多效唑对水分胁迫下苹果砧木八棱海棠光合及抗氧化酶活性等生理特性的影响
2018-08-08 02:21朱燕芳王延秀贾旭梅郭爱霞
干旱地区农业研究 2018年4期
朱燕芳,王延秀,胡 亚,贾旭梅,郭爱霞
(甘肃农业大学 园艺学院,甘肃 兰州 730070 )
西北黄土高原地区海拔高、光照充足、昼夜温差大、生产集中,是我国苹果的最优生态区,但干旱已成为制约该地区苹果生长发育的重要因子[1-2]。八棱海棠(MalusrobustaRehd.)因其抗寒抗旱、耐盐碱、与栽培品种亲和力好等优点成为北方苹果产区的常用砧木[3-4]。 利用抗性砧木及应用外源物质提高苹果属植物水分利用效率,是解决苹果生产中干旱缺水问题的主要方法之一[5]。
多效唑(PBZ)是一种三唑类抑制内源赤霉素合成的高效低毒植物生长调节剂,能够延缓植物生长,同时增强植物抗逆性[6]。有研究表明,PBZ对飞燕草(Consolidaorientalis)的花期、叶色、花色等有一定的影响[7],且PBZ能延迟番茄中灰霉菌的生长,增强抗性[8]。玉米、马铃薯等作物中,应用PBZ可降低叶片气孔开度、增加气孔阻力,减少植物体的蒸腾失水而提高植物叶片的相对含水量和抗旱性[9-10]。在果树上应用PBZ的研究多集中于水分生理研究,韩振海等[11]认为叶面喷施PBZ,可明显减少水分胁迫下果树叶片内游离脯氨酸的含量,增加叶片水势,提高叶片的抗逆性。关于苹果砧木在水分胁迫下,PBZ对光合参数和叶绿素荧光参数影响的研究报道较少。本试验探讨了水分胁迫下PBZ对八棱海棠光合特性、叶绿素荧光参数、抗氧化酶、膜脂过氧化、渗透调节物质含量的影响,以期明确PBZ提高植物耐旱性机理,同时为苹果栽培中应用外源物质提高抗旱性提供参考。
1 材料与方法
1.1 材料及处理
试验于2015年3-9月在甘肃农业大学园艺学院避雨棚进行。 2015年2月1日选取饱满一致的八棱海棠种子,蒸馏水浸泡12 h后,在4℃进行湿沙处理。3月18日选露白较好、发芽一致的种子播入蛭石中,室温下培养,待幼苗长至2片真叶时,移入直径为15 cm、装200 g营养土(草炭∶珍珠岩∶蛭石=3∶1∶1按体积比混合)的花盆中,每盆一株,统一管理。在育苗期间定期浇水、松土、除草。植株具10片真叶时,进行盆栽控水胁迫处理。采用持续称重法控制土壤含水量,每2天于傍晚称重并补充水分至最大田间持水量的45%~55%(质量含水量)。对照(CK)处理进行持续控水胁迫,不喷施任何溶液,试验设5个处理,分别用0 mg·L-1(C0,水溶液不含PBZ)、50 mg·L-1(C1)、100 mg·L-1(C2)、150 mg·L-1(C3)、200 mg·L-1(C4)的PBZ溶液进行喷施处理至滴水为止。每个处理30盆,重复3次,每3天取样并测定相应的指标。
1.2 试验方法
1.2.1 光合作用气体交换参数的测定 从胁迫处理当天开始,之后每隔3天,上午8∶00,用光合仪(Li-6400,LI-COR公司,美国)测定生长一致的幼苗第8片真叶的净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、胞间CO2浓度(Ci)。设定CO2浓度为400 μmol·mol-1,环境温度25℃,光强为800 μmol·m-2·s-1。
1.2.2 光合作用荧光参数的测定 采用光合仪Li-6400测定荧光参数,叶片暗适应后,设置测量光饱和后夹好叶片,等待dF/dt绝对值<5时记录数据,计算非光化学猝灭系数(NPQ)、PSⅡ的最大光化学效率(Fv/Fm)、PSII的实际光化学效率(ΦPSⅡ)、光化学猝灭系数(qP)等光合与荧光参数;测定重复5次。
1.2.3 生理生化指标的测定 SOD、POD、CAT活性采用李合生[12]的方法测定;电解质渗漏率采用电导率法测定,MDA 含量采用硫代巴比妥酸法测定[12];脯氨酸含量采用磺基水杨酸法测定[13]。
1.3 数据分析
试验数据应用Origin 9.0软件作图,运用SPSS 22.0软件进行数据差异显著性分析和主成分分析。统计分析采用单因素ANOVA的LSD比较差异的显著水平(α= 0.05)。在主成分分析前,用隶属函数法对数据进行转化。
采用模糊数学隶属函数法对各指标第15天数据进行分析,隶属函数的计算公式[14-15]:
(1)
反隶属函数值:
(2)
式中,Xi为指标测定值,Xmin、Xmax为所有参试材料某一指标的最小值和最大值,如果(Xi-Xmin)为负值,则采用(2)式。
2 结果与分析
2.1 外源多效唑处理对水分胁迫下八棱海棠叶片光合特性的影响
如图1所示, 随着水分胁迫的持续八棱海棠叶片的Pn、Gs、Tr呈显著下降的趋势,而Ci呈上升趋势。随着胁迫时间的延长,对照组Pn下降程度较为明显;处理15 d后,CK及C0、C1、C2、C3、C4比第0天,分别下降53.72%、50.00%、35.71%、22.28%、32.78%、31.52%,各处理组Pn下降幅度显著低于对照,C0组与CK的下降幅度基本一致,其中C2(100 mg·L-1PBZ)处理显著低于其他处理及对照(图1A)。Gs、Tr的变化与Pn变化相似,水分胁迫15 d后CK及C0、C1、C2、C3、C4的Gs分别下降了60.71%、60.00%、40.06%、36.47%、45.24%、42.68%(图1B),Tr分别下降了57.33%、53.42%、41.66%、33.78%、39.43%、43.84%,其中C2处理后的两指标下降幅度均显著低于对照及其他处理(图1C)。处理15 d后,CK及各处理的Ci分别上升了37.12%、37.36%、34.24%、26.98%、36.08%、29.28%,各处理组上升幅度均小于对照组,其中处理组C2上升幅度最小,且显著低于对照及其他处理组(图1D)。
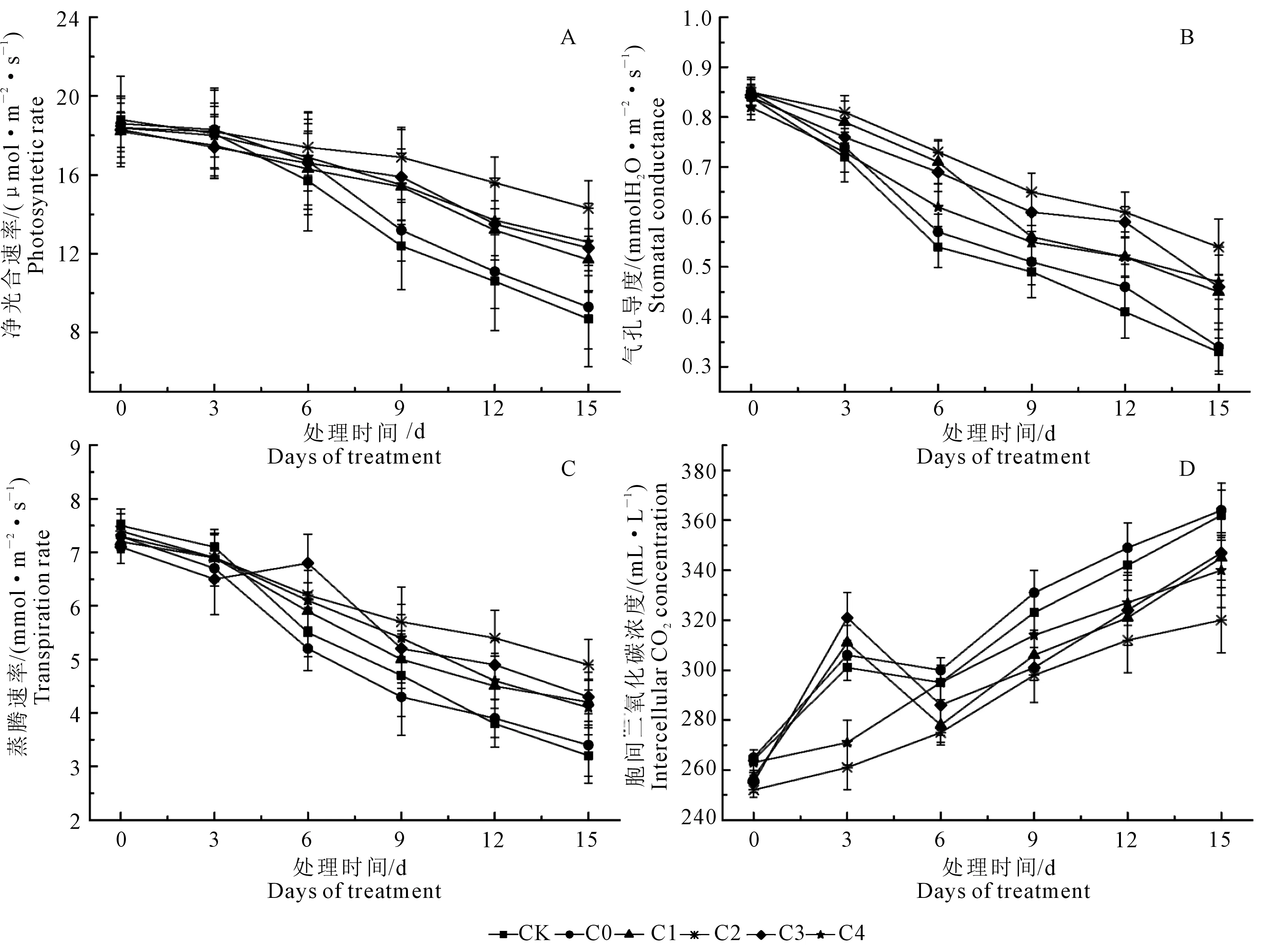
图1 外源多效唑处理对水分胁迫下八棱海棠光合特性的影响Fig.1 Effect of exogenous PBZ treatment on photosynthetic characteristics on Malus robusta Rehd. under water stress
2.2 外源多效唑处理对水分胁迫下八棱海棠荧光参数的影响
八棱海棠叶片的Fv/Fm呈缓慢下降的趋势,说明植株遭受光抑制,CK和C0持续下降,而C1、C2、C3、C4处理组变化较为平缓,表明PBZ处理能提高植株的抗旱能力(图2A)。随着水分胁迫时间的延长,为维持光系统的稳定,各处理组NPQ呈上升的趋势, 在9~15 d,CK和C0的NPQ明显高于其他处理组,与第0天相比,在第15天对照组及各处理组NPQ升高400.00%、348.28%、212.90%、153.13%、292.86%、266.67%,各处理组上升幅度低于CK(图2B)。各处理组qP均呈下降趋势,C2组下降程度最小,为44.62%,说明植株吸收的光能较多地用于光合作用,从而避免光合机构的损坏(图2C)。 各处理组的ΦPSⅡ在0~9 d,CK及各处理后变化一致,但在9~15 d,各处理组的数值均高于CK。在处理第15天,CK、C0、C1、C2、C3、C4分别比第0天下降51.22%、48.19%、35.00%、30.33%、41.25%、40.74%,说明PBZ处理后能提高PSII光合作用反应中心实际光能捕捉效率(图2D)。

图2 外源多效唑处理对水分胁迫下八棱海棠荧光参数的影响Fig.2 Effect of exogenous PBZ treatment on fluorescence parameters on Malus robusta Rehd. under water stress
2.3 外源多效唑处理对水分胁迫下砧木抗氧化酶活性的影响
由图3A可知,随着水分胁迫时间的延长,八棱海棠叶片的POD酶活性逐渐升高,CAT活性和SOD活性呈下降趋势。与第0天相比,处理15 d后,CK及各处理组分别上升141.94%、132.26%、84.85%、75.00%、73.53%、93.75%,且处理组的降低幅度低于对照组。CK、C0与C1的SOD活性呈现显著下降;C2在0~3 d短暂的升高,3~9 d持续下降,9~12 d则呈上升,12~15 d显著下降,C3在0~6 d持续上升,之后显著下降,C4在0~3 d有短暂升高,之后持续下降。与第0天相比,处理15 d后,CK与各处理组分别下降了40.71%、39.36%、28.94%、21.66%、30.35%、31.96%,其中C2的下降幅度均低于对照及其他处理组(图3B)。相比SOD,CAT活性下降呈现不同趋势。随着胁迫时间的延长,C2的CAT显著高于CK及其他处理;CK组的CAT活性在0~3 d显著下降,3~6 d呈缓慢上升趋势,6~15 d又呈显著下降的趋势,而C1、C2、C3、C4均呈缓慢下降趋势。与第0天相比,处理15 d后,CK和处理组的CAT活性分别下降了52.31%、48.06%、30.33%、31.34%、35.20%、34.65%,处理组与对照组之间存在显著差异(图3C)。
2.4 外源多效唑处理对水分胁迫下八棱海棠渗透胁迫物质含量的影响
图4显示,随着水分胁迫的持续,八棱海棠叶片Pro、REC、MDA总体呈升高趋势。在图4A中,C2组MDA含量显著低于CK和其他处理组,CK和C2在前6天呈上升趋势,6~9 d有短暂的下降,之后持续上升,C1和C4在前9 d缓慢上升,9~12 d下降,之后升高,C3持续缓慢升高,在处理15 d,CK是C2的1.27倍。与第0天相比,CK及各处理组分别上升80.77%、60.71%、56.60%、37.04%、56.36%、66.67%。胁迫3~15 d,CK组的Pro显著高于其他处理组,而C2显著低于对照和其他处理组,在处理15 d,CK是C2的1.63倍,与第0天相比,CK及各处理组分别上升312.50%、246.67%、226.83%、129.55%、166.67%、189.58%(图4B)。REC的变化与Pro一致,即C2显著低于CK和其他处理组,在处理15 d,CK是C2的1.42倍,与第0天相比,CK及各处理组分别上升142.86%、113.33%、85.91%、53.85%、93.15%、81.58%(图4C)。由此可知,八棱海棠Pro、REC、MDA的各个处理组的上升幅度明显低于对照,其中C2上升幅度最小。说明PBZ处理能有效缓解水分胁迫下渗透胁迫物质的上升。

图3 外源多效唑处理对水分胁迫下八棱海棠抗氧化酶活性的影响Fig. 3 Effect of exogenous PBZ treatment on antioxidant enzyme activity on Malus robusta Rehd. under water stress
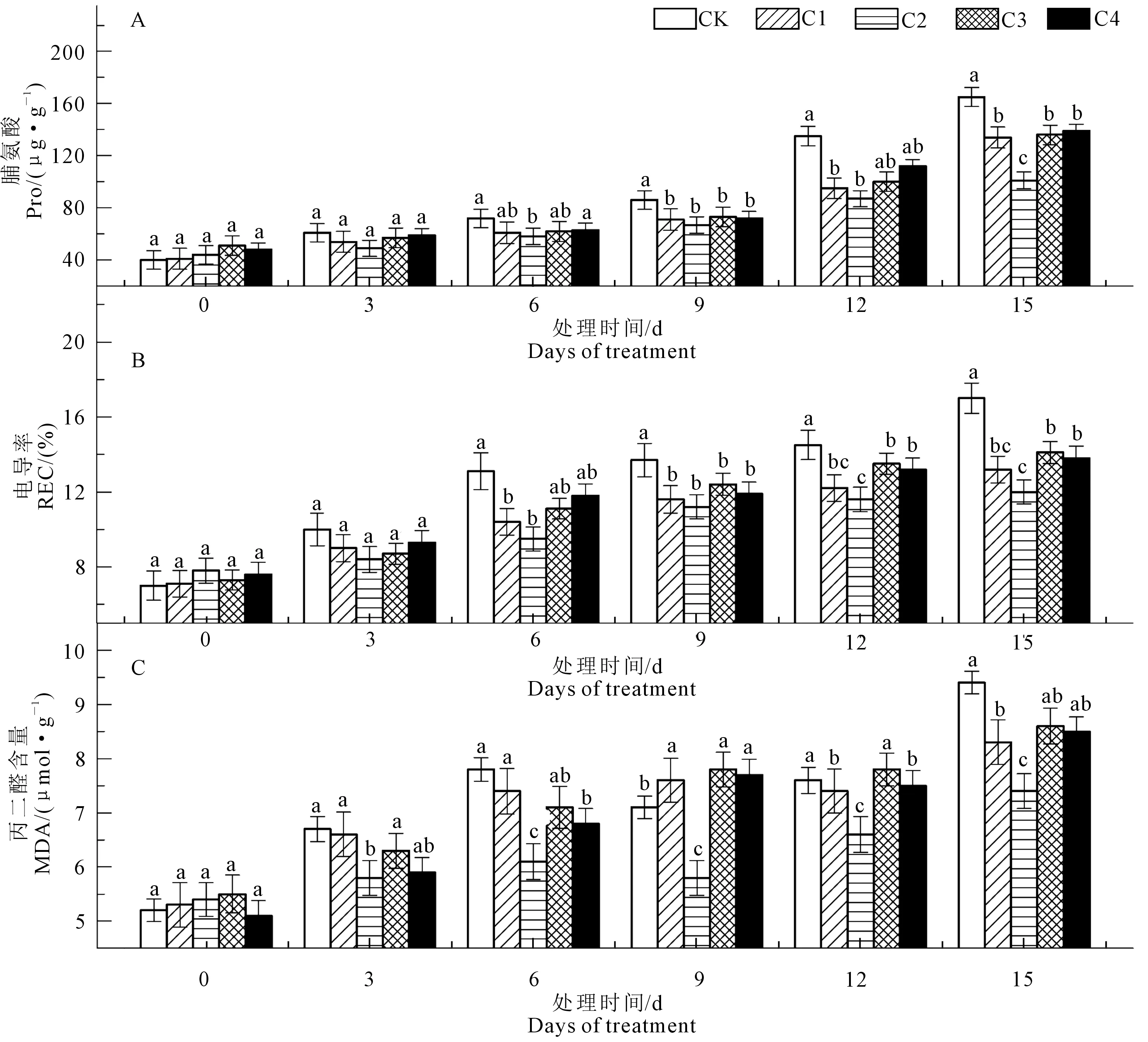
图4 外源多效唑处理对水分胁迫下八棱海棠脯氨酸、电导率、丙二醛含量的影响Fig. 4 Effect of exogenous PBZ treatment on proline, relative electrical conductivity, malondialdehyde on Malus robusta Rehd. under water stress
2.5 不同浓度多效唑处理下八棱海棠抗旱能力的综合评价
2.5.1 利用模糊隶属函数分析方法对不同浓度多效唑处理下八棱海棠抗旱能力的综合评价 为了克服单个指标的局限性,全面反映外源多效唑对八棱海棠幼苗的抗旱性影响,对各项指标进行隶属函数分析。根据其隶属函数平均值作为抗旱能力鉴定的标准进行排序。由表1可知,C2(100 mg·L-1)处理下八棱海棠幼苗抗旱性最强。

表1 利用模糊隶属函数法对不同浓度多效唑处理下八棱海棠幼苗抗旱能力的综合评价Table 1 Comprehensive evaluation of drought resistance of Malus robusta Rehd. under different paclobutrazol concentration treatments using fuzzy membership analysis
2.5.2 利用主成分分析方法对不同浓度多效唑处理下八棱海棠抗旱能力的综合评价 水分胁迫下,不同浓度PBZ处理,对八棱海棠幼苗叶片14个指标的数据经隶属函数转化后进行主成分分析(图5、表2~4)。由表2和图5可知,从14个主成分中提取特征值大于1的两个主成分,其累积贡献率达93.869%,能够较好地代表数据所反映的信息。由表3 可以看出,第一主成分对 14 个变量指标的信息提取充分。将上述 2 个主成分的得分值代入综合评价函数,计算不同浓度PBZ处理的综合得分,综合指数的得分越高,说明该处理能有效提高八棱海棠抗旱性。从表4可知,C2浓度处理下,八棱海棠幼苗抗旱性最强,这与模糊隶属函数分析法结果一致。

图5 主成分分析碎石图Fig. 5 The principal component analysis scree plot

表2 主成分列表及方差贡献率Table 2 List of principle components, percentage of variance,cumulative percentage
3 讨 论
逆境下植物光合速率下降,同化产物减少[16-17]。干旱胁迫对植物光合作用的抑制存在气孔限制和非气孔限制两种方式[18]。只有当Ci降低和气孔限制值增大时,光合速率降低是由于气孔导度降低所引起的。反之,当叶片光合速率降低且Ci浓度升高,则光合作用的主要限制因素是非气孔因素[19-20]。本试验表明不同浓度PBZ处理后,随着水分胁迫时间的延长,八棱海棠Pn、Gs和Tr均呈下降趋势,而Ci浓度总体呈上升趋势,说明光合作用下降主要受非气孔因素的影响,与牛铁泉等在苹果幼苗的研究结果一致[21]。各处理组Pn、Gs、Tr的下降幅度显著低于对照组,同时Ci的上升幅度均小于对照组,说明PBZ有效缓解了Pn、Gs、Tr的降低,以及Ci的升高,此结论与冯立娟等在大丽花中的研究结果一致[22],同时PBZ可能是通过调节植株光合系统,减缓各光合指标在逆境下的受抑制程度,使植株能更好适应逆境胁迫,以此来增强植株的抗旱性。
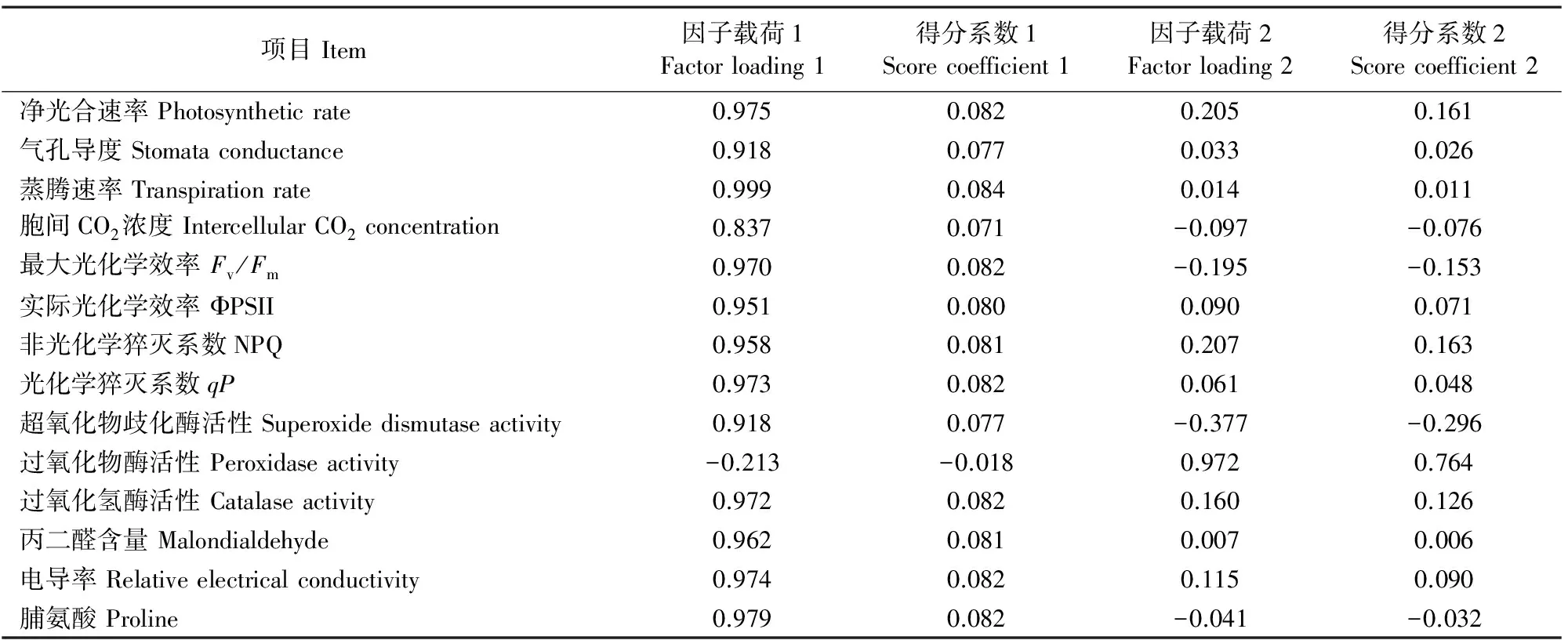
表3 因子负荷矩阵和得分系数矩阵Table 3 Component matrix and score coefficient matrix

表4 利用主成分分析法对不同浓度多效唑处理下八棱海棠幼苗抗旱能力的综合评价Table 4 Comprehensive evaluation of drought resistance of Malusrobusta Rehd. under different paclobutrazol concenreation treatments using principal component analysis
前人研究表明叶绿素荧光参数会响应非气孔限制[23]。本试验中,八棱海棠叶片的Fv/Fm、qP、ΦPSⅡ均降低,表明随着水分胁迫时间的延长,植物色素可能受到了不同程度的破坏,使得电子转换效率下降,同时植物吸收的光能用于光化学反应的比例下降,而NPQ显著上升,则表明植物散热能力增强,是植物自我保护的一种机制,这与Mohammed Mouradi 等在紫花苜蓿中研究的结果类似[24]。PBZ处理后其Fv/Fm、qP、ΦPSⅡ较CK明显提高,各处理组的下降幅度显著低于CK,同时NPQ的上升幅度低于CK,表明PBZ缓解了叶绿素荧光参数的降低幅度,减轻其胁迫损伤程度,同时提高了砧木对胁迫的适应能力,这一结果与余凯凯等在马铃薯中的研究结果一致[25]。Farzad等也证实qP下降幅度越小,表明植物越抗旱[26]。其中C2处理组,即100 mg·L-1PBZ,叶绿素荧光参数的下降和上升幅度最小,也可反映此浓度下,能及时耗散光合系统中的过剩光能,从而保护PSⅡ反应中心免受因吸收过多光能而引起的光氧化伤害[27]。
超氧化物歧化酶(SOD)是一种清除超氧阴离子自由基的酶,它催化超氧阴离子自由基与氢离子反应产生H2O2和O2,H2O2可由过氧化氢酶进一步分解或被过氧化物酶利用[28]。POD和CAT是植物体中最重要的消除自由基的酶,POD 和CAT活性高低可反映出植物对干旱的抵御能力大小[29]。本试验中随着水分胁迫时间的延长,各处理组CAT和SOD总体呈下降趋势,说明水分胁迫下八棱海棠活性酶系统遭到破坏。而POD酶活性则逐渐升高,表明PBZ处理提高了八棱海棠幼苗的活性氧清除能力。相对而言,PBZ处理以后SOD、CAT下降幅度及POD上升幅度均低于对照组,说明PBZ处理在很大程度上缓解了水分胁迫对八棱海棠抗氧化系统的损伤。
水分胁迫下,植物膜系统遭到破坏,膜内大量的物质外渗,导致植物相对电导率的急剧上升。因此,电导率作为膜透性高低的指标,可用来描述植物抗旱性的强弱[30]。本研究表明,水分胁迫下八棱海棠叶片相对电导率大幅上升,经PBZ处理的植株其相对电导率上升幅度显著低于对照组,表明PBZ处理能够有效缓解水分胁迫对膜系统造成的破坏,这与Paul等研究结果一致[31]。游离脯氨酸是一种渗透调节物质,干旱胁迫条件下植物体内会大量积累游离脯氨酸[32]。本试验中,各个处理组Pro随处理时间均逐渐升高,说明脯氨酸的积累是干旱胁迫引起的应答反应[33],MDA与Pro的变化趋势一致,说明干旱胁迫下膜系统有一定程度的破坏[34]。但PBZ处理后的REC、Pro和MDA的上升幅度总体显著低于对照,表明外源PBZ处理能提高或增强相关抗逆生理基础,即提高了渗透调节能力。其中100 mg·L-1PBZ处理下三个指标的上升幅度最小,说明此浓度下对于渗透胁迫物质的上升效果最佳。
通过比较不同指标的变化,反映不同浓度的PBZ对于植株抗旱性的提高,同时本试验选用模糊隶属函数分析法和主成分分析法对不同浓度PBZ处理下八棱海棠幼苗抗旱能力进行更进一步的综合评价[35]。由于不同指标对评价体系的正负影响,采用隶属函数和反隶属函数法对各指标数据转化后进行主成分分析,使结果更为客观合理[36]。由两种方法综合分析排序可知,C2(100 mg·L-1)处理浓度最能有效提高八棱海棠的抗旱性,但C3(150 mg·L-1)和C4(200 mg·L-1)两处理浓度排序则相反,可能是两个浓度处理后,由于PBZ浓度偏高,浓度效应不明显。本试验中虽选出了PBZ最佳浓度,但PBZ从更深层次对植株的调节机理尚不明确,今后应从基因、蛋白、代谢等各个方面更透彻了解PBZ提高植株抗旱性的机理,以明确PBZ是通过调节植株光合和抗氧化酶系统中哪些相关的基因和蛋白以及代谢产物,为干旱地区果树栽培提供更有力的理论依据。
猜你喜欢
作物研究(2022年4期)2022-09-21
舰船科学技术(2022年11期)2022-07-15
海洋通报(2021年5期)2021-12-21
今日农业(2021年13期)2021-11-26
农业资源与环境学报(2021年5期)2021-10-06
中国糖料(2021年3期)2021-07-13
湖北农业科学(2020年22期)2020-12-18
种子(2020年11期)2020-12-05
上海农业学报(2018年5期)2018-10-30
现代园艺(2018年3期)2018-02-10