
大果榆苗期对不同水分胁迫的生长与生理响应
2018-08-02 01:25刘佳宁李春友贾长荣李艳苹马玉洁
西北林学院学报 2018年4期
刘佳宁,李春友*,贾长荣,李艳苹,王 方,马玉洁
(1.河北农业大学 园林与旅游学院,河北 保定 071001;2.河南省国有济源大沟河林场,河南 济源 454650;3.河北省景县林业局,河北 衡水 053500;4.河北农业大学 林学院,河北 保定 071001)
干旱是自然界中一种常见的气象灾害,现在及未来很长的一段时间内都是人类面临的巨大挑战。干旱不仅制约着人类的生存与发展,也会对植物的分布、生长及产量产生严重影响,因而土壤水分状况与植物之间的关系一直是植物生理生态学研究领域的热点问题之一[1-3]。在遭受干旱胁迫时植物通常有2种应对的方法:一是通过形态学的变化来增强水分吸收并降低水分散失,二是通过一系列的生理生化反应来抵御干旱逆境[4]。因而从植物的生长、生理生化等特征角度出发,开展系统性的研究是了解植物对不同水分胁迫响应与适应的重要切入点[5]。
大果榆(Ulmusmacrocarpa)属于榆科(Ulmaceae)榆属(Ulmus)落叶乔木,别名黄榆、毛榆、柳榆。主产中国东北及华北海拔1 800 m以下地区,树高达20 m,树冠扁球性。喜光,根系发达,侧根萌芽性强,耐寒冷及干旱瘠薄[6]。大果榆冠大荫浓,树体高大,适应性强,在城镇常群植于草坪、山坡或密植作树篱,是著名的行道树,也是北方绿化的主要树种及防风固沙、水土保持和盐碱地造林的重要树种[7]。目前大果榆研究主要集中在苗木培育、药用价值、苗木根系生物量与土壤养分关系和光合指标测定等方面[8-11],而有关大果榆水分生理的研究甚少,本试验以1年生大果榆为材料,通过盆栽试验研究不同土壤水分条件对大果榆生长和生理特性的影响,旨在探究不同程度水分胁迫条件下大果榆的反应和适应能力,了解大果榆的抗旱机理,为大果榆在干旱地区的引种种植提供理论依据。
1 材料与方法
1.1 试验材料
试验地位于河南省济源市大沟河林场内,属暖热带季风气候,年平均气温14.6℃,全年日照1 727.6 h,全年降水量860 mm。试验土壤选用当地林地土壤0~30 cm的表层土(田间持水量为15.05%)。2017年4月选取苗高40~50 cm、地径0.4~0.6 cm、分枝数3~4的生长良好、生长状况基本一致的1年生大果榆盆栽(盆高25 cm、口径30 cm、底径20 cm),每盆装有10 kg表层土,每盆1株。将盆栽苗放置于遮雨棚内,在充足的水分条件下培养。试验共设置4个水分胁迫梯度。对照:适宜水分(土壤含水量为田间持水量的85%~90%);处理1:轻度干旱胁迫(土壤含水量为田间持水量的65%~70%);处理2:中度干旱胁迫(土壤含水量为田间持水量的45%~50%);处理3:重度干旱胁迫(土壤含水量为田间持水量的25%~30%)。每个处理6盆,采用称重法进行控水,每天18:00使用电子秤称取盆重,补充已失去的水分,使各处理维持在所设定的水分范围内。7月19日对水分胁迫的大果榆开始进行持续干旱处理并称重测量,各处理在7月27日达到所控水分范围并开始进行试验。
水分胁迫试验共进行28 d,各个处理在胁迫的第7天(8月2日)、第14天(8月9日)、第21天(8月16日)、第28天(8月23日)早上9:00选取苗木枝条中部的3片成熟叶片,采后迅速放入保温箱内并立即带回实验室进行叶绿素含量、丙二醛(MDA)含量、可溶性糖含量的测定;在第28天(8月23日)进行大果榆水分生理指标及根冠比的测定,每个指标测定均设置3次重复。
1.2 试验指标与测定方法
1)田间持水量:采用环刀法测定。2)叶片含水量:采用饱和含水量法测定。3)水分饱和亏:采用烘干法进行测定。4)叶片保水力:采用自然脱水法测定,用每次称质量叶片含水量占初始含水量的百分比表示。5)叶绿素含量:采用混合液浸提法测定。6)根冠比:采用烘干法测定,用地下部分干重与地上部分干重的比值表示。7)可溶性糖含量:采用蒽酮比色法测定。8)丙二醛含量:采用硫代巴比妥酸法测定。
1.3 数据统计分析
试验数据采用Excel-2003进行计算并运用SPSS21.0进行数据分析。
2 结果与分析
2.1 水分胁迫下大果榆叶片水分生理的变化
植物叶片含水量与其生长代谢活动有着紧密的关系,干旱胁迫下,植物首先表现出来的是体内含水量的下降[12]。由表1可知,处理3的叶片含水量与对照差异显著,是对照的53.02%,处理2和处理1与对照无显著差异。这说明轻度干旱胁迫和中度干旱胁迫对大果榆叶片影响不大,叶片在重度干旱胁迫下受到明显的影响。水分饱和亏是反映植物实际含水量距离饱和含水量的差值[13]。处理3和处理2的水分饱和亏与对照有显著差异,分别是对照的617.46%、484.13%,处理1与对照差异不显著。表明随着水分胁迫程度的加剧,土壤含水量的降低,大果榆通过自身的水分调节,增大水分饱和亏,以适应不利生境,增强抗旱能力,保证生长的顺利进行。
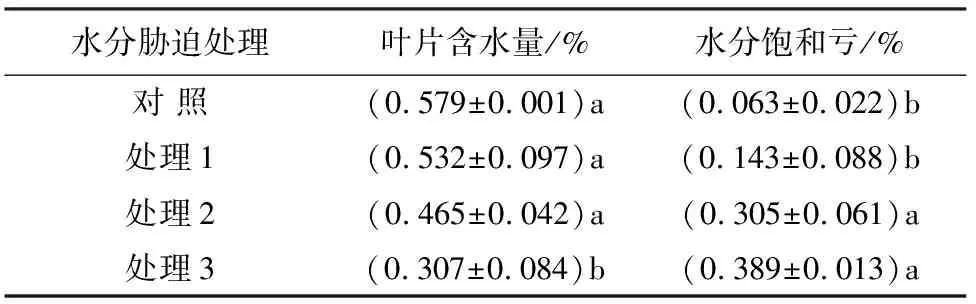
表1 大果榆叶片含水量及水分饱和亏在不同水分胁迫下的变化情况
注:同列小写英文字母不同者表示差异显著(P<0.05)。下同。
叶片保水力是叶片本身的抗旱能力,它反映了叶片内部结构抵御水分散失的能力[14]。由表2可知,随着处理时间的延长,同一处理下叶片保水力逐渐降低,且重度干旱胁迫下叶片保水力的值在每个时间段内均大于其他处理。大果榆叶片在离体时间2 h时处理2的叶片保水力与对照有显著差异,在2 h、7 h时处理3与对照的叶片保水力差异显著,在更长的时间段内4个水分处理下的叶片保水力无显著差异。表明在不同程度水分胁迫下,大果榆在一定的时间内可以较好地维持其细胞正常的生长。
2.2 水分胁迫下大果榆叶片叶绿素含量的变化
叶绿素是植物进行光合作用的主要色素,光合速率的变化与叶绿素含量的高低有着紧密联系[15]。表3显示,叶绿素含量随着水分胁迫程度的增加而逐渐减少。处理7 d后,处理3与对照的叶绿素含量差异极显著,是对照的37.62%,处理2和处理1的叶绿素含量与对照有显著差异,分别是对照的45.49%、59.40%。在水分胁迫14 d后,处理3、处理2和处理1的叶绿素含量均显著低于对照,分别是对照的33.66%、54.93%、58.24%。在处理21 d后,处理3与对照的叶绿素含量有极显著差异,是对照的34.05%,处理2和处理1的叶绿素含量均与对照差异显著,分别是对照的54.46%、63.49%。到水分胁迫后期,处理3和处理2与对照的叶绿素含量差异均极显著,分别是对照的35.21%、38.26%。处理2与对照有显著差异,是对照的73.54%。以上表明水分胁迫可能会抑制叶绿素的生物合成,加速叶绿素分解,使其含量降低。
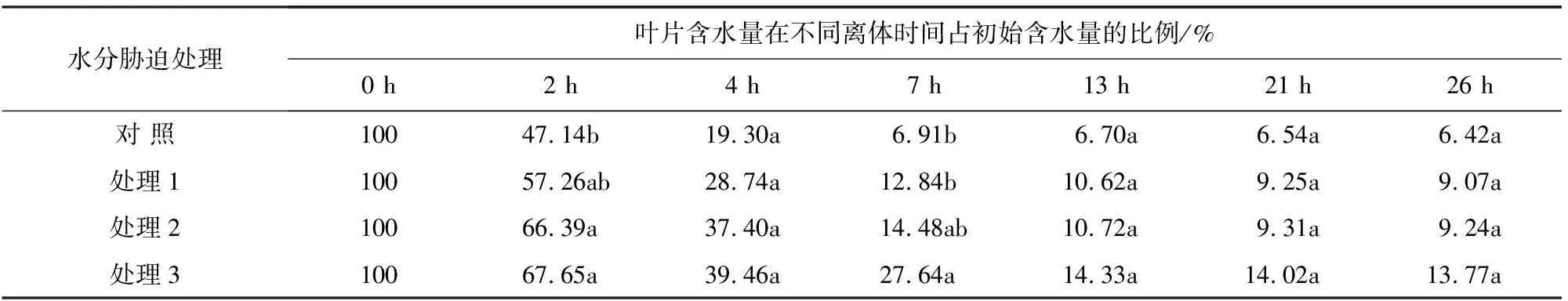
表2 大果榆叶片保水力在不同水分胁迫下的变化情况

表3 大果榆叶绿素含量在不同水分胁迫下的变化情况
2.3 水分胁迫下大果榆根冠比的变化
根冠比是反映地下部分和地上部分生长的重要指标。由表4可见,大果榆苗期根冠比随着水分胁迫程度增强而增加。处理3的根冠比与处理2有显著差异,而与对照差异极显著,是对照的212.29%。处理2与处理1的根冠比无显著差异,与对照差异显著,是对照的144.17%。处理1与对照差异不显著。这说明水分胁迫使大果榆的生长发育受到抑制,但轻度干旱胁迫对大果榆影响较小。

表4 大果榆根冠比在不同水分胁迫下的变化情况
2.4 水分胁迫下大果榆叶片丙二醛含量的变化
丙二醛(MDA)是膜脂过氧化的主要产物之一,它的产生能加剧质膜的损伤。因此,其含量多少标志着叶片脂质过氧化水平,也可间接反映植物组织的抗氧化能力[16]。由表5可知,随着水分胁迫程度的加剧,大果榆叶片的MDA含量增加。处理7 d后,处理3的MDA含量与对照有显著差异,是对照的210.50%,处理2和处理1的MDA含量与对照无显著差异。在水分胁迫的14 d后,处理3的MDA含量与对照差异极显著,是对照的317.53%,处理2和处理1与对照差异显著,分别是对照的170.99%、172.96%。处理21 d后,处理3的MDA含量与对照差异极显著,是对照的409.65%,处理2与对照的MDA含量有显著差异,是对照的241.44%,处理1与对照的MDA含量无显著差异。处理28 d后,处理3的MDA含量与对照差异显著,是对照的255.74%,处理2和处理1与对照的MDA含量差异不显著,表明随着水分胁迫时间的延长,大果榆的叶片在重度干旱胁迫下发生了明显的膜脂过氧化作用。在处理28 d后,轻度、中度和重度干旱胁迫下的大果榆MDA含量较21 d处理时均有不同程度的下降,说明大果榆在水分胁迫下能通过自身的调节机制,逐渐适应干旱环境,降低细胞膜的伤害程度。
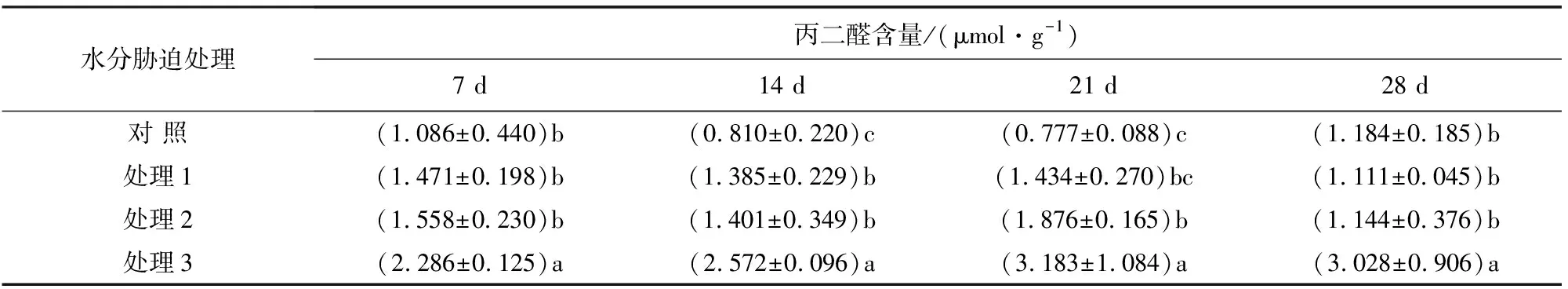
表5 大果榆丙二醛含量在不同水分胁迫下的变化情况
2.5 水分胁迫下大果榆叶片可溶性糖含量的变化
渗透调节是植物在水分胁迫下降低渗透势、抵抗逆境胁迫的一种重要方式,它主要通过植物主动积累溶质来降低细胞渗透势。可溶性糖是植物体内一种重要的渗透调节物质[4]。表6表明,随着水分胁迫程度的加剧,叶片中可溶性糖含量呈现明显上升趋势。处理7 d后,处理3的可溶性糖含量与对照差异显著,是对照的188.02%,处理2和处理1与对照的可溶性糖含量差异不显著。处理14 d后,处理3和处理2的可溶性糖含量显著高于对照,分别是对照的199.16%、154.30%,而处理1的可溶性糖含量与对照无显著差异。处理3和处理2的可溶性糖含量在水分胁迫处理21 d后都达到了最高值,分别是对照的283.30%、190.80%,与对照差异显著,处理1与对照差异不显著。处理28 d后,处理3和处理2的可溶性糖含量均有所降低,分别是对照的208.87%、148.29%,但与对照仍有显著差异。这表明大果榆在干旱胁迫下细胞内溶质数量随着可溶性糖含量的增加而上升,有利于其自身的渗透调节作用,从而提高大果榆的抗旱能力。处理1与对照的可溶性糖含量差异不显著,说明轻度干旱胁迫下大果榆的生理代谢并未受到明显影响。

表6 大果榆可溶性糖含量在不同水分胁迫下的变化情况
3 结论与讨论
植物叶片含水量(RWC)和水分饱和亏(WSD)是衡量植物抗旱能力比较稳定的2个指标[17]。本试验结果表明,RWC随土壤水分的减少而呈降低趋势,WSD则随土壤含水量的下降而升高,这与温翠平[18]对王草(Pennisetumpurpureum×P.glaucum)的研究结果一致,说明水分胁迫下植物组织能够利用的水分减少,李彦瑾[19]等认为相同条件下,植株叶片水分饱和亏的值越高,其抗旱性越强,本研究中,中度和重度干旱胁迫下大果榆叶片较高的水分饱和亏可以避免体内水分的过多散失。离体叶片的持水量是反应干旱条件下叶片抗脱水性能的综合指标之一,离体叶片在萎蔫过程中所保持的水分含量可作为叶片保水力的指标[20]。本试验中,大果榆叶片保水力在各水分处理条件下均有不同程度的降低,其随着土壤含水量的降低而升高并以重度干旱胁迫下叶片保水力最高,这一结果与仁用杏(Prunusarmeniaca×sibirica)叶片的保水力在水分胁迫下的变化方向一致,而与刺槐(Robiniapseudoacacia)和紫穗槐(Amorphafruticosa)的变化方向相反[21],说明水分胁迫下的大果榆叶片为了抵抗水分的散失而保持较高的保水力,以保证细胞内代谢活动的进行。中度与重度干旱胁迫下大果榆叶片的保水力在离体2 h时与适宜水分差异显著,重度干旱胁迫下大果榆叶片在离体7 h时与适宜水分的保水力有显著差异,出现以上现象的原因可能是在叶片离体时间的前期,中度和重度干旱胁迫下的大果榆叶片还具有一定的抗旱能力或者说体内的有关抗旱机制仍在运行,离体叶片还具有较高的保水能力,而随着离体时间的延长,叶片细胞内的生理代谢活动趋于停滞,保持水分的能力逐渐消失。
植物叶片中的叶绿素含量与植物的光合作用和生长发育有着直接关系,其含量是植物遭受逆境胁迫程度的一个重要参考指标[22]。本试验结果表明,轻度、中度、重度干旱胁迫下的叶绿素含量与对照相比均有明显的下降,干旱胁迫程度越严重,叶绿素含量越低。这可能是因为水分胁迫导致叶绿体膜系统受损加剧,膜脂过氧化产生的自由基对叶绿体造成伤害从而降低了叶绿素的含量。这与刘全勇[23]的研究结果相符合。本试验研究的是大果榆叶片的叶绿素总含量在不同干旱胁迫条件下的变化情况,至于水分胁迫下叶绿素a、叶绿素b的变化情况以及对胁迫的敏感度还需进一步的分析。
有研究表明,植物在受到资源胁迫时,通常都会改变地上地下生物量的分配比例以适应环境变化[24]。本研究结果显示,随着水分胁迫程度的加剧,大果榆的根冠比显著升高,并且在重度干旱胁迫下根冠比达到最高值,这与安玉艳[25]对杠柳(Periplocasepium)幼苗的研究结果相吻合。显然,随着土壤含水量的减少,大果榆为了满足对水分及营养物质的需求,改变了地上部分与地下部分生物量的分配,从而增强了植株在干旱环境下的适应能力。
植物体内MDA的含量可以从一方面体现出植物遭受逆境胁迫的程度,细胞膜脂过氧化作用的强弱和质膜损伤程度与MDA含量的高低有密切的关系[26-27]。本试验中,土壤含水量越低,大果榆叶片中的MDA含量越高,该结果与前人在红松[28](Pinuskoraiensis)、葡萄[29](Vitisvinifera)上所得的研究结果一致。轻度干旱胁迫的MDA含量在胁迫14 d后与适宜水分有明显差异,中度干旱胁迫在胁迫14 d、21 d后与适宜水分的MDA含量差异显著,而在水分胁迫的7 d、28 d后三者的MDA含量均无显著性差异。说明可能是由于轻度和中度干旱胁迫的胁迫程度没有超出大果榆自身的特定阈值,膜脂过氧化反应并不强烈,其具有的各种保护机制使得MDA含量维持在一定水平,也说明轻度和中度干旱胁迫下的大果榆具有较强的抗旱能力,在胁迫初期,轻度和中度干旱胁迫下大果榆叶片产生的MDA较少,对细胞膜的影响较小,在胁迫中后期大果榆通过自身的机理调节,减弱膜脂过氧化作用,从而降低了体内的MDA含量。重度干旱胁迫下,大果榆叶片的MDA含量与其他3个处理均有明显差异,说明大果榆在重度干旱胁迫下膜脂过氧化作用最强、细胞膜的受损程度最为严重,这与水分胁迫对文冠果[30](Xanthocerassorbifolia)的影响研究结果相同。
在干旱逆境条件下,植物体内可溶性糖含量会出现不同程度的增加[31]。本试验结果显示,轻度干旱胁迫下大果榆叶片中的可溶性糖含量与适宜水分无明显差异,说明大果榆叶片在轻度干旱胁迫下的细胞渗透压与适宜水分下的细胞渗透压水平相当。但中度和重度干旱胁迫下大果榆叶片中可溶性糖含量增加明显,这与吉增宝[32]对侧柏(Platycladusorientalis)、王川[33]对小麦(Triticumaestivum)的研究结果一致。说明大果榆通过积累可溶性糖来提高自身吸水能力,增强细胞的渗透调节能力,是大果榆在水分胁迫下的一种自我保护机制。其中中度和重度干旱胁迫下可溶性糖含量在21 d后均有不同程度的下降,可能是由于土壤含水量的剧烈下降使植物的生理代谢活动受到抑制,也可能是大果榆在水分胁迫的后期对生境表现出一定的适应能力,针对这一结果还需要进一步的验证。
综上所述,水分胁迫下大果榆幼苗能通过调节自身水分状况,改变地上部分与地下部分的生物量分配加大根冠比,降低细胞膜的伤害程度,同时通过积累渗透调节物质来提高植株的吸水和保水能力,从而有效抵御干旱逆境,维持植株正常生长。植物的抗旱性是一个复杂的问题,是多种因素共同作用下的体现,本文只研究了夏季不同程度水分胁迫下大果榆苗期一些生理指标的变化,还不能全面解释苗期大果榆的抗旱能力。因此,在今后的试验中可以考虑水分胁迫对整个生长季内大果榆叶片光合作用机理和相关酶活性的影响,从而为全面了解大果榆的抗旱机制及干旱地区大果榆的推广利用提供更全面的理论依据。
猜你喜欢
阅读(科学探秘)(2020年8期)2020-11-06
种子(2019年8期)2019-09-11
绿色科技(2019年2期)2019-05-21
天然产物研究与开发(2019年1期)2019-03-01
中国果业信息(2019年1期)2019-01-05
中成药(2018年8期)2018-08-29
中国饲料(2016年12期)2016-11-07
中国眼镜科技杂志(2016年18期)2016-11-03
医学研究杂志(2015年9期)2015-07-01
中国当代医药(2015年9期)2015-03-01
