陇东黄土高原地区杏园土壤综合肥力对人工生草模式的响应及其环境解释
2018-08-08 02:21王根旺
干旱地区农业研究 2018年4期
王根旺, 宋 曦
(1.甘肃省高校陇东生物资源保护与利用省级重点实验室,甘肃 庆阳 745000;2.陇东学院生命科学与技术学院,甘肃 庆阳 745000; 3.陇东学院农林科技学院,甘肃 庆阳 745000)
目前,作为生态农业建设重要组成部分的果园生草管理模式,以其绿色环保和可持续发展等特点已逐步成为我国果园建设的主流模式[1],在推动区域经济发展、减少面源污染和农民增收等方面做出了重要贡献[2]。但不同生草模式下果园土壤综合肥力差异是制约我国果业可持续发展的关键问题[3],同时也是我国现代化果业建设中亟待解决的问题之一。果园土壤综合肥力差异通常与生草品种、管理模式、气候条件、土壤养分、土壤类型和生草年限有关[4],但人工构建的生草管理模式对果园土壤养分互作与竞争[5]、吸收与转化[6]以及土壤综合肥力[3]的影响机制尚不清楚,极大地制约了果园生草复合系统中果树与生草间的养分互作与驱动机制的理解,最终影响果园生草复合系统土壤综合肥力的提升和可持续经营。
陇东黄土高原地区生态脆弱,土壤瘠薄,气候干旱少雨,寒暑及昼夜温差极大,植被分布因而有其独特的地域性[7]。杏树(PrunusarmeniacaL.)是我国原生经济树种,因其抗逆性强,生长迅速、根系发达等优点成为陇东黄土高原地区的主要造林树种和经济林种[8]。为了落实国家退耕还林还草和生态环境重建政策,2001年庆阳市着力推广建设经济林生草复合管理模式,为农民增收的同时又为该地区生态环境的改善做出重要贡献。但近年来黄土高原地区果园生草复合管理模式的研究集中在苹果园土壤微生物多样性[9]及其群落结构[10]、土壤酶活性及理化性质[5]和果实品质[11]等方面,而有关黄土高原地区杏树林人工生草管理模式相关研究以及有关影响该地区人工生草管理模式杏树林土壤综合肥力的关键环境因子的研究则鲜见报道。
为此,本研究以陇东黄土高原地区树龄16 a并在2011年起采用不同生草管理模式的杏树林根际土壤为研究对象,采用常规方法测定了土壤理化性质、酶活性及土壤微生物群落遗传多样性等14项环境因子,利用非度量多维尺度(NMDS)排序和方差分解等多元分析法,系统分析了不同生草模式下杏树样地与土壤环境因子间的关系,以及环境因子在驱动不同生草模式样地分异过程中的相对重要性,初步确定了导致不同生草模式下杏树林土壤综合肥力出现差异的关键环境因子,旨在为今后陇东地区人工生草模式杏树林土壤综合肥力的提升、科学管理及可持续发展提供科学依据和数据参考。
1 试验地概况与研究方法
1.1 试验地概况
本研究在陇东学院甘肃省高校陇东生物资源保护与利用省级重点实验室生态农业试验站进行,该区位于陇东黄土高原庆阳市镇原县太平乡慕坪村西坡组,土壤类型为黄绵土,质地为沙土。地处东经107°38′-108°40′,北纬35°69′-35°71′,海拔1 380~1 420 m。上述区域属黄土高原沟壑地貌,是典型水土流失区之一,侵蚀面积25 050.5 km2,占全区总面积的92.4%,侵蚀模数6 383~9 000 t·hm-2。属暖温带半干旱季风气候区,光、热等气候资源丰富,作物种植制度为旱地一年一熟。由于季风的强烈影响,冬季寒冷干燥,春季多风且干旱少雨,夏季温暖多雨,水热同期,年降水量400~600 mm,雨量分布不均,且主要集中在7~9月,多年平均蒸发量1 000 mm以上,地面蒸发量为350~560 mm,地下水位深,土壤瘠薄,水土流失严重[8]。
1.2 研究方法
1.2.1 试验设计及土样采集 试验于2016年5月进行,选择试验站内树龄16 a并在2011年起采用不同生草管理模式的杏树林根际土壤为研究对象(A:杏树+黄花菜;B:杏树+紫花苜蓿;C:杏树+黄花菜+紫花苜蓿;以杏树+清耕作为对照组CK),试验区面积为20 m×20 m=400 m2、阳坡(坡度2°~5°)、株距200 cm,行距150~200 cm,供试样树概况见表1。按对角线五点取样法采集土样,先拨去土壤表层的浮土(约5~8 cm)及石块、落叶等杂质,用取土钻在每一样地中采集0~40 cm深度的土壤,将5个点采集的土壤混匀成1个土样,每个样地取3个重复,所采土样放在冷藏箱中及时带回实验室根土分离,除利用鲜土提取土壤微生物细菌基因组总DNA外,其余土样在室温背阴处风干,过2 mm筛备用[7]。测定土壤养分所需的样品采用“四分法”取样,全部通过1 mm孔径筛,再从过1 mm孔径筛的土样中均匀取出1/4,过0.149 mm孔径筛[8]。
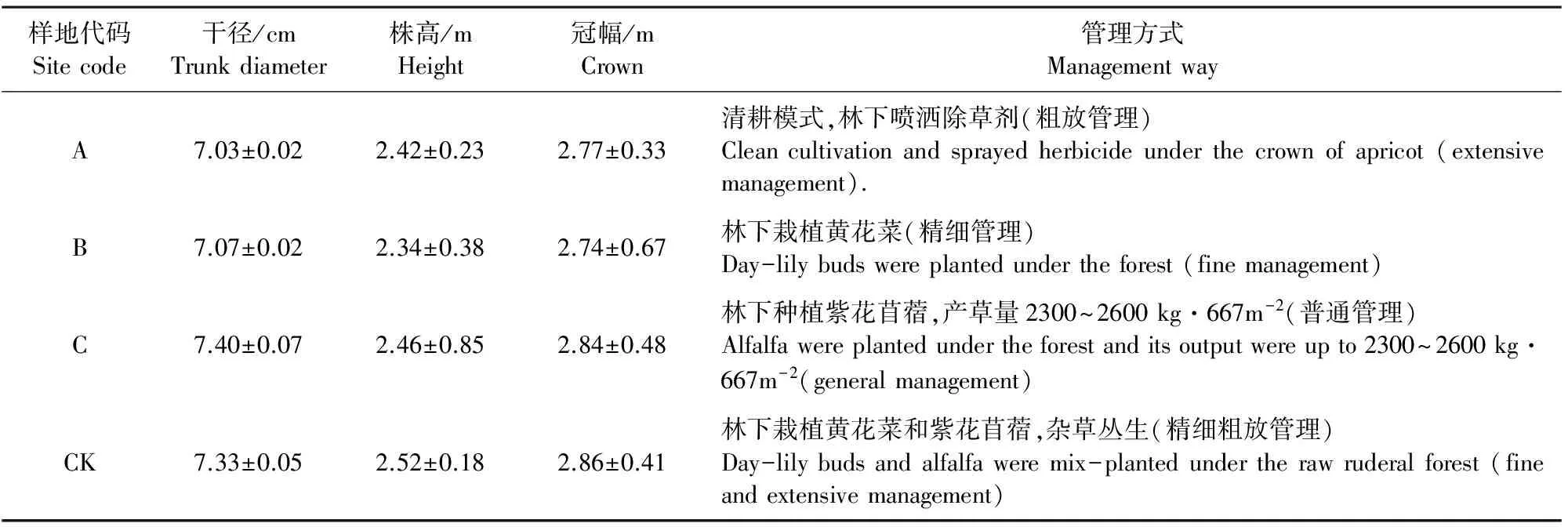
表1 供试样树概况Table 1 Basic parameters of the sample tree in the studied sites
1.2.2 土壤酶活性、理化性质及土壤微生物群落遗传多样性测定 土壤有机质和碱解氮分别采用重铬酸钾容量法-外加热法和碱解扩散法[5]测定;土壤速效磷、速效钾分别采用0.5 mol/LNaHCO3浸提-钼锑抗比色法和火焰光度计法[8]测定;电位法(pHS-3酸度计)测定土样pH值;转化酶、脲酶、碱性磷酸酶和过氧化氢酶分别采用3,5-二硝基水杨酸比色法、靛酚蓝比色法、磷酸苯二钠比色法和容量法[11]测定。采用变性凝胶梯度电泳(PCR-DGGE)技术分析微生物群落遗传多样性[7]。
1.2.3 土壤微生物群落遗传多样性分析 采用变性凝胶梯度电泳(DGGE)技术分析微生物群落遗传多样性,利用4种多样性指标进行土壤微生物群落遗传多样性分析,即Patrick丰富度指数(S):S=物种数,即条带数;Shannon-Wiener多样性指数(H):H=-∑PilnPi,Pi代表土壤样品中第i个条带的吸光度占所有条带吸光度总和(N)的比例;Margalef丰富度指数(D):D=(S-1)/lnN,S为每条泳道的总条带数[3];Pielou均匀度指数(J):J=H/Hmax,Hmax代表最大多样性值(Hmax=lnS)[7]。
1.2.4 数据处理 利用SPSS 16.0和R2.15.2(http:∥www.r-project.org/)对数据进行处理分析,用Origin 8.0和R 2.15.2做图。为了阐述供试样地与土壤环境因子间关系,基于Bray-Curtis相异指数,使用R语言“vegan”程序包中“envfit”程序,将所测定的14项土壤环境因子拟合到4个供试样地的非度量多维尺度(NMDS)排序图上,为防止样点在各象限的分布发生移位,在拟合时对NMDS的排序轴进行了旋转,以确保NMDS第一轴能够最大程度地代表群落相异性的变异。4个供试样地矩阵及环境因子数据集分别用Bray-Curtis与Euclidean距离表示[12]。方差分解运用“Vegan”数据包里的“Varpart”功能对供试样地中14个环境因子进行分析,所有数据均经过Hellinger转换,用Venn图表示土壤理化性质、酶活性及土壤微生物群落对供试杏树根际土壤综合肥力的纯效应以及共同效应,图中的数字代表上述环境因子对供试杏树根际土壤综合肥力的解释率[13]。
2 结果与分析
2.1 杏树林根际土壤酶活性及理化性质对不同生草管理模式的响应
土壤酶催化土壤生态系统中生物化学反应,参与物质循环和能量转换,其活性大小是表征土壤肥力的重要指标。由表2和表3可见,不同生草模式对陇东黄土高原地区杏树林根际土壤酶活性及理化环境因子产生了显著影响(P<0.01),表现为3种生草模式的供试土壤酶活性均明显高于CK组(P<0.05),但各环境因子对不同生草模式的响应则不尽相同。其中土壤转化酶活性在模式A和B间无显著差异,均低于模式C(P<0.05);模式B和C的碱性磷酸酶活性低于模式A(P<0.05),而其脲酶活性则高于模式A(P<0.05);而过氧化氢酶活性在模式A、B和C间差异则未达显著水平。
吴玉森等[1]研究了多年自然生草对黄河三角洲梨园土壤酶活性的影响,结果表明自然生草7 a梨园的土壤脲酶和碱性磷酸酶活性分别是对照的3.8倍和1.5倍;郑海金等[14]分析了农林复合经营模式对红壤丘陵区侵蚀坡地土壤酶活性的影响,结果显示原侵蚀裸露荒坡地建立农林复合系统可提高土壤转化酶、脲酶和磷酸酶活性,其中农-林和纯林经营系统能显著提升土壤转化酶和脲酶活性,但农-林-草系统却降低了脲酶活性。上述结果与本研究结果不尽相同。究其原因可能是由于,首先土壤脲酶是一种作用于尿素肥料的中性水解酶类,可加速土壤中潜在养分有效化,使土壤中含氮有机化合物酰胺肽键水解生成的氨是植物氮素营养来源之一[15]。如表2所示,模式B和C的脲酶活性明显高于模式A(P<0.05),说明杏树林嵌套种植紫花苜蓿可有效增加土壤氮素循环,提高了土壤养分有效化水平。其次,土壤转化酶活性在模式A和B间无显著差异,均低于模式C(P<0.05),转化酶参与土壤有机碳循环,将蔗糖分解成葡萄糖和果糖的同时为土壤微生物提供大量的速效碳源,通过刺激土壤微生物的生长繁殖,加速土壤营养物质的有效转换[16]。由表2可见,模式A和B转化酶活性均明显低于模式C,说明多种经济植物嵌套种植的人工生草模式可有效增加土壤微生物群落多样性,体现了地表植被多样性与地下土壤微生物群落遗传多样性协同变化的特性[17],进而刺激土壤转化酶活性,加速了土壤有机碳的地球生物化学循环速率,进而提高土壤养分有效化水平。第三,土壤过氧化氢酶广泛存在于微生物和植物的细胞中,可清除由于生物呼吸过程中和有机物生物氧化过程中产生的过氧化氢[7]。虽然本研究中土壤过氧化氢酶活性在模式A、B和C间差异未达显著水平,但均高于CK组,说明采取复合生草模式可明显降低土壤过氧化氢的积累,进而减少其对植物及土壤微生物的毒害作用。第四,碱性磷酸酶活性在模式B和C无显著性差异,但均低于模式A,说明种植黄花菜的生草模式可有效加速土壤有机磷的矿化速率,土壤酶参与土壤肥力形成和演变,其活性易受植物根系分泌物、有机物矿化分解和经营模式的影响[6],上述结果亦说明选用不同经济作物参与复合生草模式对土壤磷素供应水平的影响亦不相同。
就土壤理化性质而言,土壤速效钾和速效磷含量在模式A、B和C间无差异显著,但均高于CK组(P<0.05);模式B的土壤碱解氮含量高于模式A和C(P<0.05);3种处理组土壤含水量均低于对照组CK,但模式A和CK组间差异达显著水平;而有机质及土壤pH值各处理组与CK组间均无显著差异(表3)。

表2 陇东黄土高原地区不同生草模式下杏树林根际土壤酶活性Table 2 Rhizosphere soil enzyme activity of apricot orchard with different grass patterns in eastern Gansu Province
注:平均值±标准误;同列不同字母表示多重比较差异显著(Duncan,α=0.05)。下同。
Notes: Data are means ± SE (n=3); significant differences among treatments within each variable were tested using Duncan’s multiple range comparison (P=0.05) and are indicated by different letters. The following tables use the same representation.
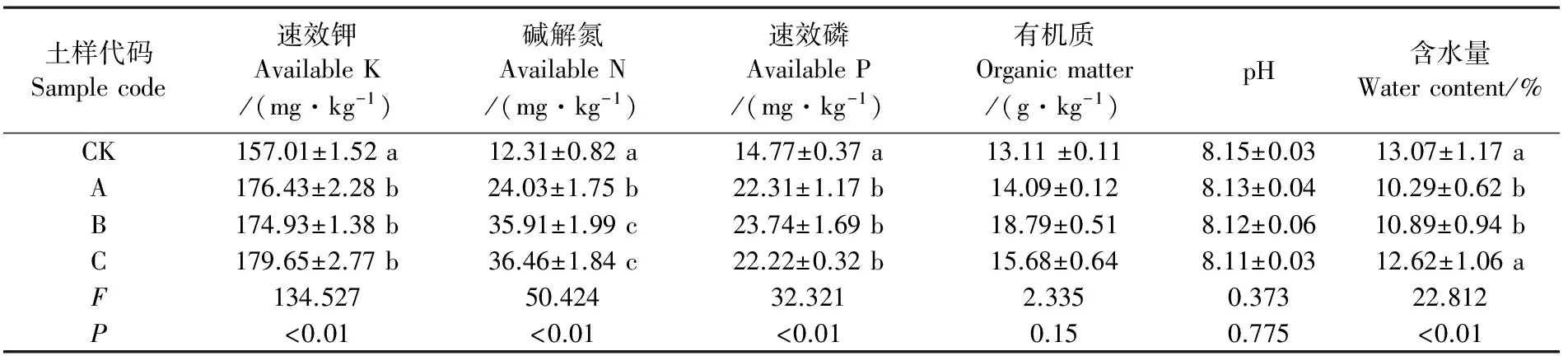
表3 陇东黄土高原地区不同生草模式下杏树林根际土壤理化性质Table 3 Rhizosphere soil physicochemical properties of apricot orchard with different grass patterns in eastern Gansu Province
李会科等[5]对黄土高原渭北苹果园生草土壤性状测定结果表明,生草能降低0~60 cm土层土壤含水量,提高0~40 cm土层碱解氮、速效磷及速效钾含量;Qian等[18]分析了我国黄土高原地区生草覆盖对苹果园土壤养分、酶活性及细菌群落遗传多样性的分析结果显示,种植白三叶草和黑麦草可明显增加土壤速效氮、速效磷、速效钾含量以及土壤含水量,其中豆科植物白三叶草可极显著增加土壤碱解氮含量;惠竹梅等[19]研究了陕西杨凌葡萄生产园行间播种白三叶草、紫花苜蓿和高羊茅,以清耕为对照,分析了土壤微生物数量的变化及其与土壤养分的关系后得出,白三叶草和紫花苜蓿可有效增加土壤碱解氮和速效钾含量,但种植高羊茅可明显降低土壤碱解氮含量。上述试验结果与本研究试验结果基本一致,相较对照组CK,模式A、B和C可有效增加土壤速效钾和速效磷含量(P<0.05),且3组处理间无显著性差异;而由紫花苜蓿参与的模式B和C土壤碱解氮含量明显高于种植黄花菜的模式A(P<0.05);但3种生草模式的土壤含水量却明显低于对照组CK(P<0.05),其中模式C 的土壤含水量与CK间未达显著差异。说明生草品种差异对土壤养分的影响不尽相同,显然豆科植物固氮作用可增加土壤氮素利用率,但种植生草可显著增加0~40 cm土层土壤微生物群落结构[20]及其种群数量[21],加速有机碳的分解,提高土壤养分的矿化速率和营养水平[4]。前人研究得出,种植生草可对果园土壤pH值和有机质含量产生显著影响,但主要集中在0~20 cm的表层土壤[5],而本研究中土壤有机质及pH值在不同生草模式间无显著性差异,究其原因可能是由于陇东黄土高原地区土壤有机质及含水量偏低,而生草果园的土壤有机质库容的演变主要以进化为主,本研究所采集土样深度为0~40 cm,加之6 a的生草年限以及陇东地区土壤较高的盐碱化程度的共同影响,使得陇东黄土高原地区土壤有机质的形成和pH值的改变尚未表现出显著影响,这种情况可能随着生草模式年限的增加而逐步显现。
2.2 杏树林根际土壤微生物群落遗传多样性对不同生草模式的响应
图1为陇东黄土高原地区杏树林不同生草模式下土壤根际微生物群落PCR-DGGE指纹图谱及其模式图。图谱上条带信息反映了土壤微生物群落多样性变化情况,其中电泳条带的多少反映出土壤中细菌群落多样性;条带的粗细则反映了种群密度的差异,条带亮度表征种群分布频率高低。由图1可见,与对照组CK相比,3种生草模式下杏树根际土壤样品DGGE条带数量增多亮度增加,说明种植生草可有效增加陇东黄土高原地区杏树林根际微生物种类和数量,改变其土壤微生物群落结构和组成。
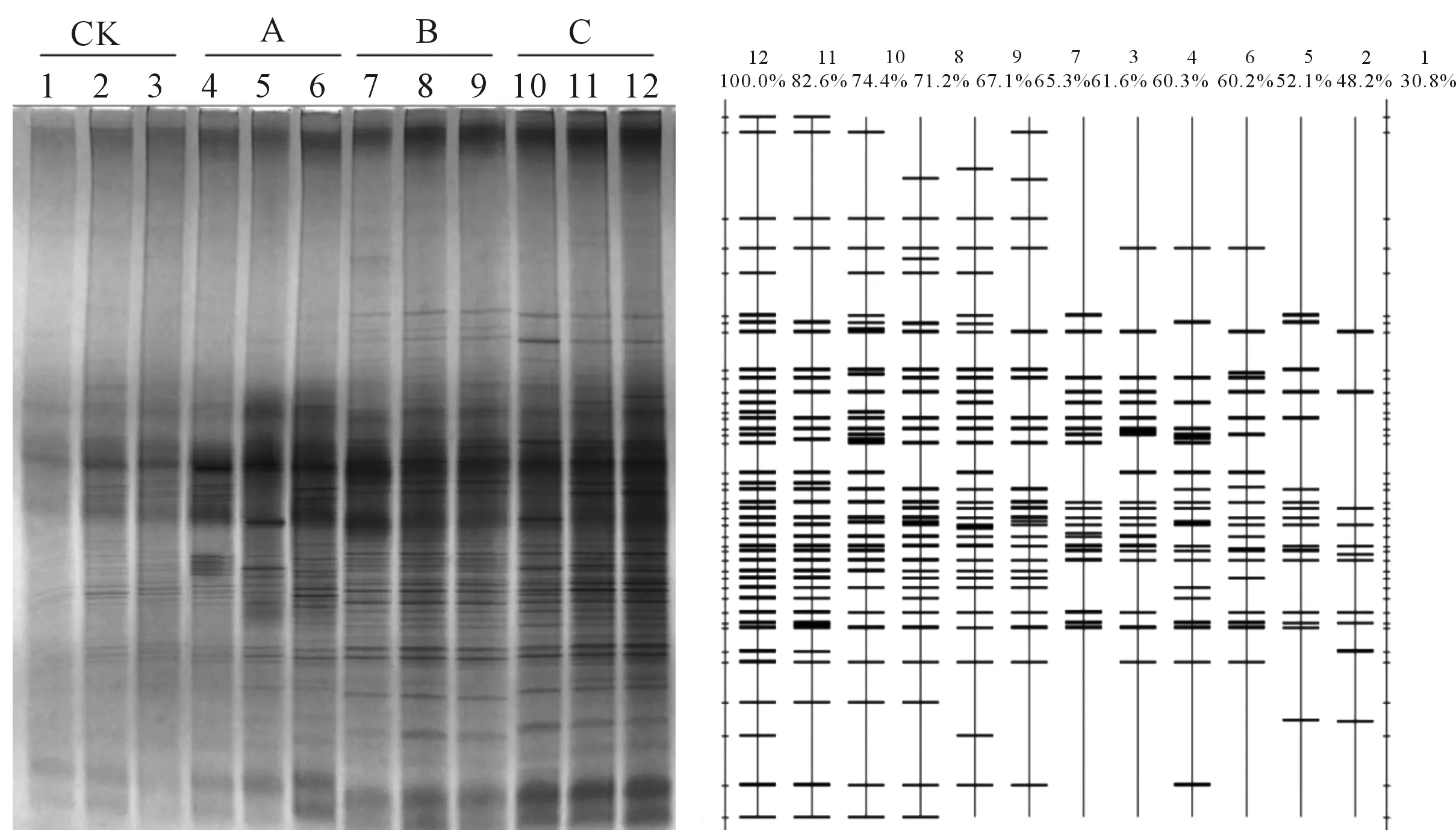
图1 不同生草模式下杏树林土壤根际微生物群落PCR-DGGE指纹图谱及其模式图Fig 1 DGGE band patterns and models of rhizosphere soil microbial community of apricot orchard with different grass patterns
图2反映了陇东黄土高原地区不同生草模式杏树林土壤根际微生物群落多样性指数变化情况。F检验结果显示,4种多样性指数在各样地间差异均达极显著水平,上述结果体现了种植不同生草可对土壤微生物群落结构产生影响。多重比较(Duncan,α=0.05)结果显示,4种多样性指数在不同生草模式下呈相似的变化趋势,即模式C>模式B>模式A>CK (P<0.05),其中Shannon-Wiener指数和Pielou 均匀度在模式B和模式C之间未达显著差异。前人研究得出果园的生草和土地利用方式均可增加土壤微生物的数量[5],改变果园土壤微生物群落结构[21]和组成[20]。本研究结果与上述试验结果相一致,其主要原因是种植生草后有利于增加土壤有机碳的积累和转化速率,加之生草凋落物可为果园土壤微生物提供大量速效碳氮源[17],进而刺激土壤微生物的生长繁殖和有机碳矿化速率;其二,种植生草后地表生物量[22]及群落结构改变可引起地下微生物群落作出响应,进而有助于土壤微生物群落结构和组成的改变[23]。
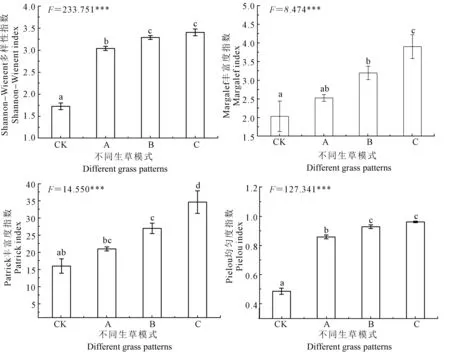
图2 陇东黄土高原地区不同生草模式杏树林土壤根际微生物群落多样性指数变化Fig. 2 Changes on rhizosphere soil microbial community diversity indexes of apricot orchard with different grass patterns in eastern Gansu Province
2.3 陇东黄土高原地区杏树林根际土壤肥力对不同生草模式的响应
对所测定的14项土壤环境变量进行中心化处理后,通过因子分析法对原始环境变量进行信息重组[24],得出各因子的特征根、贡献率及累计贡献率(表4);通过正交旋转得到因子得分矩阵及其载荷矩阵(表5),由表4可见前两个公因子的累积贡献率为86.998%,可用于评价陇东黄土高原地区不同生草模式下杏树林根际土壤肥力变化情况。根据表5分析结果可知,土壤速效钾、碱解氮、速效磷、过氧化氢酶、碱性磷酸酶、转化酶、脲酶、pH值、Shannon-Wiener多样性指数、Margalef丰富度指数及Pielou均匀度指数对第一公因子起正相关作用;土壤有机质、含水量和Patrick丰富度指数则起负相关作用。对第二公因子而言,土壤含水量、碱解氮、有机质、过氧化氢酶、脲酶、Patrick丰富度指数起正相关作用;而土壤速效磷、速效钾、碱性磷酸酶、转化酶、pH值、Shannon-Wiener多样性指数、Margalef丰富度指数和Pielou均匀度指数起负相关作用。
根据14项环境变量在两个公因子中得分矩阵构建因子得分函数:
Y1= 0.162X1+0.056X2+0.206X3-0.148X4+
0.102X5+0.183X6+0.12X7+0.063X8+
0.178X9+0.134X10-0.01X11+0.133X12+
0.128X13-0.104X14
Y2= -0.099X1+0.121X2-0.211X3+0.455X4+
0.036X5-0.169X6-0.009X7+0.107X8-
0.471X9-0.026X10+0.239X11-0.024X12-
0.019X13+0.021X14
式中,Y1,Y2为第一、第二公因子的得分:X1~X14为土壤环境变量的标准化值,分别对应土壤速效钾(X1)、碱解氮(X2)、速效磷(X3)、有机质(X4)、过氧化氢酶(X5)、碱性磷酸酶(X6)、转化酶(X7)、脲酶(X8)、pH (X9)、Shannon-Wiener多样性指数(X10)、Patrick丰富度指数(X11)、Pielou均匀度指数(X12)、Margalef丰富度指数(X13)和含水量(X14)。
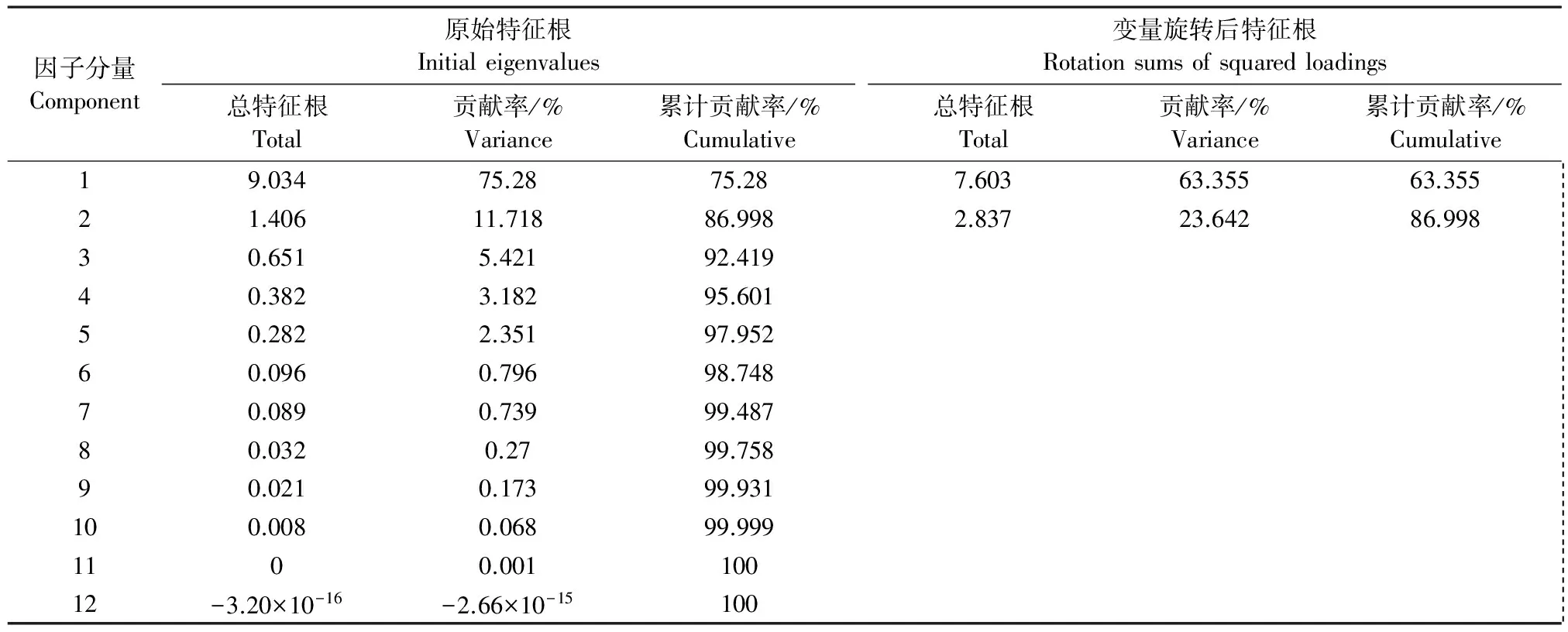
表4 因子分量的特征根、贡献率及累积贡献率Table 4 Factor component characteristic root and contribution rate and cumulative contribution rate
注:提取方法为主成分分析法。
Notes: Extraction method were principal component analysis.
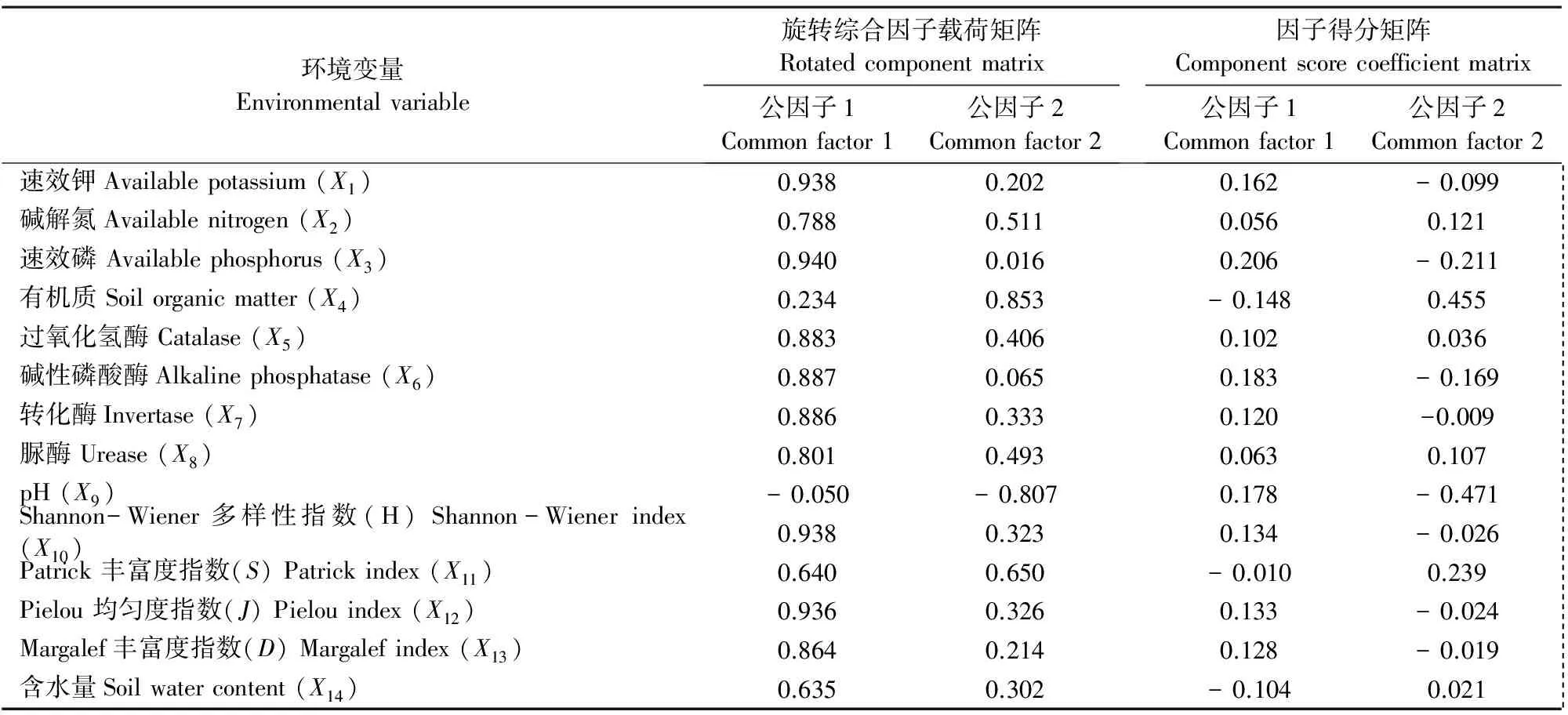
表5 土壤环境因子旋转载荷矩阵及其因子得分矩阵Table 5 Rotating comprehensive factor loading matrix and component score coefficient matrix
注:因子提取采用主成分法,旋转方法为方差最大正交法。
Notes: The factors were extracted by principal component analysis and rotated by varimax, then calculated the factor loading and the score of the factors.
将14项环境变量均值标准化后代入上述两式,根据公因子特征根值计算其权重分别为0.728和0.272,根据W=Y1T1+Y2T2公式加权求和,计算不同生草模式下杏树林根际土壤综合肥力得分(图3),式中W为土壤综合肥力得分;T1,T2为第一、第二公因子的权重。
图3反映了陇东地区不同生草模式下杏树林根际土壤综合肥力变化情况,3种生草模式均不同程度增加了杏树林根际土壤综合肥力;其中以套种紫花苜蓿及黄花菜的模式C土壤综合肥力最高(P<0.05),而模式B和A间差异未达显著水平。前人研究提出果园土壤肥力差异易受生草品种及其播种方式的影响[25],究其原因可能是由于不同生草品种的混合播种对土壤有机碳的积累[26]及土壤微生物多样性[27]的影响不尽相同,进而导致土壤综合肥力的变化存在差异,而这种差异亦随土壤类型及生草年限的差异而不同[28]。本研究结果与上述结果基本一致,以套种紫花苜蓿及黄花菜的模式C土壤综合肥力最高(P<0.05),而模式B和A间差异未达显著水平,但均高于对照组CK,说明陇东黄土高原地区开展果园生草模式需增加生草品种的同时,要选用本地适生豆科植物进行混合播种,将有利于该地区杏树果园土壤综合肥力的增加。
2.4 影响杏树林不同生草模式土壤肥力差异的环境因子
为了阐明不同生草模式下杏树样地与土壤环境因子间关系,以及环境因子在驱动不同样地分异过程中的相对重要性,基于Bray-Curtis相异指数,将所测定的14项土壤环境因子拟合到4种样地的非度量多维尺度(NMDS)排序图上,胁强系数Stress=0.0481,说明该二维NMDS分析具有一定的拟合效果,可解释样地环境因子信息量的99.95%。
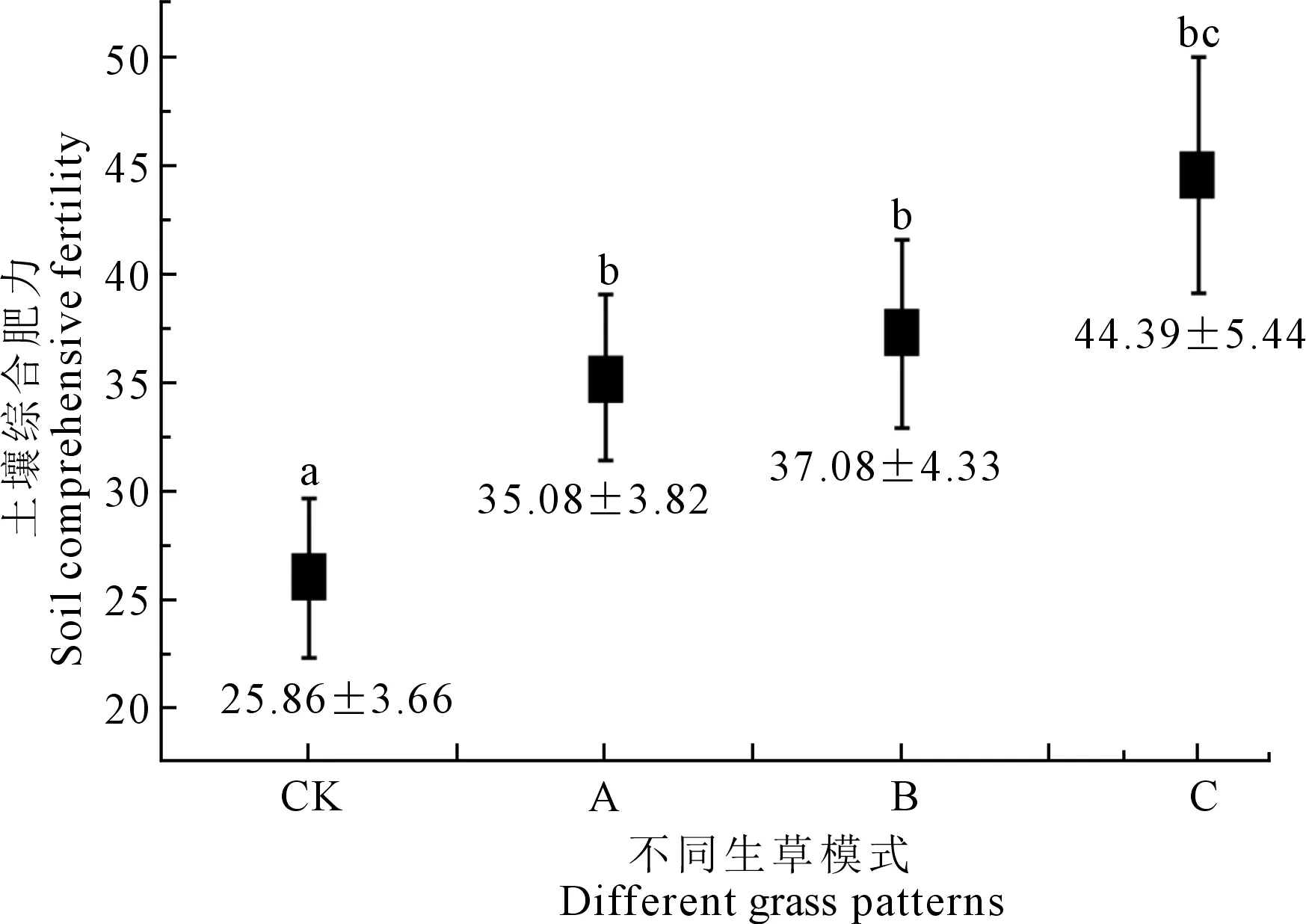
图3 不同生草模式杏树林根际土壤综合肥力变化Fig.3 Changes on rhizosphere soil comprehensive fertility of apricot orchard with different grass patterns
由图4可见,基于Bray-Curtis相异性指数,模式B和C在NMDS排序图上发生重合,而模式A介于模式B、C和CK之间,说明4种样地存在显著差异。14个环境因子变量拟合至NMDS排序图的结果显示,14项环境因子中有9个环境变量与样地NMDS排序结果显著相关。由图4可见,各样地在NMDS排序图上的分异主要与土壤酶活性中的脲酶(r2=0.8111,P=0.002),转化酶(r2=0.743 3,P=0.002)和过氧化氢酶(r2=0.689 1,P=0.007)显著相关;4种土壤微生物群落遗传多样性指数中主要与Patrick丰富度指数(r2=0.848 9,P=0.001),Shannon-Wiener多样性指数(r2=0.697 4,P=0.006)和Pielou 均匀度指数(r2=0.649 7,P=0.009)相关;就土壤理化因子而言,6项理化因子中只有土壤有机质与样地分异有关(r2=0.510 5,P=0.035)。上述结果说明影响杏树林不同生草模式土壤肥力差异的最主要环境因子分别是Patrick丰富度指数和土壤脲酶,其余依次为经营模式(r2=0.758 5,P=0.001)、转化酶、过氧化氢酶、土壤微生物群落Shannon-Wiener多样性指数和Pielou均匀度指数。前人研究[5]得出生草栽培可明显增加杏树根际微生物数量,生草栽培对果园生理生态的影响随草种和生草模式不同而有所差异,如豆科植物有助于土壤氮素积累,禾本科植物能有效提高土壤有机质含量,这与本研究结果相一致,说明陇东地区气候和土壤条件具有独特的地域性,生草栽培对果园土壤生物学及非生物学特性的影响随草种和生草模式不同而有所差别,即如何增加土壤微生物数量以及配植适应陇东地域特点的林下生草对于陇东地区杏树果园的良性发展至关重要。

注:排序图上显示了与各样地分异有关的土壤环境因子,椭圆代表平行样品间的标准差;*代表与各样地分异相关的土壤环境因子显著水平:***表示P<0.001,**表示 P<0.01,*表示 P<0.05Note: The correlations between 13 vectors and 12 rhizosphere soil samples treated by different planting patterns were tested using the R package (based on 999 permutations), and significant vectors (r2 values and P values were shown on the right panels) were fitted to the NMDS ordination plots; Ellipses represent standard deviation among parallel samples图4 陇东黄土高原地区杏树林不同生草模式的非度量多维尺度(NMDS)排序及其与环境因子间关系Fig.4 Nonmetric multidimensional scaling (NMDS) ordination patterns of apricot orchard with different grass patterns and relationship with their environment factors in eastern Gansu Province
2.5 不同生草模式下杏树根际土壤综合肥力的影响因子方差分解
图5为影响不同生草模式下杏树根际土壤综合肥力的环境因子方差分解Venn图,结果显示,本研究所涉及的14项环境变量可划分为3类环境因子(土壤微生物学特性、土壤酶活性及土壤理化特性)。根据方差分解残差可知,3类环境因子总叠加效应共同解释了不同生草模式下杏树根际土壤综合肥力变异的79%(图5);而3类环境因子独立效应分析结果显示,土壤理化特性只解释了综合肥力差异的3%,而土壤酶活性与微生物群落分别解释了15%和13%。从3类环境因子交互作用来看,影响不同生草模式下杏树根际土壤综合肥力的因素主要来自土壤理化特性和土壤酶活性(39%)以及土壤微生物群落和酶活性(41%),而三者的共同作用可解释土壤肥力变异的63% (图5)。

图5 土壤环境因子对不同生草管理模式杏树根际土壤综合肥力的方差分解Fig.5 Variance analysis on rhizosphere soil comprehensivefertility of apricot orchard with different grass patterns
上述结果说明,在陇东地区对杏树林采用不同生草模式后土壤综合肥力出现明显差异,而造成其土壤肥力差异最主要的环境因子为土壤酶活性和土壤微生物学特性。之前在对4种生草模式进行NMDS排序分析时发现,影响不同生草模式土壤肥力差异的最主要环境因子分别是Patrick丰富度指数和经营模式,其余依次为土壤脲酶、转化酶、过氧化氢酶、土壤微生物群落Shannon-Wiener多样性指数和Pielou 均匀度指数,所测定的6项理化因子中只有土壤有机质与样地分异有关,且贡献率为相关因子中最低,这一结果与方差分解结果相吻合。土壤生态系统中理化因子是物质循环、能量转换和信息传递的不可或缺的环节,可与其它环境因子相互作用,共同完成生物地球化学循环过程[6]。但本研究中土壤理化因子对于该地区不同生草模式杏树林土壤综合肥力的单独效应只有3%,但纵观共同效应后可见,土壤理化特性与土壤微生物特性间可共同解释土壤肥力差异的11%,而NMDS排序中土壤有机质含量对于样地分异之间呈显著关系(r2=0.510 5,P=0.035),说明陇东黄土高原地区土壤有机质含量是物质和能量循环的基础,土壤中有机成分的高低直接影响了土壤微生物群落结构和组成,有机质含量高低决定了土壤微生物对营养物质的矿化的速率[29],进而引起土壤微生物数量以及群落结构和组成的改变[4],同时与植物根际分泌物实现共代谢[17],加速无机物的同化作用的同时刺激了植物根际土壤酶活性,进而完成地球生物化学循环[23]。因此,对陇东地区杏树林实施不同生草模式时应增施有机肥,增加生草品种多样性可为其根际微生物提供大量的速效碳、氮源的同时有效刺激杏树根际胞外酶的分泌进而有效提高土壤综合肥力。
3 结 论
本研究以陇东黄土高原地区不同模式(A:杏树+黄花菜;B:杏树+紫花苜蓿;C:杏树+黄花菜+紫花苜蓿;以杏树+清耕作为对照组CK)人工生草6年的杏树林根际土壤为研究对象,测定了14项土壤生物学及非生物学环境因子,利用NMDS排序和方差分解初步分析了不同生草模式下杏树样地与土壤环境因子间关系,以及环境因子在驱动不同生草模式样地土壤综合肥力分异过程中的相对重要性,结论如下:
1)杏树林嵌套种植紫花苜蓿可有效增加土壤氮素循环;黄花菜生草模式可有效加速土壤有机磷的矿化速率;复合生草模式可有效增加土壤微生物群落多样性,进而刺激土壤转化酶活性,加速了土壤有机碳的地球生物化学循环速率的同时,显著降低了土壤过氧化氢的积累(P<0.05)。土壤理化方面,草种差异对土壤养分的影响不尽相同,豆科植物固氮作用可增加土壤氮素利用率。陇东地区土壤有机质及含水量偏低,6a的生草年限和陇东地区土壤较高的盐碱化程度的共同影响,使得该地区土壤有机质的形成和pH值的改变尚未表现出显著影响,而这种情况可能随着生草经营模式年限的增加而逐步显现。
2)基于PCR-DGGE技术分析结果说明生草模式可对土壤微生物群落结构产生显著影响,4种多样性指数在不同生草模式间呈相似的变化趋势,即模式C>模式B>模式A>CK。说明生草有助于增加土壤有机碳的积累,刺激土壤微生物的生长繁殖和有机碳矿化速率,同时草种差异是导致了杏树根际土壤微生物群落结构和组成变化的主要因素,即土壤微生物群落结构和组成随地表植被多样性及群落结构改变而呈不同的响应模式。
3)利用因子分析法探讨了陇东黄土高原地区杏树林根际土壤肥力对不同生草模式的响应,结果显示3种生草模式增加了杏树林根际土壤综合肥力,其中以套种紫花苜蓿及黄花菜的模式C土壤综合肥力最高(44.39±5.44),而模式B和A间差异未达显著水平。说明陇东地区开展果园生草经营模式需增加生草品种的同时,要选用本地适生豆科植物进行混合播种,将有利于该地区杏树果园土壤综合肥力的增加。
4) 基于Bray-Curtis相异指数,将所测定的14项土壤环境因子拟合到4种样地NMDS排序图上,结果显示影响陇东地区杏树林不同生草模式土壤肥力差异的最主要环境因子是Patrick丰富度指数和土壤脲酶,其余依次为经营模式、土壤转化酶、过氧化氢酶、土壤微生物群落Shannon-Wiener多样性指数和Pielou 均匀度指数。陇东地区生草栽培对果园土壤生物学及非生物学特性的影响随草种和生草模式不同而有所差别,即如何增加土壤微生物数量以及配植适应陇东地域特点的林下生草对于陇东地区杏树果园的良性发展至关重要。
5)对不同生草模式下杏树根际土壤综合肥力进行因子方差分解后结果显示,在陇东地区对杏树林采用不同生草模式后土壤综合肥力出现明显差异,而造成其土壤肥力差异最主要的环境因子为土壤酶活性和土壤微生物学特性。土壤中有机成分的高低直接影响了土壤微生物群落结构和组成,因此对陇东地区杏树林实施不同生草模式时应增施有机肥,增加生草品种多样性可为其根际微生物提供大量的速效碳、氮源的同时有效刺激杏树根际胞外酶的分泌进而有效提高土壤综合肥力。
致谢:感谢陇东学院王鑫教授在土样采集及其理化性质和酶活性测定过程中给予的帮助;感谢青海师范大学石国玺副教授在数据分析中的悉心指导。
猜你喜欢
机械研究与应用(2022年3期)2022-07-25
河北果树(2022年1期)2022-02-16
河北果树(2022年1期)2022-02-16
农家致富顾问·上半月(2021年6期)2021-12-26
中国土壤与肥料(2021年5期)2021-12-02
中国土壤与肥料(2021年5期)2021-12-02
中国饲料(2021年17期)2021-11-02
学苑创造·A版(2020年11期)2020-01-07
浙江农业科学(2019年6期)2019-06-24
现代园艺(2018年3期)2018-02-10