
啮齿动物主动母性行为动态改变的神经机制*
2018-08-13 08:01张一帆齐星亮蔡厚德
心理科学进展 2018年8期
张一帆 齐星亮 蔡厚德,
(1南京师范大学心理学院,南京 210097) (2南京晓庄学院幼儿师范学院,南京 211171)
1 引言
主动母性行为(active maternal behavior)是雌性哺乳动物在哺乳期内有效照料幼崽的一种动机行为,涉及组织与调节一系列母性反应(maternal responsiveness)以适应幼崽发展需要的神经生物学过程(Pereira,2016; Pereira & Ferreira,2016)。哺乳动物幼崽在出生后并不能立即独立生活,因而雌性的照料决定了幼崽的存活率和生理、行为发育状况(Dobolyi,Grattan,& Stolzenberg,2014;Pereira & Ferreira,2016)。在人类中,母亲的照料可以提供充分的刺激和训练,使婴儿在运动、感知、认知和社会情绪能力等方面得到良好发展(Lonstein,Lévy,& Fleming,2015)。另外,成为母亲的过程也伴随着巨大的生理改变和心理压力,导致哺乳期女性容易出现产后抑郁(postpartum depression)等心理问题(Kim,Strathearn,& Swain,2016)。产后抑郁的母亲在育幼时比较迟钝、刻板和易怒,使婴儿长大后倾向于淡漠和社交困难(Olazábal et al.,2013b)。因此,阐明哺乳动物主动母性行为及其神经机制,不仅可以为探究行为进化和早期发展开辟新的研究领域,也对人类母亲产后抑郁等临床问题的干预有借鉴意义。
考察主动母性行为的神经机制通常采用一些侵入性的方法(如脑组织损伤等),但这类实验不能安全地运用于人类,因此研究者通常基于动物实验的结果对主动母性行为的一般机制提出假设,以便采用非侵入性手段在人类中加以验证(Lonstein et al.,2015)。在非人类哺乳动物中,实验室大鼠(rat)和小鼠(mouse)等啮齿动物的主动母性行为得到了最为详尽的研究(Lonstein et al.,2015)。大鼠和小鼠的主动母性行为包括对幼崽的衔回(retrieving)、舔舐/理毛(licking/grooming)和嗅探(sniffing)等子成分(Afonso,King,Chatterjee,&Fleming,2009; Numan,2006)。在整个哺乳期(产后0~22天)内,雌鼠的主动母性行为经历了从产后早期的发动(onset) (产后 0天)和维持(maintenance) (产后 0~11 天)阶段,到产后晚期的衰退(decline) (产后 12~22天)阶段的动态改变(Olazábal et al.,2013b; Pereira & Ferreira,2016)。在这一过程中,雌鼠能适应性地调整主动母性行为的表现,以满足幼崽不同发育阶段的需要(Pereira,2016)。较早的研究表明,主动母性行为的发动主要与分娩激素事件有关(Moltz & Wiener,1966); 在激素水平下降后,主动母性行为的维持转而受到幼崽刺激的调节(Rosenblatt,1967;Reisbick,Rosenblatt,& Mayer,1975; Rosenblatt &Siegel,1981); 到了产后 12天左右,雌鼠与幼崽的接触减少,主动母性行为开始走向衰退(Reisbick et al.,1975; Pereira & Morrell,2011)。近期,采用条件化位置偏好(conditioned place preference,CPP)任务的研究(Pereira & Morrell,2010; Wansaw,Pereira,& Morrell,2008)发现,啮齿动物主动母性行为的动态改变与幼崽奖赏价值(incentive value)的阶段性变化有关。值得关注是,还有大量研究(Cortés-Mendoza,Díaz De León-Guerrero,Pedraza- Alva,& Pérez-Martínez,2013; Numan et al.,2010; Benedetto,Pereira,Ferreira,& Torterolo,2014; Numan & Young,2016;Sabihi,Dong,Durosko,& Leuner,2014;Stolzenberg & Champagne,2016)探查了主动母性行为动态改变的神经机制。结果表明,幼崽奖赏价值变化引起的主动母性行为动态改变,不仅涉及分娩激素事件开启下丘脑内侧视前区(medial preoptic area,MPOA)−中脑腹侧被盖(ventral tegmental area,VTA)−伏隔核(nucleus accumbens,NA)−腹侧苍白球(ventral pallidum,VP)通路,而且需要杏仁核基底外侧核(basolateral amygdala,BLA)的情绪性学习功能以及内侧前额皮层(medial prefrontal cortex,MPFC)的行为选择作用对上述通路进行实时调节。
本文首先利用CPP任务的行为学证据分析幼崽奖赏价值阶段性变化与啮齿动物主动母性行为动态改变的关系; 然后系统阐述产后早期主动母性行为的发动与维持以及晚期衰退的神经机制;最后探讨未来需要研究的一些重要问题或方向。
2 CPP 任务的行为学证据
考察啮齿动物产后不同阶段的主动母性行为可以采用多种行为学方法,如母性行为测试(maternal behavior testing)、杠杆按压(bar-pressing)和 CPP 等(Pereira & Morrell,2009; Olazábal et al.,2013a)。尽管母性行为测试能够反映主动母性行为的变化,但难以评价幼崽对雌鼠的奖赏作用,因此需要将主动母性行为转化为某种操作性行为,如针对幼崽的杠杆按压或趋近行为(Olazábal et al.,2013a)。但是,雌鼠习得的按压杠杆行为如果得不到幼崽的及时强化,继续按压的动机会自然减少,不便于长时间实验观察(Dalley,Cardinal,&Robbins,2004)。与此不同,雌鼠对幼崽的趋近行为会在不能获得幼崽的情况下持续较长时间。因而,研究者(Pereira & Morrell,2009; Pereira &Morrell,2011)常用CPP任务,通过雌鼠趋近幼崽的行为考察幼崽作为一种具有高度奖赏价值的诱因对产后早晚期雌鼠主动母性行为的影响。
在CPP实验中,首先训练雌鼠将两个具有不同知觉特征(如颜色)的场景分别与两个刺激物建立联结,然后移除刺激物,检测雌鼠选择趋向哪个场景(Tzschentke,2007)。在经典CPP任务中,两个场景分别对应一个实验刺激(如幼崽或可卡因)和一个中性的“控制”刺激(如生理盐水) (Seip et al.,2008; Wansaw et al.,2008)。研究(Seip et al.,2008;Wansaw et al.,2008)发现,当对应幼崽的场景与中性场景比较时,产后早期约 80%的雌鼠选择了对应幼崽的场景,而产后晚期 60%以上的雌鼠选择了中性场景; 相比之下,当对应可卡因的场景与中性场景比较时,产后早期和晚期均有约 80%的雌鼠选择对应可卡因的场景。而且,采用同步幼崽/可卡因 CPP任务(concurrent pup/cocaine choice conditioned place preference task)的研究支持以上结果。在同步CPP任务中,两个场景分别对应幼崽和可卡因,以便在可卡因与幼崽竞争的条件下基于可卡因的剂量评价幼崽引发雌鼠主动母性行为的奖赏价值(Mattson,Williams,Rosenblatt,& Morrell,2001)。研究(Pereira & Morrell,2010,2011)发现,产后早期雌鼠对幼崽对应场景和可卡因(1.0 mg/kg)对应场景的偏好基本相同(约 40%);到了晚期,绝大多数(约 80%)雌鼠偏向可卡因对应场景,没有雌鼠选择幼崽对应场景。上述证据(Seip et al.,2008; Wansaw et al.,2008; Pereira &Morrell,2010; 2011)表明,产后晚期雌鼠对幼崽的偏好相对于早期明显下降,但对可卡因的偏好一直较高。这提示产后晚期主动母性行为的衰退并不是一般性动机水平的减少所致,而是幼崽奖赏价值降低减弱了雌鼠的母性动机,因而不能有效引发主动母性行为(Mattson & Morrell,2005;Pereira & Morrell,2011)。
可见,主动母性行为动态改变与幼崽奖赏价值的阶段性变化有关。问题是,从产后早期主动母性行为的发动和维持,到晚期的逐渐衰退,有哪些脑结构和神经通路参与了这一过程的加工与调节?其中涉及怎样的神经机制?以下将梳理相关领域的研究证据加以阐述。
3 主动母性行为动态改变的神经机制
3.1 产后早期主动母性行为的发动
在分娩前后,雌鼠可以被幼崽的气味所吸引。而且,分娩后雌鼠会立即开始对幼崽进行衔回、舔舐和嗅探(Lonstein et al.,2015)。主动母性行为的上述发动过程涉及分娩激素事件开启MPOA-VTA-NA-VP通路,包括三个阶段:(1)分娩激素事件激活 MPOA; (2) VTA-NA中多巴胺(dopamine,DA)水平的提高; (3) NA-VP的去抑制。图1A说明主动母性行为发动的神经通路机制。
3.1.1 分娩激素事件激活MPOA
在未育雌性大鼠,来自嗅球(olfactory bulb,OB)的幼崽嗅觉信号通过杏仁核内侧核(medial amygdaloid nucleus,MeA)与杏仁核皮质核(cortical amygdaloid nucleus,CA)向负责厌恶加工的下丘脑前核(anterior hypothalamic nucleus,AHN)传递,使雌鼠厌恶幼崽(Numan,2006)。但是,分娩激素事件能够激活MPOA,导致嗅觉信号可以经MeA/CA传至 MPOA,从而发动主动母性行为(Numan,2006)。雌鼠分娩时妊娠期的激素变化达到高峰,包括雌二醇(estradiol)水平上升、雌二醇/孕酮(progesterone)比例增加(Fleming,Ruble,Krieger,& Wong,1997; Lonstein et al.,2015)。证据(Numan,Rosenblatt,& Komisaruk,1977)表明,仅在妊娠后期雌鼠的MPOA中埋植雌二醇苯甲酸酯(estradiol benzoate),就可迅速激发主动母性行为。还有证据(Stolzenberg & Champagne,2016)表明,雌激素受体与雌二醇结合后可以协同共激活因子p160向基因的转录起始位点聚集染色质修饰酶(chromatinmodifying enzymes),打开MPOA中处于沉默状态的多种受体基因,导致MPOA能够对MeA/CA传递的嗅觉信号做出反应,进而作用于脑内奖赏系统使幼崽成为具有高奖赏价值的目标(Gagnidze,Weil,Faustino,Schaafsma,& Pfaff,2013; Numan,2006)。
3.1.2 VT A-NA中DA水平的提高
在幼崽气味的刺激下,处于激活状态的 MPOA可以兴奋VTA的DA神经元,从而提升NA中的DA水平,使幼崽奖赏价值得以表达(Stolzenberg& Champagne,2016)。经典 CPP研究(Pereira &Morrell,2010)表明,MPOA 失活后,产后早期的雌鼠失去对幼崽的偏好,不再选择对应幼崽的场景; 而控制组雌鼠中,有 80%的个体选择了对应幼崽的场景。与此类似,VTA失活的雌鼠也完全失去了幼崽偏好; 而控制组有 50%选择了对应幼崽的场景(Seip & Morrell,2009)。这些结果提示,MPOA激活后VTA释放至NA中的DA水平可能决定了幼崽奖赏价值的高低(Seip & Morrell,2009)。研究者(Afonso et al.,2009)推测,NA中的DA活动可能是通过一种基线DA抑制(basal DA suppression)机制赋予幼崽高水平的奖赏价值。未接触幼崽时,分娩雌鼠NA中的基线DA水平显著低于未育雌鼠; 接触幼崽后,分娩雌鼠 NA中的DA水平显著超过未育雌鼠,DA水平的落差有利于雌鼠迅速将幼崽作为具有较高奖赏价值的目标进行关注(Afonso,Shams,Jin,& Fleming,2013;Olazábal et al.,2013a)。
3.1.3 NA-VP 的去抑制
NA中DA水平的快速上升,会触发NA与控制动作程序的VP之间的去抑制机制,使DA活动转化为主动母性行为(Numan & Young,2016)。证据(Sesack & Grace,2010)表明,NA中90%的传出神经元是合成抑制性 γ-氨基丁酸(gamma-amino butyric acid,GABA)的中型多棘神经元(medium spiny neuron,MSN),可以通过D2受体接受DA的抑制性调节。但是,一些研究(D'Cunha,King,Fleming,& Lévy,2011; Parada,King,Li,& Fleming,2008)发现,MSN接受D2受体抑制剂处理并不明显影响主动母性行为,接受催产素(oxytocin)受体抑制剂处理却明显干扰主动母性行为。进一步研究(Romero-Fernandez,Borroto-Escuela,Agnati,&Fuxe,2013)表明,MSN中除了含有D2受体之外,还有催产素受体,两者以异聚体(heteromer)形式并存。在幼崽吮乳刺激下,来自PVN的催产素与催产素受体相结合,可以提升DA与D2受体的亲和力(Romero-Fernandez et al.,2013)。雌鼠接触幼崽前,NA中的DA和催产素水平都较低,MSN处于活动状态,向 VP释放 GABA,使其保持关闭,导致主动母性行为的动作程序不能启动。接触幼崽后,NA中的DA和催产素水平大幅提高。在催产素的调节下,大量DA与D2受体相结合,抑制了MSN的活动,使VP去抑制,导致主动母性行为的动作程序得以启动,最终表现出主动母性行为(Numan & Young,2016)。
3.2 产后早期主动母性行为的维持
分娩数小时后,尽管雌鼠体内的激素水平已经大幅降低(Stolzenberg & Champagne,2016),但雌鼠仍会将幼崽作为高奖赏价值的目标,并维持频繁的主动母性行为(Olazábal et al.,2013b)。在这一阶段,不仅来自幼崽的嗅觉信号可以增加 MPOA中受体基因的表达,而且BLA通过情绪性学习的强化作用以及MPFC通过调节对幼崽的行为选择都可以影响主动母性行为的表现。图1B说明主动母性行为维持的神经通路机制。
3.2.1 嗅觉信号增加受体基因表达的作用
在主动母性行为维持阶段,激素增加 MPOA中受体基因表达的作用会迅速被幼崽的气味等刺激引起的感觉信号所接替(Dobolyi et al.,2014;Olazábal et al.,2013b)。研究(Stolzenberg & Champagne,2016)表明,长时间接触幼崽能够在大鼠和小鼠MPOA中引起一系列细胞间信号的反应,包括钙信号(calcium signaling)、细胞外调节激酶的磷酸化(phosphorylation of extracellular regulated kinase,pERK)和转录因子环磷腺苷反应成分结合蛋白(cyclic AMP response element binding protein,CREB)水平的改变。而且,体内ERK下游产物或CREB的缺乏,会使小鼠主动母性行为受损(Jin,Blendy,& Thomas,2005; Kuroda et al.,2007)。与雌激素受体的作用类似,上述细胞间信号也可以增加 MPOA神经元中多种受体基因的表达(Cortés-Mendoza et al.,2013; Riccio,2010),这使来自幼崽的嗅觉信号也可以激活MPOA,并提升VTA-NA中的 DA水平,进而导致雌鼠继续将幼崽作为高奖赏价值的目标(Peña & Champagne,2015; Numan & Young,2016)。
3.2.2 BLA 的情绪性学习作用
但是,嗅觉信号引起的MPOA激活水平可能较低,主动母性行为的维持还需要负责情绪性学习的BLA对NA-VP去抑制机制的调节(Numan &Young,2016)。证据(Lee,Clancy,& Fleming,1999)表明,损毁BLA会破坏大鼠以幼崽为目标的杠杆按压行为。还有证据(Lee et al.,1999; Numan et al.,2010)显示,BLA的损毁或失活不仅干扰了动物对情绪刺激的联结性学习,而且破坏了主动母性行为。这提示BLA可以操作性地习得关于幼崽的情绪信息,从而促进雌鼠的主动母性行为。BLA及其邻近区域对 NA和 VP各有一条兴奋性谷氨酸(glutamate)神经投射(Numan et al.,2010; Numan &Young,2016)。通往NA的投射含有突触前的D1受体,用药物单独抑制D1或MSN上的D2受体对主动母性行为影响较小,而同时抑制D1和D2受体可有效破坏主动母性行为的持续表现(Parada et al.,2008; Sesack & Grace,2010),提示主动母性行为的维持依赖于D1受体与MSN上D2受体的联合作用。Numan和 Young (2016)认为,上述谷氨酸投射能够兴奋NA中的MSN,使VP保持抑制; 当DA水平上升时,DA与突触前的D1受体结合,使谷氨酸投射的兴奋降低,这会易化 NA对VP的去抑制。而且,在VP完全去抑制的前提下,BLA及其邻近区域通往VP的神经投射可以产生超常激活,导致每次主动母性行为都能加强 VP中神经回路的突触联结,进而使主动母性行为得到强化(Numan & Stolzenberg,2009; Numan &Young,2016)。
3.2.3 MPFC 的行为选择作用
MPFC是脑内整合情绪信息的高级中枢,可以利用从 BLA传来的情绪信号对刺激的奖赏价值进行评估,从而制定行为决策(刘飞,蔡厚德,2010; Dilgen,Tejeda,& O’Donnell,2013)。MPFC包含边缘下区(infralimbic region,IL)和边缘前区(prelimbic region,PrL)等子区域(Pereira & Morrell,2011)。在操作性条件学习的一般过程中,IL负责按照已习得的关于刺激的奖赏价值进行反应,PrL则依据当前的奖赏价值进行反应(Dalley et al.,2004)。采用杠杆按压范式的研究(Balleine &Dickinson,1998; Killcross & Coutureau,2003)表明,PrL失活(IL单独起作用)的大鼠在奖赏不稳定的条件下仍然表现出稳定的条件化反应,失去了对当前奖赏价值的敏感性; 而IL失活(PrL单独起作用)的大鼠反之,如果不能得到及时的奖赏,条件化反应立即减少。换句话说,IL以习惯模式起作用,PrL则以目标导向模式起作用(Dalley et al.,2004)。IL和PrL在产后早期主动母性行为中的调节作用与上述模式一致。同步CPP研究表明,PrL失活(IL单独起作用)消除了雌鼠的可卡因偏好,使 70%雌鼠选择了对应幼崽的场景; IL失活(PrL单独起作用)消除了雌鼠的幼崽偏好,使80%雌鼠选择了对应可卡因的场景; 而控制组中选择两类场景的雌鼠各占40% (Pereira & Morrell,2011)。由此来看,在主动母性行为的维持中,IL可能基于BLA已习得的情绪性经验选择幼崽,表现为习惯模式; PrL则可能基于BLA提供的当前的情绪性经验选择幼崽,表现为目标导向模式(Dalley et al.,2004; Olazábal et al.,2013b)。
研究显示,MPFC的两个子区域调节主动母性行为的神经通路有所不同,这可能与它们的上述功能特性有关。IL主要通过兴奋性投射影响MPOA的激活(Olazábal et al.,2013b),这使主动母性行为倾向于以简单和刻板的方式表现。相比而言,PrL参与主动母性行为调节的机制则比较复杂。PrL中存在至少两组与主动母性行为有关的神经元。第一组是向MPOA投射的兴奋性神经元(Olazábal et al.,2013b),含有催产素受体,接受来自PVN的催产素投射(Sabihi et al.,2014)。证据(Febo,Numan,& Ferris,2005; Sabihi et al.,2014)表明,幼崽的吮乳行为会引起PVN分泌催产素和MPFC的激活,且用催产素受体拮抗剂处理 PrL会损害雌鼠的主动母性行为,提示来自幼崽的感觉刺激可以通过催产素影响 PrL中的兴奋性神经元,进而提高MPOA的激活水平。第二组是向NA中MSN投射的兴奋性谷氨酸神经元,含有突触前的D2受体(Sesack & Grace,2010)。当NA中DA水平上升时,DA与 D2受体结合会削弱 PrL对MSN的兴奋性影响,进而易化 VP的去抑制(Olazábal et al.,2013b; Sesack & Grace,2010)。可见,PrL既可以通过MPOA调控DA的释放,又可以通过 VP去抑制参与主动母性行为的调节,从而使雌鼠的主动母性行为表现更为灵活。
3.3 产后晚期主动母性行为的衰退
与产后7天相比,产后13天左右雌鼠舔舐幼崽的频率从每分钟1次下降到0.5次,而初次衔回的潜伏期从约5秒上升到88秒; 产后22天前后幼崽断奶时,雌鼠的主动母性行为完全终止(Pereira & Morrell,2009; 2011)。产后晚期雌鼠主动母性行为的上述衰退过程与幼崽的独立性需求不断提高有关(Pereira & Ferreira,2016)。证据(Pereira & Morrell,2009; Olazábal et al.,2013b)表明,产后晚期幼崽的视觉、听觉和运动能力明显提升,倾向于独自离巢觅食,并且会抗拒雌鼠的衔回。研究者(Olazábal et al.,2013b)认为,随着幼崽独立性的提高,雌鼠接受到的幼崽感觉刺激会逐渐减少,在主动母性行为中的操作性情绪学习效率也不断降低。伴随这一过程,BLA难以继续习得来自幼崽反应的情绪性信号,导致 VP的去抑制和主动母性行为受到的强化作用减弱,使产后晚期的主动母性行为逐渐减少(Pereira &Ferreira,2016; Olazábal et al.,2013b)。
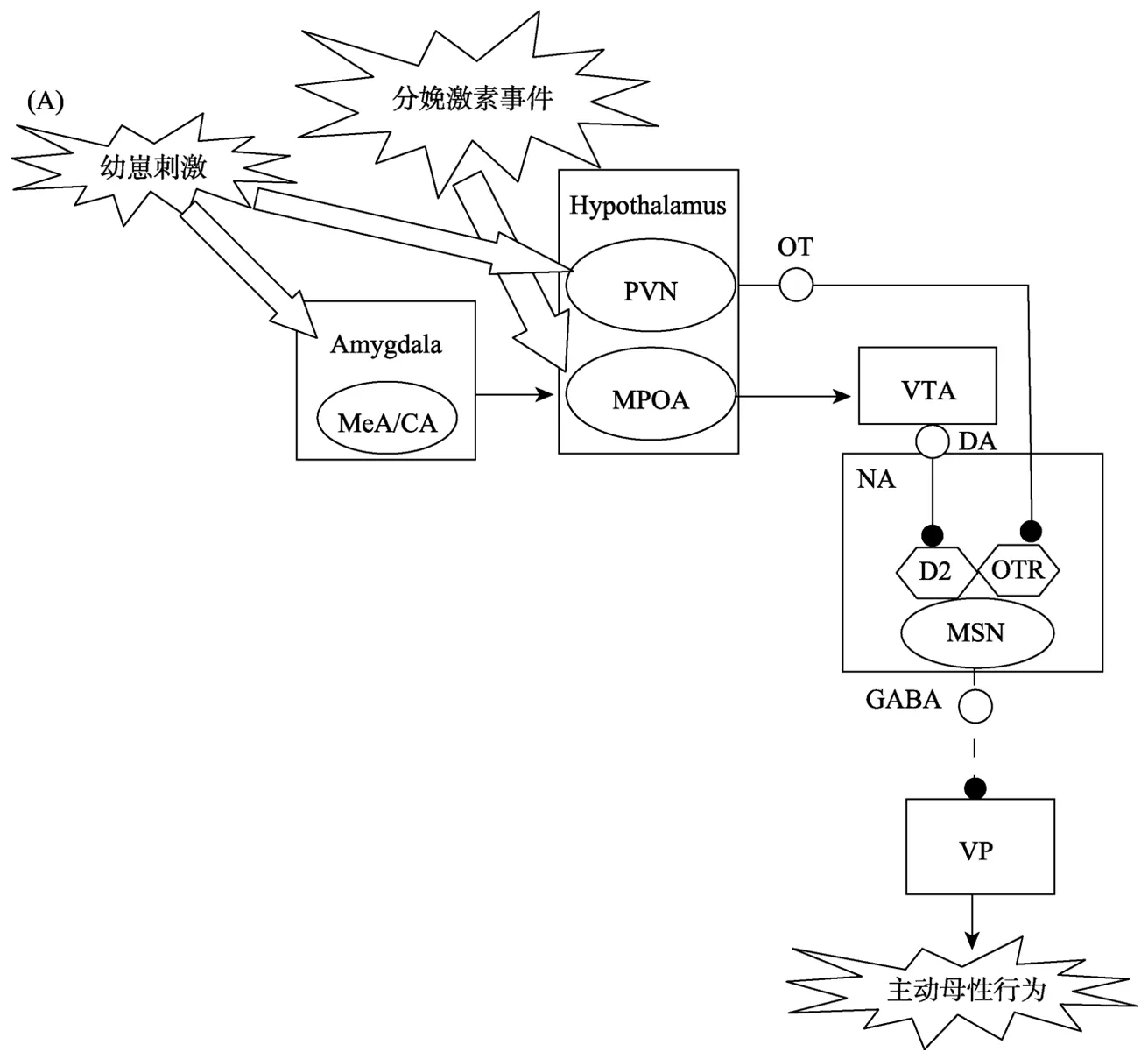

图1 主动母性行为动态改变的神经通路机制
MPOA的功能在产后晚期也发生了重大改变(Olazábal et al.,2013b)。研究(Pereira & Morrell,2009)发现,产后晚期 MPOA 失活使雌鼠对幼崽的衔回、舔舐和嗅探等行为的频率大幅提升,提示MPOA在产后晚期可能对主动母性行为起抑制作用。黑色素聚集素(melanin-concentrating hormone,MCH)参与了 MPOA对产后晚期主动母性行为的调节。研究(Benedetto et al.,2014)发现,向MPOA灌注 MCH能够损害主动母性行为,而产后晚期MPOA中 MCH 的水平不断上升,在断奶时达到顶峰,这与主动母性行为减少的趋势相一致。Pereira和Ferreira (2011)认为,产后晚期MPOA可能会抑制VTA中DA神经元的活动,从而降低幼崽的奖赏价值,也会使主动母性行为逐渐减少。
另外,MPFC的两个子区域在产后晚期对主动母性行为的调节作用与维持阶段有明显不同。证据表明,产后晚期IL对MPOA的兴奋性投射明显减弱(Olazábal et al.,2013b),IL的失活对主动母性行为不产生明显影响(Pereira & Morrell,2011;Olazábal et al.,2013b),提示IL可能不再参与主动母性行为的调节。证据(Pereira & Morrell,2009;2011)还表明,产后晚期PrL的失活会损害雌鼠主动母性行为的表现,而 MPOA失活(PrL完好)的雌鼠主动母性行为的表现则大幅提升,在受到幼崽抗拒时也不能对主动母性行为进行及时调整,提示PrL在产后晚期只以一种刻板的方式维系一定的主动母性行为。
可见,产后晚期主动母性行为的衰退是雌鼠对幼崽发展需要变化的一种响应。在这一过程中,BLA的情绪性学习效率下降对产后产晚期主动母性行为的衰退可能起主导作用。有研究者(Olazábal et al.,2013b)推测,产后晚期MPFC的功能改变可能与来自 BLA的情绪性学习信号减少有关,而MPFC对MPOA的兴奋性投射减弱又可能导致后者表现出抑制主动母性行为的作用。图1C说明主动母性行为衰退的神经通路机制。
4 总结与展望
综上所述,哺乳期啮齿动物的主动母性行为会经历从产后早期的发动和维持到晚期衰退的动态改变,涉及雌鼠调整自身的母性反应以适应幼崽奖赏价值阶段性变化的一系列复杂神经机制,与多条神经通路、多种心理成分和多个节点调节的神经加工有关。首先,MPOA-VTA-NA-VP是一条控制主动母性行为的基本通路。在主动母性行为发动阶段,分娩激素事件可以激活 MPOA,进而开启这一基本通路; 在维持和衰退阶段,来自BLA和MPFC的调控通路可以对基本通路进行实时调节,进而影响主动母性行为的表现。其次,以上通路机制包含多种成分的心理加工。在基本通路中,MPOA是加工母性动机和幼崽奖赏价值的中枢,VTA-NA-VP则是控制动机行为程序的一般通路。在调控通路中,BLA可以操作性地学习有关幼崽的情绪信息,使主动母性行为得以强化;MPFC则可以利用来自BLA的情绪性经验,通过奖赏评估调节雌鼠对幼崽的选择偏好,使主动母性行为在产后早晚期表现出不同的模式。最后,来自BLA和MPFC的调控通路可以通过多条神经投射对基本通路的多个节点进行调节。MPOA是基本通路的始端节点,其激活可以控制VTA释放DA,使雌鼠能够将幼崽作为高奖赏价值的目标;NA-VP则是基本通路的末端节点,NA中DA水平的提高可以使 VP去抑制,导致主动母性行为得以表现。在主动母性行为的维持阶段,BLA既可以通过对 NA的兴奋性投射易化通路末端 VP的去抑制,又可以直接向 VP发放兴奋性神经投射,这两条投射在基本通路末端的共同作用可以使主动母性行为得到有效强化; MPFC中的IL只兴奋基本通路始端的MPOA,而PrL则既可以兴奋始端的MPOA,也可以促进末端VP的去抑制,IL与PrL的协同作用使主动母性行为不仅具有稳定性,又有灵活性。在主动母性行为的衰退阶段,BLA对通路末端的去抑制和强化作用明显减退,通路始端 MPOA功能的转变使幼崽奖赏价值降低,MPFC对通路的调节作用也逐渐撤出,最终导致主动母性行为逐渐关闭。
然而,尽管调控哺乳期啮齿动物主动母性行为动态改变的神经机制已初见端倪,但仍有一些重要问题或方向亟待深入探究。
第一,MPOA中抑制性神经元的作用。MPOA包含背侧区(dorsal MPOA,MPOAd)和腹外侧区(ventrolateral MPOA,MPOAvl) (Olazábal et al.,2013b)。证据(Olazábal et al.,2013b)表明,MPOAd中分布着向脑内厌恶系统投射的抑制性神经元,MPOAvl中则分布着向 MPOAd投射的抑制性神经元。还有证据显示,前一类是协同表达甘丙肽(Galanin)的 GABA神经元,可以抑制雌鼠的主动回避幼崽行为(Wu,Autry,Bergan,Watabe-Uchida,& Dulac,2014; Zha & Xu,2015); 后一类则可能是协同表达MCH的GABA中间神经元,可以抑制MPOA对VTA的兴奋性传出,与产后晚期MPOA转变为抑制性角色有关(Benedetto et al.,2014;Olazábal et al.,2013b)。可见,这两类抑制性神经元可能通过不同的功能参与了产后早晚期主动母性行为的调节。进一步研究需要关注如下问题:(1) MPOAd中的GABA传出神经元如何与兴奋性传出神经元相互协同以调节产后早期主动母性行为?(2)产后晚期MPOAvl中的GABA中间神经元是否会通过抑制MPOAd中的GABA传出神经元使雌鼠主动回避幼崽?这在主动母性行为衰退的过程中又扮演怎样的角色?(3)产后不同阶段MPFC的功能改变是否也会影响MPOA中抑制性神经元的作用?
此外,脑内的五羟色胺(serotonin,5-HT)系统对MPOA的投射也可能参与主动母性行为的调控(陈磊磊等,2017),这是否会通过MPOA中的两类GABA 神经元起作用,尚待研究。
第二,NA不同子区域的作用。NA分为壳(shell)区和核(core)区。证据表明,电损毁壳区可破坏产后早期主动母性行为(Li & Fleming,2003),但采用布比卡因(bupivacaine)将 NA 整体失活却没有明显影响(Pereira & Morrell,2011),这提示NA 的壳区和核区的功能及其相互关系可能比较复杂。证据(Sesack & Grace,2010; Root,Melendez,Zaborszky,& Napier,2015)还表明,壳区和核区的MSN分别有投射到 VP的腹内侧区(ventromedial VP,VPvm)和背外侧区(dorsolateral VP,VPdl),其中,壳区到 VPvm的投射是主动母性行为去抑制的通路,而核区到VPdl的投射是调节自主运动去抑制的通路。NA整体失活导致的壳区受损可能使主动母性行为去抑制出现困难,而核区受损则会导致自主运动过度释放(Li & Fleming,2003;Pereira & Morrell,2011)。研究者(Nicola,2007; Pereira& Morrell,2011; Olazábal et al.,2013a)推测,较高的自主运动水平有利于主动母性行为的表现,从而在一定程度上弥补了壳区受损带来的损害,使行为的输出结果可能保持不变。值得注意的是,NA壳区受损妨碍主动母性行为的机制还不清楚。证据(Numan,2006)显示,NA壳区除了有一条对VPvm 的去抑制通路之外,还有一条表达脑啡肽(enkephalin)的神经通路可以易化 BLA的谷氨酸投射对 VP的直接激活作用,进而增进对主动母性行为的强化。未来研究可以通过 NA核区的单独损毁或失活探查自主运动如何参与产后早晚期主动母性行为的表现,还可以考察NA壳区向VP投射的两条通路如何在产后早晚期相互作用及其对主动母性行为动态改变的影响。
第三,MPFC中前扣带回(anterior cingulate,ACg)的调节功能。研究表明,在妊娠前损毁雌鼠MPFC中包含ACg在内的广泛区域,会使分娩后雌鼠衔回和舔舐幼崽的频率降低、秩序性受损(Afonso,Sison,Lovic,& Fleming,2007); 而单独失活 ACg主要损害主动母性行为的秩序性(Pereira & Morrell,2011)。可见,与PrL和IL不同,ACg可能主要通过调节多种行为成分的时间秩序来组织母性反应。有证据(Kesner,2000)表明,损毁ACg破坏了雌鼠对按时间顺序排列的空间线索记忆。而且,ACg与负责母性动机加工的MPOA和情绪性学习的BLA均有神经联系(Olazábal et al.,2013b)。今后的研究可以关注雌鼠在复杂的育幼环境中如何通过ACg的上述神经联系计划和组织适当的主动母性行为。
第四,幼崽声音信号识别与主动母性行为的动态改变。研究(Banerjee & Liu,2013; Marlin,Mitre,D’Amour,Chao,& Froemke,2015)表明,幼崽声音信号的改变不仅会引起雌鼠听觉皮层神经元电活动的可塑性变化,也使MPOA的激活水平发生了相应改变,提示听觉皮层对幼崽声音信号的识别可能参与了主动母性行为的调节。需要注意的是,来自幼崽的声音信号是一种间接的或远距离的刺激,雌鼠对其意义的识别可能涉及更复杂的认知加工过程,但幼崽声音信号调节主动母性行为动态改变的神经机制还不清楚。研究者(Banerjee & Liu,2013)推测,听觉皮层可以通过杏仁核(amygdala)和海马腹侧下托(ventral subiculum;vSub)与 MPOA 相互作用,完成对幼崽声音信号的识别及其情绪性和情境意义的学习与记忆。在此基础上,听觉皮层的神经信号不仅可以直接通过MPOA影响VTA-NA中的DA水平,还可以经MPFC到达 NA,调节主动母性行为的去抑制过程。未来研究可以建立产后早晚期的幼崽声学模型(acoustic model) (Banerjee & Liu,2013),系统考察主动母性行为从发动、维持到衰退的过程中听觉皮层神经可塑性变化与上述两条神经通路功能联结的形成与消退之间的关系,也可以比较幼崽听觉信号与其他感觉信号的识别在调节雌鼠主动母性行为动态改变中是否扮演不同角色。
最后,哺乳期人类母亲亲子互动行为的神经机制。尽管哺乳期的人类母亲并不表现出与啮齿动物主动母性行为成分(衔回、舔舐和嗅探等)完全对应的行为表现,但她们在照料婴儿过程中会利用多种感觉通道的信息和主动的行为反应与婴儿互动(包括相互凝视、身体接触和声音交流),以回应婴儿的需求(Strathearn,2011; Kim et al.,2016)。一些功能性磁共振成像(functional magnetic resonance imaging,fMRI)研究(Swain et al.,2008;Seifritz et al.,2003; Laurent & Ablow,2012)检查了人类母亲听到自己婴儿哭泣时的神经功能反应,发现纹状体(striatum)、下丘脑(hypothalamus)、杏仁核、MPFC和前扣带皮层(anterior cingulate cortex)等脑区的激活明显增强。这提示,母亲识别自己婴儿声音信号所激活的脑区与啮齿动物主动母性行为所涉及的脑区十分相似,也包含了负责动机、情绪和行为组织等多种心理加工成分的神经结构。但是,人类母亲与婴儿的亲子互动还可能涉及高级的认知执行功能。例如,亲子互动质量高的母亲在观看自己的婴儿玩耍视频时,与自我觉知和认知灵活性有关的额中回(middle frontal gyrus)和额下回(inferior frontal gyrus)的激活水平更高,且纹状体的 NA与这两个脑区的功能联结增强(Atzil,Hendler,& Feldman,2011; Lonstein et al.,2015),提示人类母亲的亲子互动依赖于皮层通过一系列高级功能与皮层下结构相互影响。另外,产后抑郁或有抑郁倾向的母亲上述脑区的神经活动与正常母亲有明显差异,如听到自己婴儿声音引起的NA激活低于正常母亲,而左侧MPFC的激活又高于正常母亲(Swain et al.,2008; Laurent& Ablow,2012),这也许可以为产后抑郁母亲的脑功能定向临床干预提供借鉴。然而,在 fMRI的实验中被试需要固定身体姿势,因此不可能考查真实的亲子互动过程中人类母亲的脑功能活动情况(Lonstein et al.,2015)。未来研究有必要引入可以允许母亲在亲子互动中完成某些身体动作的脑功能技术,如脑电图(electroencephalography,EEG)和功能性近红外光学成像(functional near-infrared spectroscopy,fNIRS)等,并将来自EEG、fNIRS和fMRI的研究证据相互参照,进一步探究哺乳期人类母亲亲子互动行为的神经机制(Kim et al.,2016)。
猜你喜欢
科学导报(2022年28期)2022-05-24
潍坊学院学报(2021年3期)2021-08-23
中国生殖健康(2020年2期)2020-12-09
做人与处世(2020年10期)2020-06-29
发明与创新·大科技(2019年6期)2019-09-06
江西社会科学(2018年8期)2018-08-29
科技创新与品牌(2018年5期)2018-07-24
电影新作(2014年1期)2014-02-27
中国乡村医药(2011年10期)2011-08-30
中国循证儿科杂志(2011年6期)2011-01-19
