3.昆明市濒危动植物收容拯救中心,云南 昆明650224;4.宾夕法尼亚州立大学理学院,宾夕法尼亚斯泰特科利奇16563)
物种保护的最终目标是保存足够的遗传多样性[1-3],从而保持其适应生境变化的潜力。遗传多样性研究不仅能反映出濒危植物多样性的高低,还能为制定有针对性的保育策略提供科学依据[4-6]。有学者研究表明,很多濒危植物种群的遗传多样性水平低,可能是导致其濒危的主要原因,并在研究基础上提出了相应的保护措施。但是,究竟何种保护措施是有效的,目前并没有统一的标准。研究指出,珍稀濒危植物保护的终极目标在100年内保持其90%以上的遗传多样性才能称之为 “成功保护”[7-8]。因此,在采取保育措施时,如何根据目的物种遗传多样水平的高低,确定种群的有效样本量大小,才能既不会造成人力等各种资源的浪费,又达到 “成功保护”的目标,需要进一步研究[9-13]。
富民枳 (Poncirus polyandra)属典型的极小种群野生植物,其天然种群已经灭绝[14]。目前,对富民枳采取了一些保护措施,共建立9个种质资源收集圃和1个野外回归种群。然而,富民枳种质资源的遗传多样性水平如何,采取的保护措施在遗传水平上是否有效,都不得而知。
利用分子标记研究种质资源的遗传多样性,可为其遗传管理提供理论依据[15-16]。SSR遗传标记因其具有重复性、稳定性高的优点,广泛应用于遗传多样性研究[17-18]。因此,本研究采用SSR分子标记对富民枳进行遗传分析,旨在获得准确可靠的遗传多样性信息,进而评价种质保存和种群恢复是否达到 “成功保护”的目标,为开展基于遗传多样性结果的其他有效遗传管理策略提供科学依据。
1 材料与方法
1.1 试验材料
从富民枳的9个种质资源收集地和1个人工回归地,共采集170份新鲜叶片样品 (表1)。在详细记录植株编号的同时,于2016年春季采集健康幼嫩的叶片,用硅胶快速干燥并拍照,带回实验室,用75%质量分数的无水乙醇擦拭清洗,最后用液氮冷冻贮存以备提取DNA。
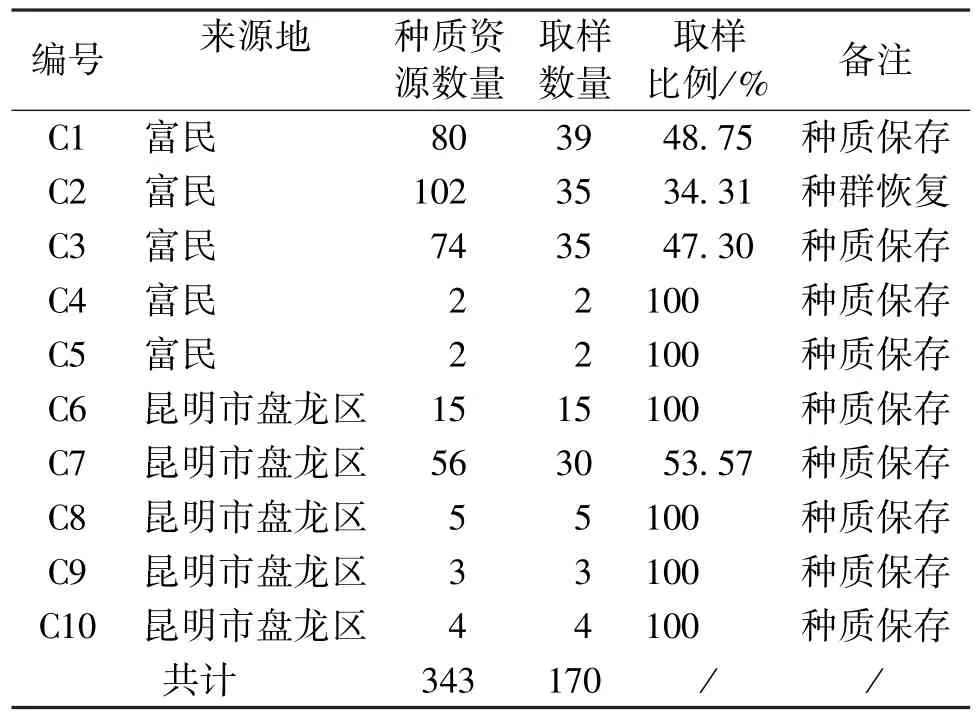
表1 供试材料Tab.1 Plant materials for this study
1.2 试验方法
1.2.1 DNA提取
采用改良CTAB法提取基因组DNA[19]。0.8%琼脂糖凝胶电泳检测其质量,紫外分光光度计检测DNA浓度,合格样品置于-20℃保存备用。吸取2L DNA和6L 1×溴酚蓝混合进行琼脂糖凝胶电泳,300V电压下跑12min。
1.2.2 PCR反应体系及扩增程序
基于张珊珊等[19]筛选出的5对具片段多态性的引物,对170个样本进行PCR扩增。采用15L体系如下:2×Taq Master Mix 7.5L,正向引物1L(10mmol/L),反向引物1L(10mmol/L),模板DNA1L,灭菌ddH2O 4.5L。PCR反应程序:94℃预变性5min,94℃变性30s,63℃退火30s,72℃延伸20s,30个循环;72℃延伸10min,4℃保存。将扩增好的PCR产物进行琼脂糖凝胶电泳 (2L样品+6μL溴酚蓝),300V电压下12min,获取鉴定胶图。
1.2.3 毛细管电泳
通过胶图确定模板浓度,加水稀释到电泳所需浓度。上机进行毛细管电泳:(1)将HiDi与500bp的内标按130∶1混合,配成mix;(2)用国产96孔反应板分装mix,每个孔中加入10μLmix;(3)对应着在96孔板中加入0.5μL样品模板,离心到4 000rpm即停;(4)用金属浴加热器将混合板95℃加热预变性5min,拿出后立即放入-20℃冰箱;(5)冷却后拿出,4 000rpm离心30s,解冻、混匀;(6)上3730测序仪进行毛线管电泳;(7)获取下机结果并分析,筛选出多态性较好的位点。
1.3 数据分析
采用POPGENE Version 1.32软件计算有效等位基因数 (Ne)、平均期望杂合度 (He)、观测杂合度 (Ho)和Shannon信息指数 (I);利用PIC_Calc 0.6软件计算各引物的多态性信息含量(PIC)值;利用Gene mapper 4.1进行数据准确位点的分析后,通过Populations 1.2.30软件构建邻接 (Neighbour-joining)树;利用FigTree v1.4.0软件绘制聚类图;利用Origin 7.0软件对富民枳遗传多样性水平变化趋势进行拟合分析。
2 结果与分析
2.1 SSR-PCR扩增产物的多态性分析
从图1看出,毛细管电泳检测结果准确可靠。富民枳的遗传多样性较低:平均PIC值为0.337 8,平均 Nei's遗传多样性指数为0.430 0,平均Shannon信息指数 (I)为0.642 8(表2)。不同引物组合的有效等位基因数平均为1.798 7;其平均期望杂合度 (He)为0.430 0,平均观测杂合度(Ho) 为0.568 7。

图1 富民枳部分样品在Ciclev10001597m引物处扩增的毛细管电泳检测Fig.1 Capillary electropherogram obtained by Ciclev10001597m primer of some samples from Poncirus polyandra

表2 基于5对SSR引物所扩增的产物在170份富民枳种质资源中的遗传多态性Tab.2 Genetic diversity estimated based on the amplified products by 5 SSR primers among 170 germplasms resources of Poncirus polyandra
2.2 聚类分析
利用邻接法 (Neighbor-joining)对富民枳样本进行聚类分析 (图2)。由图2可知,170个富民枳样本共聚类了65个分支末梢,属于同一个分支末梢的富民枳被默认为同等的遗传关系。在每个末梢抽取一个样本的基础上,重新计算5对不同引物的扩增片段在抽取的这65个有效样本资源中的多态性 (表3),并绘制相应的聚类图 (图3)。

图2 基于邻接法绘制的170个富民枳样本聚类图Fig.2 Dendrogram of 170 Poncirus polyandra samples based on neighbor-joining clustering method

表3 基于5对SSR引物所扩增的产物在65份富民枳种质中的遗传多态性Tab.3 Genetic diversity estimated based on the amplified products by 5 SSR primers among 65 germplasms of Poncirus polyandra
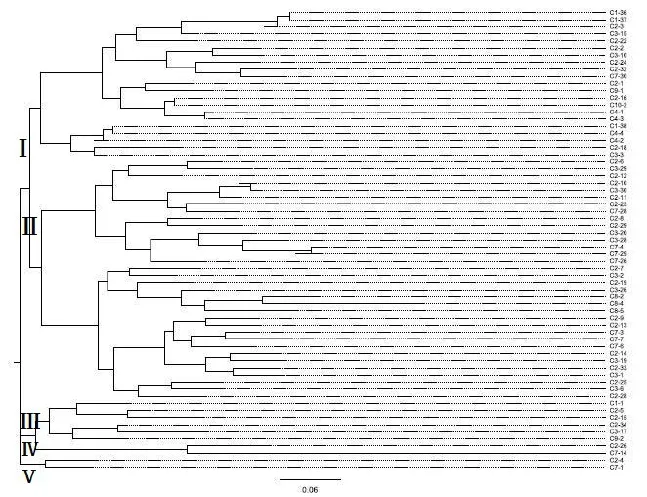
图3 基于邻接法绘制的65个富民枳样本聚类图Fig.3 Dendrogram of 65 Poncirus polyandra samples based on neighbor-joining clustering method
由表3可知65个有效富民枳样本资源的多态性:多态性信息含量 (PIC)均在0.25-0.5之间,PIC范围是0.302 6-0.429 5,平均0.353 6;Shannon信息指数 (I)平均为0.681 6,范围是0.562 3-0.865 8。平均期望杂合度 (He)为0.442 3,范围是0.346 8-0.541 0;平均观测杂合度 (Ho)为0.431 9,范围是0.323 1-0.515 6。由图3可知,富民枳的65个有效样本资源共分为5个亚类 (Ⅰ类-Ⅴ类),其中Ⅰ类包括来自C1、C2、C3、C4、C7、C9和C10共7个地方的21个样本资源,Ⅱ类包括来自C2、C3、C7和C8共4个地方的34个样本资源,Ⅲ类包括来自C1、C2、C3、C7和C9共5个地方的6个样本资源,Ⅳ类包括来自C2和C7两个地方的2个样本资源,Ⅴ类也包括来自C2和C7两个地方的2个样本资源。
2.3 评价不同种质资源收集地的遗传多样性
富民枳10个种源地170份样本的遗传多样性计算结果见表4。由表4可知,10个来源地的遗传多样性各不相同,其中,C1、C5、C6、C8和C10计5个地方的Shannon信息指数在0.502 9-0.567 4之间;C2、C3、C4、C7和C9共5个地方Shannon信息指数在0.609 8-0.640 8之间。基于前文 “关于属于同一分支末梢的部分富民枳样品遗传关系可能相同”的结论,将聚类分析中系统发育树每个末梢的所有样本按所属地进行分组,即同一末梢 (基因型相同)来自相同地方的样本只取1个分配到相应的地方,也就保证每个地方保存的样本均是来自不同末梢,使每个地方的样本之间不存在重复。
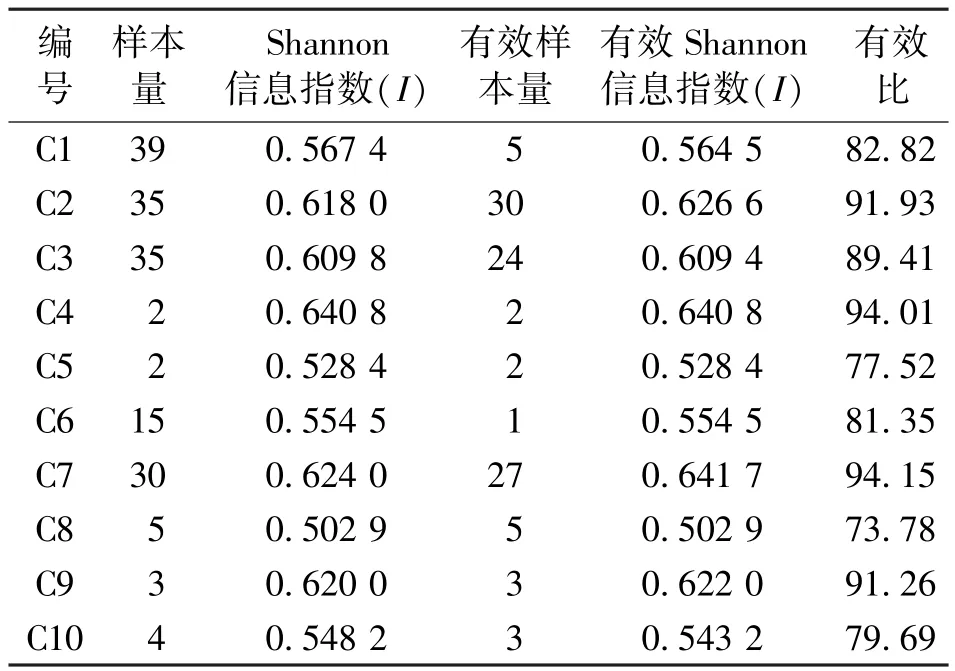
表4 富民枳在不同样本来源地的遗传多样性Tab.4 Genetic diversity of Poncirus polyandra in different origins of samples
3 讨论
3.1 富民枳的遗传多样性
物种遗传多样性保护是生物多样性保护的重要部分[20],遗传多样性研究对于了解濒危物种在未来环境下的适应或进化以及物种保护具有重要意义。对于极小种群野生植物,开展其遗传多样性研究可为制定能保持物种遗传多样性和进化潜力的综合保育措施提供相应的科学依据[21-25]。据报道,部分极小种群野生植物,例如巧家五针松 (Pinus squamata)[26]、云南蓝果树 (Nyssa yunnanensis)[27]、水松 (Glyptostrobus pensilis))[28]和版纳青梅 (Vatica xishuangbannaensis)[29]等的遗传多样性都偏低,遗传分化水平较高。本研究显示,170份富民枳种质资源的遗传多样性 (I=0.642 8)要低于65份富民枳种质资源的的遗传多样性 (I=0.681 6),其原因可能是聚类图2中同一个分支末梢的种质材料遗传关系相同。多态性信息含量(PIC)均值为0.337 8,Nei's遗传多样性指数(Nei)和有效等位基因数 (Ne)都偏低,杂合度观测值与杂合度期望值也相差甚远,表明群体遗传多样性很低[30]。因此,综合分析表明富民枳种质资源的遗传多样性水平较低,采取保护措施必须建立在最大程度保护其遗传多样性的基础上。
3.2 基于富民枳遗传多样性研究的种质保存和种群恢复评价
分析种群遗传多样性和确定有效种群规模是当前保护遗传学研究的主要内容。国内外基于遗传多样性分析提出了不同物种的保护措施[31-33],这些研究在比较各种分子标记方法有效性的同时,根据种群间和种群内的遗传多样性,提出了注重种群的保护策略[34-37]。前期研究用SSR分子标记分析有效样本量与遗传多样性指数的关系,结果显示随着有效样本量的增加,富民枳遗传多样性随之增加。有效样本量小于29时,遗传多样性变化幅度较大,当有效样本量大于29时,遗传多样性趋于稳定 (I=0.691 0±0.002 0),这说明利用遗传多样性指数来衡量富民枳种质资源的遗传多样性水平,至少需要29株。进一步对现存富民枳遗传多样性分析结果进行S型曲线拟合发现,有效样本量大于10时,遗传多样性才能达到90% (I=0.621 9)以上[23]。因此,无论开展天然种群保护还是人工种群的重建与恢复,都应在本研究 “65株”种质资源的基础上,选取不少于10株的有效样本材料 (或复本),就可以最大程度上保存富民枳现有种质资源的遗传多样性。但是为了避免 “样本量过多导致人力和物力的浪费”,应尽量将种群的样本量配置控制在“65株”有效种质资源的29株 (或复本)以内。
富民枳目前野外天然种群已经灭绝,对其现存种质资源的科学保护刻不容缓。从65份富民枳种质资源材料的聚类图看,可将其分为5大类。这5大类的植株分别来自7种、4种、6种、2种和2种种质资源收集地,也就是说从C1到C10,富民枳的遗传多样性分别来自于聚类图中的2(Ⅰ和Ⅲ)、 5(Ⅰ-Ⅴ)、 3(Ⅰ-Ⅲ)、 1(Ⅰ)、 0、 0、 4(Ⅰ~Ⅳ)、1(Ⅱ)、2(Ⅰ和Ⅲ) 和1(Ⅰ) 类,可以认为C2和C7两个种质资源收集地的富民枳样本的来源最丰富。另外,本研究对每个来源地的富民枳样本分别进行了遗传多样性分析,结果表明遗传多样性的差异性很大。其中,C2(I=0.626 6)、C4(I=0.640 8)、C7(I=0.641 7) 和C9(I=0.622 0)计4个地方的Shannon信息指数都达到总样本的90%(I=0.621 9)以上,但是有效样本量大于10的只有C2(30个有效样本)和C7(27个有效样本)两个地方。综合聚类图分析结果,可以认为只有人工回归地C2和种质资源收集地C7现阶段的遗传管理是有效的。
3.3 保护建议
基于对富民枳现存种质资源遗传多样性的分析结果,建议采取以下保护措施: (1)完善种质资源收集地的资源配置。在8个未达到保护目标的种质资源收集地,从65株有效样本聚类图的每个亚类中选择精选植株 (或其复本)进行搭配,为保证每个收集地90%以上的遗传多样性,有效样本量不少于10株,使得种质资源保存合理化,同时降低富民枳DNA的位点纯合率,避免近交衰退;(2)开展回归引入,恢复天然种群。在前期建立人工种群1个基础上,在天然种群邻近区域,逐年回归种植富民枳苗木,新建天然种群2-3个;(3)开展近地保护措施,建立其人工种群。在气候相似、生境相似和群落相似的自然或半自然地段培植管护富民枳,并采集以种子为主的繁殖材料进行资源配置,从65株有效样本中选择不少于10株的繁殖材料 (或其复本)作为精选群体重建种群;(4)开展相应的监测工作。对备份种质和重建种群的遗传多样性变化及其幅度进行监测,以保证富民枳种质资源收集地具有较高的遗传多样性水平。
4 结论
本研究结果表明保存下来的富民枳遗传多样性水平较低。拟合样本量与Shannon遗传多样性指数之间的S型曲线发现,在有效样本量为10时,遗传多样性达到群体水平的90%;在有效样本量至少为29时,其遗传多样性基本能代表富民枳群体的遗传水平。进而评价了种质保存基地和野外恢复种群的遗传多样性,9个种质资源保存基地中只有1个保存基地达到保护目标,目前采取的种群恢复策略在遗传水平上是有效的。本研究为实施富民枳其它科学保育策略的实施提供了理论基础。
猜你喜欢
今日农业(2022年13期)2022-09-15 01:18:00
金桥(2022年4期)2022-05-05 06:09:56
内蒙古统计(2021年4期)2021-12-06 02:49:20
加油站服务指南(2021年7期)2021-10-14 07:28:14
测控技术(2018年4期)2018-11-25 09:46:52
当代陕西(2018年12期)2018-08-04 05:49:12
中国麻业科学(2018年6期)2018-04-09 11:22:12
上海精神医学(2017年5期)2017-11-29 06:03:10
湖南农业(2017年5期)2017-05-13 09:24:30
西南农业学报(2016年5期)2016-05-17 05:42:21
