
How plant density affects maize spike differentiation, kernel set,and grain yield formation in Northeast China?
2018-08-06 12:08ZHANGMingCHENTaoHojatollahLatifmaneshFENGXiaominCAOTiehuaQIANChunrongDENGAixingSONGZhenweiZHANGWeijian
ZHANG Ming, CHEN Tao, Hojatollah Latifmanesh, FENG Xiao-min, CAO Tie-hua, QIAN Chun-rong,DENG Ai-xing, SONG Zhen-wei, ZHANG Wei-jian
1 Institute of Crop Sciences, Chinese Academy of Agricultural Sciences/Key Laboratory of Crop Physiology and Ecology, Ministry of Agriculture, Beijing 100081, P.R.China
2 Jilin Academy of Agricultural Sciences, Changchun 130124, P.R.China
3 Heilongjiang Academy of Agricultural Sciences, Harbin 150086, P.R.China
Abstract A two-year field experiment was conducted to evaluate the effects of plant density on tassel and ear differentiation, anthesissilking interval (ASI), and grain yield formation of two types of modern maize hybrids (Zhongdan 909 (ZD909) as tolerant hybrid to crowding stress, Jidan 209 (JD209) and Neidan 4 (ND4) as intolerant hybrids to crowding stress) in Northeast China. Plant densities of 4.50×104 (D1), 6.75×104 (D2), 9.00×104 (D3), 11.25×104 (D4), and 13.50×104 (D5) plants ha–1 had no significant effects on initial time of tassel and ear differentiation of maize. Instead, higher plant density delayed the tassel and ear development during floret differentiation and sexual organ formation stage, subsequently resulting in ASI increments at the rate of 1.2–2.9 days on average for ZD909 in 2013–2014, 0.7–4.2 days for JD209 in 2013, and 0.5–3.7 days for ND4 in 2014, respectively, under the treatments of D2, D3, D4, and D5 compared to that under the D1 treatment. Total florets,silking florets, and silking rates of ear showed slightly decrease trends with the plant density increasing, whereas the normal kernels seriously decreased at the rate of 11.0–44.9% on average for ZD909 in 2013–2014, 2.0–32.6% for JD209 in 2013,and 9.7–28.3% for ND4 in 2014 with the plant density increased compared to that under the D1 treatment due to increased florets abortive rates. It was also observed that 100-kernel weight of ZD909 showed less decrease trend compared that of JD209 and ND4 along with the plant densities increase. As a consequence, ZD909 gained its highest grain yield by 13.7 t ha–1 on average at the plant density of 9.00×104 plants ha–1, whereas JD209 and ND4 reached their highest grain yields by 11.7 and 10.2 t ha–1 at the plant density of 6.75×104 plants ha–1, respectively. Our experiment demonstrated that hybrids with lower ASI, higher kernel number potential per ear, and relative constant 100-kernel weight (e.g., ZD909) could achieve higher yield under dense planting in high latitude area (e.g., Northeast China).
Keywords: corn, dense planting, spike differentiation, anthesis-silking interval (ASI), kernel set
Received 14 August, 2017 Accepted 20 December, 2017
ZHANG Ming, E-mail: zhangmingsdau@126.com; Correspondence SONG Zhen-wei, Tel: +86-10-62156856, E-mail: songzhenwei@caas.cn
© 2018 CAAS. Publishing services by Elsevier B.V. All rights reserved.
doi: 10.1016/S2095-3119(17)61877-X
1. Introduction
Maize (Zea maysL.) is one of the most important cereal crops, accounting for more than 40% of the total cereals production in the world (FAO 2013). For the past several decades, grain yield improvement of maize mainly depended upon enhanced tolerance to increased plant density(Tollenaar 1989; Echarteet al. 2000; Duvick 2005; Rossiniet al. 2011). In general, kernel number accounts for most of the variation in yield, which is more important than kernel weight for high grain yield under dense planting (Otegui and Bonhomme 1998; Andradeet al. 1999; Echarteet al.2000). However, kernel number per ear declines sharply with increasing plant density, which may consequently affect the harvested kernel number and grain yield (Otegui 1997; Maddonniet al. 2006). It has been found that kernel number per ear is dependent on ear floret number, which is not only normally affected by environmental conditions but differs among maize genotypes (Otegui 1997; Càrcovaet al. 2000). On the other hand, many studies also have indicated that kernel number per ear can be affected by intra-specific competition of maize due to limited nutrient supply, water availability, and light interception under high plant density (Tollenaar and Lee 2006; Paganoet al. 2007).These limitations result in the decrease in photosynthetic and growth rate per plant, and hence kernel number per ear (Andradeet al. 1999). Changes in plant density could influence final kernel number per ear through effects on the developing ear before flowering, on pollination and fertilization, or on the abortion of kernels after fertilization(Wilson and Allison 1978). For example, Sangoi (2002)and Sarlangueet al. (2007) reported that dense planting reduced the number of total florets and silking florets due to delayed female spike differentiation. Whereas other studies indicated that kernel number per ear was not related to the degree of floral differentiation reached at silking stage but to the kernel abortion of late pollinated silks because of increased the anthesis-silking interval (ASI) under dense planting (Otegui 1997; Uribelarreaet al. 2002; Paganoet al.2007; Ciet al. 2012). Furthermore, it was also reported that shading stress under high plant density led to increased grain abortion, which was associated predominantly with a decrease in kernel number per ear (Tollenaar and Lee 2006; Van Roekel and Coulter 2012; Cuiet al. 2015).These contradictory results arise from uncertainty about the comprehensive effects of plant density on maize spike differentiation, kernel set, grain yield formation, and their relationships.
Northeast China is one of the most important maize production areas in China. The maize cropping area and production in the region account for 29.8 and 32.0% of the total maize area and production of China during the past decade. According to national statistics and our investigation in 2010, the grain yield and plant density of maize were about 5 255 kg ha–1and 52 500 plant ha–1in the region, which account for approximately two-thirds and onesecond of these in U.S. Corn Belt (Duvick 2005; Mansfield and Mumm 2014). Northeast China, a typical high latitude rainfed area, is characterized by limited crop growth duration with low air temperature but adequate solar radiation.Therefore, the effect of dense planting of maize on kernel number in Northeast China may differ from that in middle and low latitude regions with higher air temperature but less solar radiation such as North China Plain (Jacobs and Pearson 1991; Songet al. 2012; Cuiet al. 2015). Although many researches have been done to evaluate the effects of plant population on maize grain yield in Northeast China in recent year (Ciet al. 2012; Liuet al. 2012; Songet al. 2012;Chenet al. 2013), little is known about how plant density of different varieties affects the tassel and ear development,the ASI, and the kernel formation of maize in the region.
Hereby, we conducted a two-year field experiment with two types of modern maize hybrids (tolerant and intolerant hybrids to crowding stress) and five plant density levels to investigate the effect of plant density on spike differentiation,kernel set, grain formation, yield composition of maize, and their relationship in Northeast China. The present study may facilitate the strategy of cultivar selection and adoption of agronomic management for dense planting of maize in Northeast area.
2. Materials and methods
2.1. Experimental site description
The field experiment was conducted at the Gongzhuling Experimental Station, Institute of Crop Sciences,Chinese Academy of Agricultural Sciences (43°30´23´´N,124°48´34´´E, 220 m above sea level) in spring maize growing season during 2013 and 2014. The station is located in the center of Jilin Province, China in the north temperature zone with continental monsoon conditions.Annual precipitation is 562 mm with almost 81% occurring in May to September, and annual average temperature and sunshine duration are 5.6°C and 2 710 h, respectively.The soil is classified as Haplic Phaeozem in FAO-Unesco classification, and Cumulic Hapludoll (Mollisol) in the US classification (Kouet al. 2012). The topsoil (0–20 cm) has the following characteristics: soil bulk density 1.38 g cm–3,soil organic matter 28.1 g kg–1, available nitrogen 143.3 mg kg–1, available phosphorus 66.5 mg kg–1, and available potassium 150.8 mg kg–1(Songet al. 2013). The mean temperature during maize growing season was 20.8 and 20.4°C, respectively, in 2013 and 2014. The precipitation during maize growing season was 553.9 and 434.3 mm,respectively, in 2013 and 2014. More meteorological details are presented in Fig. 1.
2.2. Experimental design and management

Fig. 1 Daily precipitation and mean air temperature during the entire growing period of maize cropping in 2013 and 2014 at Gongzhuling experimental station, Jilin Province, China.
The experiment was a split-plot randomized complete block design with three replicates. Three modern maize hybrids were applied according to their different tolerance to crowing stress (Zhongdan 909 (ZD909) as the tolerant hybrid,Jidan 209 (JD209), and Neidan 4 (ND4) as the intolerant hybrids, respectively). Main plots consisted of maize hybrids ZD909 and JD209 in 2013, and ZD909 and ND4 in 2014,while subplot consisted of five plant density treatments of 4.50×104(D1), 6.75×104(D2), 9.00×104(D3), 11.25×104(D4),and 13.50×104(D5) plants ha–1, respectively. Each plot was 72 m2(10 m×7.2 m) in size, including 12 rows of maize with a row spacing of 0.6 m. The maize was sown on 6 May 2013 and 27 April 2014 and harvested on 27 September 2013,and 25 September 2014, respectively. Adequate chemical fertilizer was applied at rates of 300 kg N ha–1, 75 kg P2O5ha–1, and 120 kg K2O ha–1as urea, triple super phosphate,and muriate of potash, which could eliminate the nutrient limitations under higher plant densities. Before sowing, onethird of N fertilizer and all of P and K fertilizers were applied as a starter fertilizer, while the rest of N fertilizer was taken as a top-dressing at the jointing (V6) stage of maize. The land was manually weeded at 15-day interval during the maize growing season. Pesticides were sprayed to control pests and diseases in all the plots when required.
2.3. Sampling and measurements
Tassel and ear development processIn each plot, three representative plants were taken at 2–3 days interval in 2014 from the V4 (4th-leaf) stage of maize to determine the development process of tassel and ear. In the laboratory,the growth cone was stripped with a dissecting needle,which was fixed using Carnoy’s fluid, then the development process of tassel and ear were photographed with a stereoscopic microscope (SZ61, Olympus Corporation) or a digital camera (DSC-WX300, Sony Corporation). The lengths of tassel and ear, the branches of tassel and the diameter of ear were measured.
Tassel and ear flower rateAll plants were labeled within 10 m2area before anthesis in each plot. The number of anthesised tassel and silked ear were recorded after 16:00 every day. Then the ASI was calculated as follows:
ASI (d)=Date of anthesis stage–Date of silking stage
Where date of anthesised or silking stage were recorded when the percentage of anthesised tassel or silked ear reached 50%.
Ear floret differentiationIn each plot, 10 representative ears were selected to measure ear floret differentiation at the 8th day after silking in each treatment. Ovules with wilting silks indicated that fertilization occurred, while those with turgid silks indicated that fertilization didn’t occur. Ovules without silks indicated degeneration (Cuiet al. 2015). The number of total florets, silking florets, and fertilized florets were recorded. Then the floret fertility rate, kernel set, and abortive rate were calculated as follows:
Floret fertility rate (%)=(The number of fertilized florets/The number of total florets)×100
Kernel set (%)=(The number of normal kernels/The number of fertilized florets)×100
Abortive rate (%)=(The number of total florets–The number of normal kernels)/The number of total florets×100
Grain yield measurementActual grain yields were determined by harvesting three rows of each plot. Fifteen of the harvested ears in each plot were used to determine the yield components. All grain samples were air-dried on the ground by threshing and oven-dried at 70°C to a uniform moisture content of about 14%, then weighed.
2.4. Statistical analysis
The analysis of variances for measuring data was performed independently using the procedure of SPSS 13.0(SPSS Inc, Chicago, IL, USA). The significant differences among the treatments were detected by the least significant differences (LSD) at 0.05 probability level (Steel and Torrie 1980). The relationship between ASI and yield traits was calculated using Pearson correlation coefficients (Chenet al. 2011).
3. Results
3.1. Tassel and ear differentiation
For all hybrids, increased plant density had no significant effect on tassel development. In 2014, the initial time of growth cone elongation (Fig. 2-A-1 and B-1), growth cone segmentation (Fig. 2-A-2 and B-2), spikelet differentiation(Fig. 2-A-3 and B-3), flower differentiation (Fig. 2-A-4 and B-4), and pollen grain formation (Fig. 2-A-5 and B-5) were almost same for hybrids ZD909 and ND4. However, as plant density increased, the length of tassel decreased for all hybrids in all years (Fig. 3). Differences in tassel length occurred in tassel flower differentiation stage (i.e.,approximately 45 days after sowing, DAS). Compared to the D1 treatment of maize hybrids ZD909, JD209, and ND4, tassel length was reduced by 1.1–9.6%, 2.8–16.1%,and 6.1–17.4%, respectively, under D2, D3, D4, and D5 treatments at the beginning of anthesis (i.e., approximately 70–75 DAS). However, the number of tassel branches didn’t show significant differences among the treatments for all maize hybrids (Fig. 4).
The early development of ear showed the same trend as tassel development that no significant differences were found in the initial time of ear differentiation among the treatments and years. In 2014, the growth cone of ear began to elongate at 50 DAS for ZD909 and at 45 DAS for ND4 (Fig. 5-A-1, A-2, B-1, and B-2). However, the floret differentiation was delayed with plant density increased (Fig. 5-A-3 and B-3). With the development of ear growth, floret silk occurred at the base of ear under D1 and D2 treatments at 68 DAS, whereas ear remained floret differentiation under D3, D4, and D5 treatments(Fig. 5-A-4 and B-4). Furthermore, the ovaries in the base of ear under D1 treatment were much higher than that under other treatments at 75 DAS (Fig. 5-A-5 and B-5).As a result, dense planting delayed the ear development since the floret began to differentiate. Moreover, the differences in ear length occurred at approximately 65 DAS among the treatments (Fig. 6). Compared to the D1 treatment, ear length was reduced by 7.3–30.4% on average for ZD909 in 2013–2014, 28.8–53.9% for JD209 in 2013, and 25.2–40.4% for ND4 in 2014, respectively,under D2, D3, D4, and D5 treatments at the beginning of floret differentiate (i.e., approximately 61–65 DAS). Then the differences in ear length between D1 and D2, D3, D4,D5 reached 5.3–30.7%, 11.7–19.5%, and 1.8–38.6% on average for ZD909 in 2013–2014, JD209 in 2013, and ND4 in 2014, respectively, at the end of silking stage (i.e.,approximately 75–90 DAS). Meanwhile, the ear diameter was also reduced significantly along with the plant density increasing (Fig. 7). It was observed that ear diameter was lower under D2, D3, D4, and D5 treatments than that under D1 treatment by 16.4–51.1% and 14.6–47.7% for ZD909 and JD209, respectively at the end of silking stage (i.e.,approximately 90 DAS) in 2014.
3.2. Anthesis-silking interval (ASI)

Fig. 2 Effects of plant density on tassel differentiation for Zhongdan 909 (ZD909) and Neidan 4 (ND4) in 2014. D1, D2, D3, D4,and D5 denote the plant densities of 4.50×104, 6.75×104, 9.00×104, 11.25×104, and 13.50×104 plants ha–1, respectively. A-1–5 denote 38, 47, 56, 64, and 71 days after sowing for ZD909; B-1–5 denote 38, 42, 53, 62, and 71 days after sowing for ND4.
Plant density affected the pollen shedding and floret silking(Fig. 8). As the plant density increased, the anthesis and silking stages (when the rate of anthesised or silking plants accounts for 50% of total plants) of maize was postponed for all hybrids. Furthermore, the effect of plant density on initiation of floret silking among the treatments were higher than that on anthesis stage, then eventually resulting in the higher ASI differences along with increased plant density under all hybrids (Fig. 9). In comparison to D1 treatment for maize hybrids ZD909, JD209, and ND4, ASI increased by 1.2–2.9, 0.7–4.2, and 0.5–3.7 days under D2, D3, D4,and D5 treatments, respectively. Moreover, the silked plant rate decreased under higher plant densities for all hybrids(Fig. 8). Under D3, D4, and D5 treatments, the silked plant rates were 92.6–98.0% on average for ZD909 in 2013–2014,94.1–97.1% for JD209 in 2013, and 75.1–92.7% for ND4 in 2014, respectively.
3.3. Ear florets and grain formation

Fig. 3 Effect of plant density on tassel length in 2013 and 2014. D1, D2, D3, D4, and D5 denote the plant densities of 4.50×104,6.75×104, 9.00×104, 11.25×104, and 13.50×104 plants ha–1, respectively. ZD909, Zhongdan 909; JD209, Jidan 209; ND4, Neidan 4.Values are the mean±stand error.
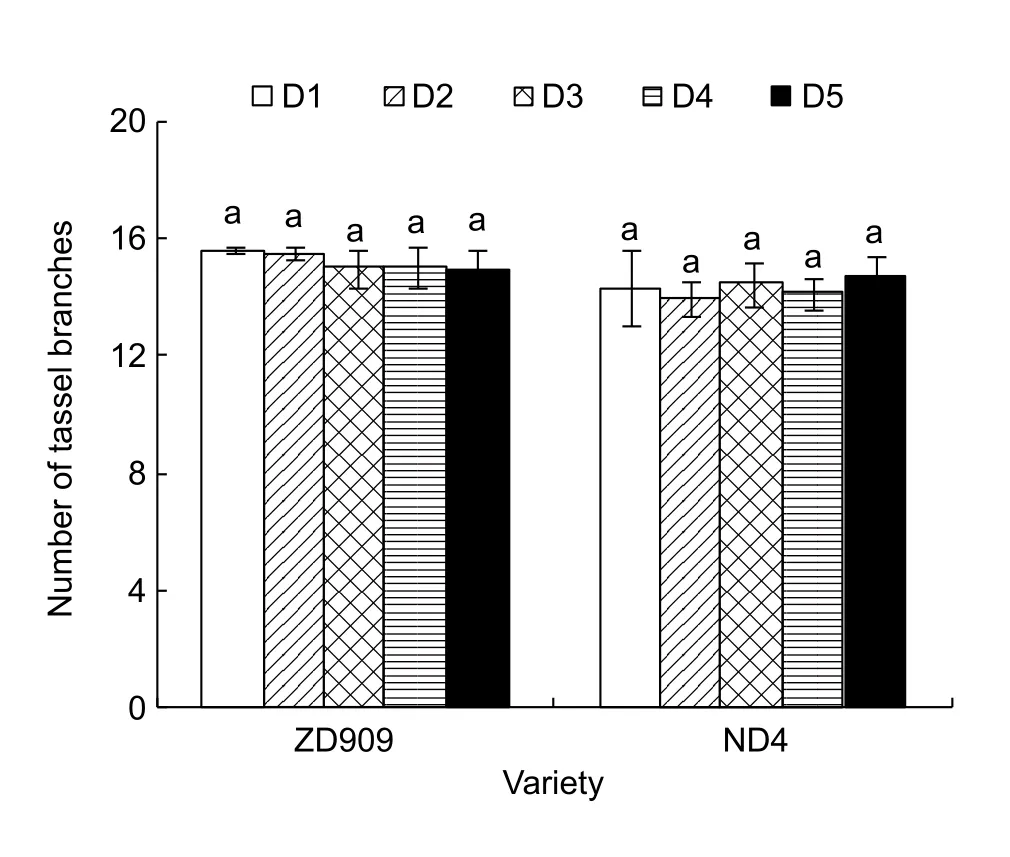
Fig. 4 Effects of plant density on number of tassel branches in 2014. D1, D2, D3, D4, and D5 denote the plant densities of 4.50×104, 6.75×104, 9.00×104, 11.25×104, and 13.50×104 plants ha–1, respectively. Means within each year and variety followed by the same letter do not differ at P<0.05. ZD909,Zhongdan 909; ND4, Neidan 4. Values are the mean±stand error.
Generally, dense planting reduced the number of florets,silking florets, fertilized florets, and normal kernels per ear(Table 1). Compared to the D1 treatment, the decrease rate of silking florets, fertilized florets, and normal kernels per ear ranged in 1.6–15.7%, 2.1–17.5%, 3.6–31.9%,and 2.0–44.9% for all hybrids under D2, D3, D4, and D5 treatments. Moreover, floret fertility rate and kernel set under different plant densities ranged in 73–95% and 74–99% for all hybrids with decrease trends. It was also observed that the abortion rate increased rapidly (ranged from 9–43% on average for ZD909 in 2013–2014, 13–31% for JD209 in 2013, and 17–30% for ND4 in 2014, respectively) as the plant density increased. Moreover, differences among the varieties were observed. ZD909 had the greatest number of floret, silking floret, and fertilized floret per ear which were higher than these of JD209 by 16.1–25.7%, 15.4–24.5%,and 8.9–25.2% under the same plant density, as well as than these of ND4 by 7.0–13.9%, 6.5–17.3%, and 5.8–18.5%,accordingly.
3.4. Grain yield and its components

Fig. 5 Effects of plant density on ear differentiation for Zhongdan 909 (ZD909) and Neidan 4 (ND4) in 2014. D1, D2, D3, D4, and D5 denote the plant densities of 4.50×104, 6.75×104, 9.00×104, 11.25×104, and 13.50×104 plant ha–1, respectively. A-1–5 denote 52, 57, 68, 75, and 79 days after sowing for ZD909; B-1–5 denote 48, 54, 68, 75, and 79 days after sowing for ND4.
The number of rows per ear showed no significant difference among the treatments (Table 2). However,the number of kernels per row reduced by 4.5–22.5%,9.3–28.1%, and 9.0–30.0% under D2, D3, D4, and D5 treatments for ZD909, JD209, and ND4, respectively,compared to that of D1 treatment. Furthermore, hybrid ZD909 showed relative constant 100-kernel weight (ranged from 24.1–28.1 g on average) compared that of JD209(24.0–33.9 g) and ND4 (23.5–32.3 g). ZD909 gained the highest grain yield by 13.7 t ha–1under the plant density of 9.00×104plant ha–1, whereas JD209 and ND4 reached the highest grain yields by 11.7 and 10.2 t ha–1under the plant density of 6.75×104plant ha–1.
3.5. Relationship between ASI and yield traits

Fig. 6 Effect of plant density on ear length in 2013 and 2014. D1, D2, D3, D4, and D5 denote the plant densities of 4.50×104,6.75×104, 9.00×104, 11.25×104, and 13.50×104 plants ha–1, respectively. ZD909, Zhongdan 909; JD209, Jidan 209; ND4, Neidan 4.Values are the mean±stand error.
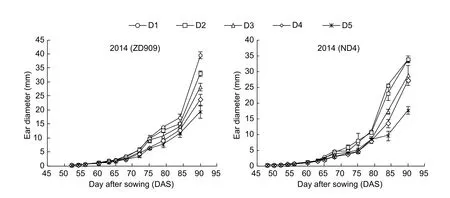
Fig. 7 Effects of plant density on ear diameter in 2014. D1, D2, D3, D4, and D5 denote the plant densities of 4.50×104, 6.75×104,9.00×104, 11.25×104, and 13.50×104 plants ha–1, respectively. ZD909, Zhongdan 909; ND4, Neidan 4. Values are the mean±stand error.
Pearson correlation analysis showed that ASI had a negative effect on the kernel number per row, and 100-kernel weight(Table 3), which suggests that less ASI may mitigate the decrease trends of kernel number per row and 100-kernel weight under dense planting.
4. Discussion

Fig. 8 Effect of plant density on pollen shedding and florets silking dynamics of maize. D1, D2, D3, D4, and D5 denote the plant densities of 4.50×104, 6.75×104, 9.00×104, 11.25×104, and 13.50×104 plants ha–1, respectively. ZD909, Zhongdan 909; JD209,Jidan 209; ND4, Neidan 4.

Fig. 9 Effects of plant density on the anthesis-silking interval (ASI) of maize. D1, D2, D3, D4, and D5 denote the plant densities of 4.50×104, 6.75×104, 9.00×104, 11.25×104, and 13.50×104 plants ha–1, respectively. ZD909, Zhongdan 909; JD209, Jidan 209; ND4,Neidan 4. Means within each year and variety followed by the same letter do not differ at P<0.05. Values are the mean±stand error.
Kernel number per ear of maize is more important than kernel weight for high grain yield under dense planting.Therefore, accurate prediction of kernel number may then be useful in making early estimation of proper plant density and correspondingly grain yield (Otegui and Bonhomme 1998;Andradeet al. 1999; Echarteet al. 2000). In our study, kernel number per ear decreased with the plant density increasingfor all hybrids, suggesting that plant density may affect kernel formation during each stage of the ear development before flowering, the pollination and fertilization, and/or the extent of grain-filling. Although previous studies have found that kernel number per ear are mainly determined by floret number, which may differ due to different maize genotypes (Sarquíset al.1998; Càrcovaet al. 2000), our study have also indicated that kernel number can be affected by specific conditions,such as plant density.

Table 1 Effects of plant density on ear florets differentiation of maize
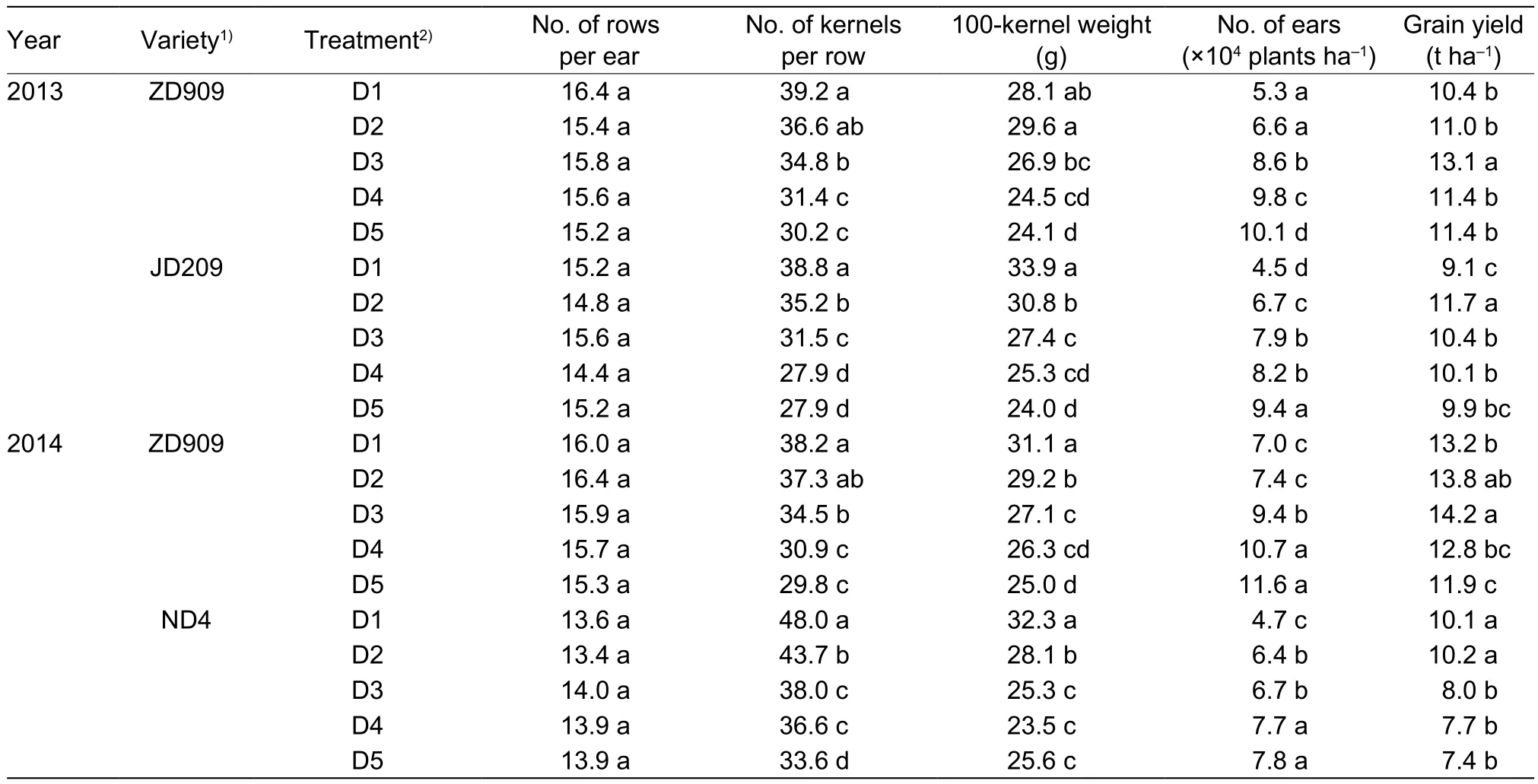
Table 2 Effects of plant density on grain yield and yield components of maize

Table 3 Relationship between anthesis-silking interval (ASI)and grain yield traits
Firstly, higher plant density may postpone the ear development. In our study, although the initial time of ear differentiation was not affected by plant density, laterinitiated ear shoots have a decreased growth rate under higher plant density, resulting in shortened ear length.However, higher plant density had less effect on the transformation of spikelet primordia into functional florets by the time of silking. Floret number and silking floret number showed slightly decreased trends along with the increase of plant density. However, those trends were not significant when plant density increased from 6.75×104to 13.50×104plants ha–1for all hybrids in general. Wilson and Allison (1978) indicated that floret number didn’t affect by plant density. Càrcovaet al. (2000) also suggested that the number of silking floret were mainly determined by different maize genotypes rather than plant density. Our results were similar to previous studies, thus which can be seen that the effects of plant density on ear length were higher than that on total and silking floret number. Consequently, less ear length may reduce sink capacity of maize in turn.
Secondly, higher plant density can affect the floret fertilization. Silking florets of maize earshot may or may not be fertilized which depends on the ASI. In extremely adverse conditions, ASI was so long that results in florets fertilization failure (Otegui 1997). Our study showed that the differentiation period of ears and tassels was delayed substantially, especially the stage of ear floret silking was postponed more than that of tassels anthesis under higher plant density. Under this circumstance, the ASI was extended which resulted in decreased floret fertility rate along with the increase of plant density and reduced final grain yield. On the other hand, pollen shedding is another factor which affects the floret fertility. Although the number of pollen grains was not measured in the present study, the anthesis plant reached 100% under all treatments. Previous research also showed that the amount of pollen produced by tassels were much more than the required amount of ear filament at the largest setting percentage (Càrcovaet al.2000; Edmeadeset al. 2000). Both evidences indicated that the tassel differentiation have less effect on floret fertility. Besides, a smaller tassel size is suggestive of more assimilate partitioning towards the ear (Sangoiet al.2002). Therefore, our results showed that delayed floret differentiation under higher plant density increased the ASI,resulting in decreased number of fertilized floret per ear instead of tassel differentiation.
Thirdly, higher plant density can affect kernel set of maize ear during grain filling stage. Many studies have demonstrated that final kernel number can be affected by plant photosynthesis (Otegui and Bonhomme 1998) and biomass production (Tollenaar 1989) during a critical period.Our study showed that kernel set showed a significant decrease trend under higher plant density (i.e., 11.25×104–13.50×104plants ha–1) during grain filling stage, which led to less final kernel number per ear. High plant density increased intra-specific competition of maize due to limited nutrient supply, water availability, and a light interception during the grain filling stage, hence resulting in fertilized floret abortion (Daynard and Muldoon 1983; Lemcoff and Loomis 1994; Rossiniet al. 2011). Among those limitations,light enter caption might be the key factor. Prine (1971)reported that unfavorable light environment could increase kernel set failure after florets fertilization. Cuiet al. (2015)also indicated that light limitation can decrease N and P absorption which can decrease photosynthetic and growth rate per plant in turn, and hence kernel number per ear in North China Plain. However, in our study, the effect of plant density on kernel set was less than that on floret fertility which may suggest that adequate sunshine hour during the grain filling stage in high latitude area (e.g., Northeast China)may increase the fertilized florets into final grain kernels.
Finally, advanced grain yield of maize under high plant density can be attributed to genetic improvement (Sangoiet al. 2002; Tollenaar and Lee 2006; Ciet al. 2012). Plant density has been recognized as a major factor determining the degree of interplant competition for resources such as incident radiation, soil moisture, and soil nutrients increased in an almost linear fashion with increasing stand density (Fasoula and Tollenaar 2005). While newer hybrids compared to the older hybrids were more tolerant to compound stress (Ciet al. 2011). In present study, the tolerant hybrid ZD909 gained the highest grain yield by 13.7 t ha–1under the plant density of 9.00×104plants ha–1, whereas the intolerant hybrids JD209 and ND4 reached the highest grain yields by 11.7 and 10.2 t ha–1under the plant density of 6.75×104plant ha–1, respectively. The comparison among two types of hybrids showed that the variety of ZD909 has more tolerance for plant density than the intolerant varieties due to shortened ASI, decreased higher rows per ear and kernels per row, and relative constant 100-kernel weight(Paganoet al. 2007; Rossiniet al. 2011; Songet al. 2012).
5. Conclusion
Higher plant density reduced the grain number per ear, which can be the result of a lower flower primordial formation, poor pollination due to flowering asynchrony, and higher grains abortion after fertilization. Furthermore, our study showed that kernel number per ear was not limited by the number of completely developed flowers per ear to an extent, but flowering dynamics was a decisive factor in final kernel number per ear because of longer ASI. Thus, to achieve higher grain yield under dense planting, the varieties with lower ASI and higher ear florets should be applied in high latitude area (e.g., Northeast China).
Acknowledgements
This research was supported by the National Basic Research Program of China (2015CB150404), the National Natural Science Foundation of China (31671642), the Key Program of Science and Technology Department of Jilin Province,China (LFGC14205), and the Innovation Project of Chinese Academy of Agricultural Sciences (CAAS-XTCX2016008).We acknowledge the editor and anonymous referees for their constructive suggestions to improve the manuscript.
 Journal of Integrative Agriculture2018年8期
Journal of Integrative Agriculture2018年8期
- Journal of Integrative Agriculture的其它文章
- Is the “One Province One Rate” premium policy reasonable for Chinese crop insurance? The case in Jilin Province
- ldentification and genetic analysis of multiple P chromosomes of Agropyron cristatum in the background of common wheat
- lnfluence of protein characteristics and the proportion of gluten on end-use quality in Korean wheat cultivars
- Development of EST-PCR markers specific to the long arm of chromosome 6V of Dasypyrum villosum
- ldentification of novel soybean oil content-related genes using QTL-based collinearity analysis from the collective soybean genome
- Silencing of OsXDH reveals the role of purine metabolism in dark tolerance in rice seedlings