
不同水分胁迫情境下冬小麦生长发育的RZWQM2模拟
2018-07-28 03:19蒋腾聪窦子荷何建强
农业机械学报 2018年7期
蒋腾聪 窦子荷 姚 宁 冯 浩 于 强 何建强
(1.西北农林科技大学旱区农业水土工程教育部重点实验室,陕西杨凌 712100;2.西北农林科技大学中国旱区节水农业研究院,陕西杨凌 712100;3.中国科学院水利部水土保持研究所黄土高原土壤侵蚀与旱地农业国家重点实验室,陕西杨凌 712100)
0 引言
半干旱地区是我国重要的农业类型区之一,水分是限制该地区农业生产的主要因子[1]。近年来随着全球气候变暖,作物生长期关键生育期干旱发生频率及强度的增加,给该地区农作物生长发育造成较大影响[2]。关中地区位于中国西北部,冬小麦是其主要粮食作物之一。该地区秋季雨多、春季干旱,水分亏缺已经成为制约冬小麦稳产、高产的主要因素。因此,探讨受旱条件下冬小麦生境过程,提高水分利用率,发展节水农业,对提高该地区的粮食产量至关重要[1]。
以往有关水分胁迫的研究以大田试验为主[3-6],但是试验周期长、成本高,试验结果具有明显的区域性,无法开展大量的重复试验。而作物模型是以作物为研究对象,根据农业系统学与作物科学原理,对作物与环境、经济因子及其关系的定量化表达,经校正和验证并达到一定精度后,可用于作物生长发育过程及结果的定量预测、监测、预警与决策支持,以促进作物生产实现高产、优质、高效、生态与安全的目标[7]。利用作物模型开展“数值模拟试验”对大田试验进行补充,可在某些方面发挥重要作用且已经得到越来越广泛的应用[8-10]。目前有关小麦生长的模拟模型主要有Sirius[11]、AFRCWHEAT2[12]、APSIM[13]、DSSAT[14]和RZWQM[15]等。
目前,大多数作物模型都是通过“水分胁迫因子”即水分供需比来描述水分胁迫对作物生长和发育过程的影响,但它们的表达形式有所不同[16-17]。例如,APSIM模型以相对有效含水率来描述水分胁迫与氮素固定和物候发育的关系,而以水分供给与需求的比值来描述水分胁迫对光合作用和叶片伸展的影响[17];DSSAT模型则通过比较潜在蒸腾(或植物水分需求)和潜在根系吸水(或植物可吸收的土壤水)之间的大小关系来描述水分胁迫对作物的影响[17]。然而,现有作物模型对水分胁迫条件下作物生长的模拟仍存在一定的误差,例如,姚宁等[18]利用DSSAT中的CERES-Wheat模型模拟旱区冬小麦生长过程时发现,在越冬期和返青期受旱条件下,模拟结果相对较差,且随着受旱时段提前和受旱程度的加重,模拟精度更低。
根区水质模型(RZWQM)是由美国农业部农业系统研究所开发,于1992年推出的农业系统和资源管理模型。该模型综合了物理、生物和化学过程,模拟农业生产系统中作物生长及水分、养分和杀虫剂在作物根区的运动和传输[19]。虽然该模型在各地得到了广泛的应用和验证,但也存在对水分胁迫条件下作物生长模拟精度较低的问题。例如,MA等[20]利用RZWQM模型模拟大豆生长时发现,严重水分胁迫条件下模拟产量与实测值差距较大;YU等[21]利用RZWQM模型模拟华北平原冬小麦-夏玉米轮作体系时发现,冬小麦生长过程中出现水分胁迫时叶面积指数被低估;SASEENDRAN等[22]应用RZWQM模型时发现,较高的水分胁迫会导致大豆叶面积指数和产量被低估。此外,房全孝等[23]利用RZWQM模型模拟小麦产量时发现,严重干旱处理的小麦叶面积指数模拟值明显低于实测值,该模型模拟作物对严重土壤水分胁迫的响应机制还需进一步改善。
为增强RZWQM2模型模拟水分胁迫条件下作物生长的功能,SASEENDRAN等[24]于2014年在该模型原有水分胁迫因子WSI1基础上,提出了两种新的作物水分胁迫因子WSI2和WSI3。其中,WSI2以WSI1为基础,改变了根系吸水项的计算公式;WSI3则以WSI2为基础增加了土壤蒸发项。然而,自新的水分胁迫指数建立以来,还未在我国干旱和半干旱地区进行验证,RZWQM2模型模拟干旱胁迫条件下冬小麦生长的精度是否相应得到了提高,目前尚不清楚。
因此,本研究比较RZWQM2模型提供的3种水分胁迫因子,判断新增的水分胁迫因子是否提高RZWQM2模型模拟存在水分胁迫时冬小麦生长过程的精度,验证RZWQM2模型在我国干旱和半干旱地区的适应性,并选出适用的水分胁迫因子,从而为进一步将该模型应用于我国干旱半干旱地区农业生境系统管理研究提供参考。
1 材料和方法
1.1 田间试验设计

图1 2012—2013年和2013—2014年两年度冬小麦生育期内逐日气温和太阳辐射量Fig.1 Daily temperature and solar radiation in 2012—2013 and 2013—2014 growing seasons of winter wheat
冬小麦田间分段受旱试验在陕西杨凌西北农林科技大学节水灌溉试验站(108°04′E, 34°17′N,海拔506 m)大型活动遮雨棚下进行,降雨时关闭遮雨棚,以隔绝降雨对试验的影响。试验小区种植面积为8 m2,小区间设有埋深为1.5 m的聚乙烯塑料隔离层,可防止侧渗。试验区土壤为塿土,0~20 cm土层土壤pH值为8.14,有机碳质量比8.20 g/kg,全氮质量比0.62 g/kg。该地区属于暖温带季风半湿润气候区,降水量年内分布不均。2012—2013年和2013—2014年两年度冬小麦生育期内气温和太阳辐射量动态变化见图1。
试验因素为灌溉水平和受旱时段。灌溉水平设置灌水定额40 mm(I1)和80 mm(I2)2个水平;受旱时段分为越冬+返青时段受旱(D1)、返青+拔节时段受旱(D2)、拔节+抽穗时段受旱(D3)和抽穗+灌浆时段受旱(D4)4个水平;对照处理(CK)灌水定额为80 mm,各生育期全灌水。共计9个处理,各处理3个重复(表1)。供试冬小麦品种为“小偃22”,条播种植,播深5 cm,行距25 cm,密度400万株/hm2。播种时间为2012年和2013年的10月15日,分别于次年6月2日和6月7日收获。播前施用N 140 kg/hm2和P2O550 kg/hm2,生长期内不再追肥。其他试验细节参照姚宁等[25]的研究。

表1 冬小麦不同生长阶段受旱试验的灌水处理Tab.1 Experimental treatments of water stress at different stages of winter wheat mm
1.2 RZWQM2模型与水分胁迫因子
1.2.1RZWQM2模型简介
RZWQM2模型是一个基于过程的农业系统模拟模型,主要包括6个子模块:物理运移模块、化学反应模块、养分循环模块、杀虫剂反应模块、作物生长模块和管理操作模块。其中物理运移模块用Green-Ampt方程[26]描述土壤水分入渗过程;Brooks-Corey (BC)方程[27]描述土壤水分特征曲线;Richards方程[28]描述分层土壤间土壤水分重分布情况。RZWQM2模型嵌套了DASST 4.0模型[19],用来模拟作物生长发育过程和最终产量。本文中以DSSAT 4.0中的CERES-Wheat模块模拟冬小麦的生长过程和最终产量。
1.2.2RZWQM2模型中的3种不同水分胁迫因子
(1)水分胁迫因子WSI1
RZWQM2模型中延用了DSSAT 4.0中的水分胁迫因子,即潜在根系吸水量与潜在蒸腾量的比值[24]。本文将其作为第1种水分胁迫因子WSI1。
在水分充足的条件下,潜在根系吸水量大于潜在蒸腾量,不存在水分胁迫。随着土壤水分被根系吸收,土壤变干,潜在根系吸水速率下降。在某一阶段达到一个阈值,即膨压因子(ITURFAC),该因子主要影响叶片伸展性生长。对于C3、C4植物,低于膨压因子根系吸收的水分将不能通过渗透调节和细胞壁的扩展来维持膨压,供给细胞分裂和叶片伸展性生长。当潜在蒸腾量等于或超过潜在根系吸水量时,胁迫因子ISWFAC被激活,主要影响光合作用及其他与干物质积累相关的过程。模型中这两个因子被作为直接乘数应用于叶片生长和干物质积累的模拟中,参数范围为1(没有胁迫)到0(完全胁迫)。相关计算式为
(1)
(2)
式中ISWFAC——影响光合作用及其他与干物质积累相关过程的水分胁迫因子
PTRWUP——潜在根系吸水量,mm
PRWUEP1——物种特性参数
EP0——潜在蒸腾量,mm
其中,PRWUEP1目前在DSSAT中设置为1.5,表明在潜在根系吸水量为潜在蒸腾量的1.5倍时,作物的伸展性生长开始遭受水分胁迫;潜在蒸发蒸腾量(EP0)用改进的Shuttleworth-Wallace ET模型[29]计算得到。
潜在根系吸水量(PTRWUP)由Ritchie方程[30]得到,为
抗震设防烈度为8度,按罕遇地震设计基本地震加速度值为0.20g,设计地震属于第一组,场地类别为Ⅱ类,则Tg=0.4s。阻尼比取0.02,地震影响系数曲线的阻尼调整系数按1.0采用,竖直地震载荷对整个结构的动力响应影响较小,一般情况下,只考虑水平向地震作用[5],故分别在结构的两个水平主轴方向计算地震作用。由图3求得加速度谱值如表2。
(3)
式中RRLV(i)——第i层的根长密度,cm/cm3
k1——常数,取0.001 32
k2——常数,当土壤水排水下限大于等于0.30 cm3/cm3时,k2取45.0;当土壤水排水下限小于0.30 cm3/cm3时,k2=130θLL(i)
k3——常数,取7.01
θSW(i)——第i层土壤体积含水率,cm3/cm3
θLL(i)——第i层作物可利用水分下限,cm3/cm3
ΔZ(i)——土层i的深度,cm
(2)水分胁迫因子WSI2
由于式(3)基于单根吸水的径向流理论,该理论存在诸多假设[31]。首先,该理论假设土壤导水率不变,即凋萎系数时的土壤导水率。这一假设在土壤含水率较高时存在很大误差。其次,该理论假设即使在土壤干燥的条件下作物根系和土壤间的水势梯度也为常数。实际中,根水势随时间变化,水势梯度也在变化[24]。Nimah and Hanks方程[32]是一种计算根系吸水的宏观模型,较式(3)没有上述假设。因此SASEENDRAN等[24]希望通过改用Nimah and Hanks方程计算水分胁迫因子中的根系吸水项,以提高水分胁迫的模拟精度,得到第2种水分胁迫因子WSI2。Nimah and Hanks方程为
(4)
式中Hr——根部有效压力水头,cm
Rr——根阻力项
h(z,t)——土壤基质水头,cm
s(z,t)——考虑含盐量的渗透水头,假设为0 cm
Δx——根表面到土壤中测量h(z,t)点的距离,假设为1 cm
Δz——深度增量,cm
R(z)——有效根密度函数,即为在Δz的深度间隔内其有效根与总有效根的比率
K(θ)——土壤导水率,cm/h
Sr(z,t)——根系吸水量,cm
每层瞬时吸水量的累计值即为根系吸水量。通过调节根部有效压力水头(Hr)使总的根系吸水量小于或等于潜在蒸腾量。Hr=15 295.8 cm时的根系吸水量为潜在根系吸水量。
WSI2相关计算式为
(5)
(6)
式中PTRWUP-NH——通过Nimah and Hanks方程计算的潜在根系吸水量,mm
(3)水分胁迫因子WSI3
当土壤干燥时,用于蒸散的能量较少,因而用于加热土壤、空气和冠层的能量较多,SASEENDRAN等[24]将这部分能量导致的作物水分胁迫考虑进水分胁迫因子中。通过修改WSI2中SWFAC的计算公式(式(5)),即在其分子中加上土壤实际蒸发量,分母中将潜在蒸腾量改为潜在蒸发蒸腾量,提出第3种水分胁迫因子WSI3,即
(7)
(8)
式中ES——土壤蒸发的可用水量,通过Richards方程求解土壤表层垂直向上的水流通量得到,上限为潜在土壤蒸发量,mm
ET——潜在蒸发蒸腾量,mm
1.3 模型输入数据
模型运行所需的2012—2014年逐日气象数据(包括最高气温、最低气温、大气相对湿度、太阳辐射和风速等)由试验地附近的陕西省杨凌国家一般气象站提供。试验在遮雨棚下进行,故模型中降雨量为0 mm。模型中土壤质地、颗粒组成及容重由田间试验测得,凋萎系数、田间持水率和饱和度为实测数据,土壤的饱和导水率采用模型默认值(表2)。模型中最大根深设为200 cm,根系分布指数为3.0[33-34]。因缺乏实测值,且本研究没有考虑氮素对作物生长发育的影响,所以土壤养分数据采用模型默认值。

表2 试验区初始土壤性质Tab.2 Initial soil properties of experimental plots
研究表明,选取不存在水分亏缺试验处理的数据来校正模型参数较为合理[18,35]。因此本研究用完全灌水处理(CK)的土壤水分动态、物候期、生物量和产量数据对模型进行校正,其他处理用于模型验证。在模型校正和验证过程中选用模型默认的水分胁迫因子WSI1。在对RZWQM2模型中CERES-Wheat模块进行作物品种参数校正时,以姚宁等[18]得到的“小偃22”的品种参数作为初始值,利用RZWQM2模型自带的PEST软件对冬小麦品种参数进行调试(表3)。模型中作物品种参数反映的是作物品种的遗传特征[36],因此在利用水分胁迫因子WSI2、WSI3对冬小麦生长过程进行模拟时也采用了上述的作物品种参数。

表3 冬小麦品种“小偃22”的遗传参数Tab.3 Genetic parameters of winter wheat cultivar of “Xiaoyan 22”
1.4 3种水分胁迫值的模拟效果评价
(9)
(10)
其中
(11)
式中RMSE——均方根误差
Si——第i个模拟值
Oi——第i个观测值

n——实测值的个数
2 结果与分析
2.1 RZWQM2模型的校正和验证
模型校正和验证后,两年度CK处理下20~40 m土层内土壤动态含水率的RRMSE分别为13.72%和14.05%。各受旱处理土壤含水率模拟值相比,低水水平各处理的模拟精度低于高水水平处理,同一灌水水平下前期受旱各处理的模拟精度低于后期受旱处理。但模拟的各受旱处理下土壤含水率的变化趋势与观测值基本一致,RRMSE为7.21%~13.49%,模拟效果整体较好。
从模型校正的结果可以看出,2012—2013年和2013—2014年两年度CK处理的开花期和成熟期模拟值与观测值基本一致(表4),表明RZWQM2模型可以很好地模拟充分供水条件下的冬小麦物候期。而各受旱处理开花期和成熟期的模拟值之间没有差异且分别与同年的对照处理相同,其中D1处理下模拟值和实测值物候期最大相差9 d,表明RZWQM2模型没有考虑水分胁迫对冬小麦物候期的影响。
两年度CK处理最终生物量和籽粒产量ARE的平均值为8.29%和3.62%,模拟精度较高。但各胁迫处理下冬小麦生物量和籽粒产量的模拟结果相对较差,且模拟误差随受旱时期的提前而增大(表4),其中D1处理严重低估了冬小麦的产量和生物量。2012—2013年度高水处理I2D1的生物量和产量均低于低水处理I1D1,与实测结果不符。这可能是由于I2D1处理的初始含水率低于I1D1处理,模型模拟过程中后期高灌水量没能抵消前期水分胁迫对冬小麦生长造成的影响,导致最终产量的下降。

表4 RZWQM2模型的校正与验证结果Tab.4 Results of calibration and verification of RZWQM2 model
2.2 不同水分胁迫因子的模拟结果比较
2.2.13种水分胁迫因子输出值的模拟结果比较
在用RZWQM2模型模拟冬小麦生长发育的过程时,水分胁迫因子的输出值反映作物对水分胁迫的响应。本研究中冬小麦越冬期和拔节期受旱处理下(D1),随着冬小麦的生长,WSI1最先表现出水分胁迫,然后是WSI3、WSI2,胁迫强度由大到小依次为WSI1、WSI3、WSI2,但越冬期末(播种后120 d)胁迫强度由大到小依次为WSI3、WSI2、WSI1(图2a、2b)。其余各处理3种水分胁迫因子反映出的受旱时段基本相同且与试验设置基本一致,受旱程度由大到小均为WSI3、WSI2、WSI1,胁迫程度的差异随受旱时段的推后而减小(图2,水分胁迫因子范围0(没有水分胁迫)到1(完全胁迫))。
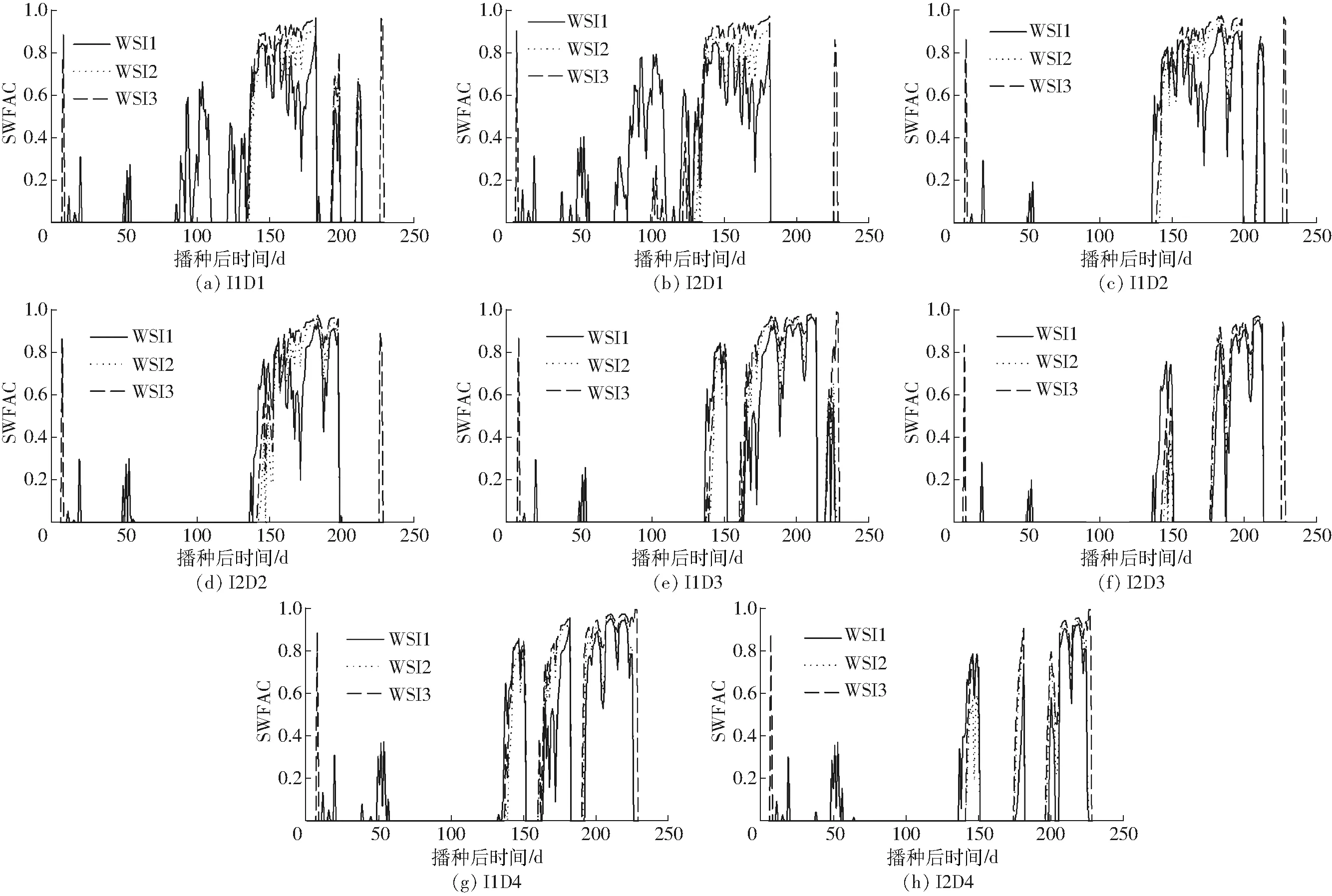
图2 2012—2013年度模拟的不同受旱条件下水分胁迫因子(WSI1、WSI2、WSI3)的动态变化Fig.2 Dynamic changes of SWFAC under different scenarios of water stress in 2012—2013 growing season by using three stress indices of WSI1, WSI2 and WSI3
2.2.2土壤水分状况的模拟结果比较
由于冬小麦根系主要生长分布于20~40 cm,因此本研究主要对比3种水分胁迫因子在该土层的水分模拟状况(图3)。总体来看,3种因子对土壤含水率的动态变化模拟效果较好,RRMSE为7.17%~13.28%。且3种因子模拟的土壤含水率的模拟值变化趋势基本一致,均表现为抽穗期(180 d)之前模拟值低于实测值,此阶段土壤水分的消耗以土壤蒸发为主,间接反映出模型模拟的土壤蒸发偏高;而抽穗期之后模拟值高于实测值,随着作物生长土壤水分主要通过作物蒸腾散失,土壤水分模拟值偏高,反映出模型模拟的作物生长蒸腾量偏低。WSI2和WSI3均采用Nimah and Hanks方程计算根系吸水量,两者模拟的土壤含水率一致,相较于WSI1偏低,作物生长后期差异明显。

图3 2012—2013年度3种不同水分胁迫因子(WSI1、WSI2、WSI3)下各处理土壤含水率(20~40 cm)的模拟效果比较Fig.3 Comparisons of measured and simulated soil moistures (20~40 cm) under different scenarios of water stress in 2012—2013 growing season by using three stress indices of WSI1, WSI2 and WSI3
2.2.3累积蒸腾量的模拟结果比较
从冬小麦累积蒸腾量的模拟值来看,除D1处理外,水分胁迫因子WSI1最高,WSI2次之(图4)。实际蒸腾量间接反映了作物对水分的吸收状况,WSI2和WSI3均采用Nimah and Hanks方程模拟根系吸水量,但两者的模拟值仍有差异。这是由于WSI3考虑了潜热对冠层的影响,通过在SWFAC的分子和分母中加入土壤蒸发项,使模拟的水分胁迫程度偏大,造成作物生长状况较差、蒸腾量较小。与其他处理不同的是,D1处理下WSI2模拟的累积蒸腾量最高,其中I2D1处理下利用胁迫因子WSI1、WSI2、WSI3模拟的2012—2013季冬小麦累积蒸腾量分别为119.5、137.6、101.8 mm。
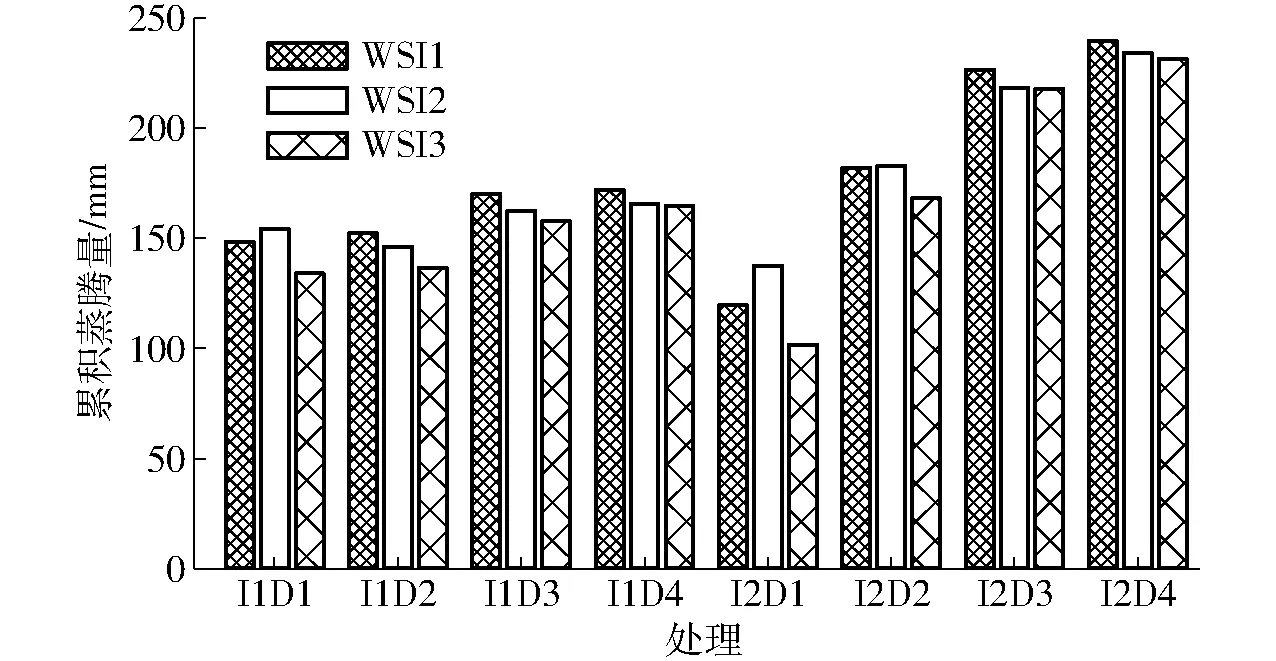
图4 2012—2013年度3种不同水分胁迫因子(WSI1、WSI2、WSI3)下各处理累积蒸腾量Fig.4 Simulated total transpirations under different scenarios of water stress in 2012—2013 growing season by using three stress indices of WSI1, WSI2 and WSI3
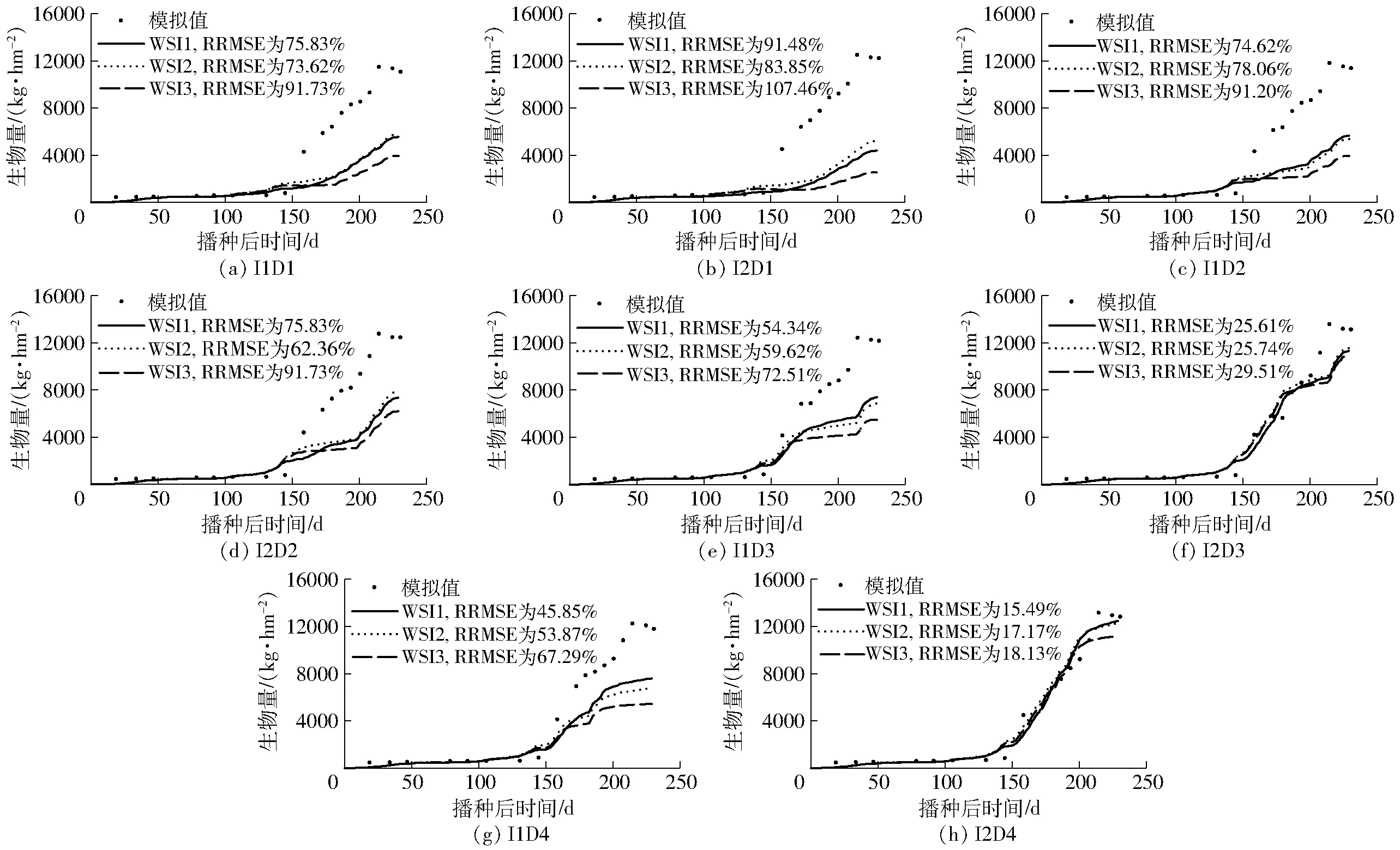
图5 2012—2013年度3种不同水分胁迫因子(WSI1、WSI2、WSI3)下各处理冬小麦生物量的模拟效果比较Fig.5 Comparisons of measured and simulated winter wheat biomass under different scenarios of water stress in 2012—2013 growing season by using three stress indices of WSI1, WSI2 and WSI3
2.2.4作物生长状况的模拟结果比较
从模型对冬小麦生物量的模拟结果来看,3种水分胁迫因子均对高水处理、后期胁迫的处理模拟效果较好,而低水处理、前期受旱的模拟效果较差。且自返青期后(约播后130 d)生物量模拟值均低于实测值,表明3种因子模拟的水分胁迫程度均偏大,对冬小麦生物量的累积抑制作用过强。而三者相比,WSI2模拟的各胁迫处理下冬小麦生物量结果最优,WSI1次之,WSI3较差(图5)。其中D1处理下,WSI2对生物量的模拟较WSI1提高明显,RZWQM2模型对前期受旱冬小麦生长发育的模拟得到改善。
同生物量的模拟结果类似,3种水分胁迫因子对产量的模拟精度均表现出随着受旱程度的加深和受旱时段的提前逐渐降低的趋势。三者相比,WSI2的模拟精度最高,2年平均ARE较WSI1降低了1.43个百分点,其中D1处理的模拟精度改善明显(表5)。WSI2对冬小麦产量的模拟效果得到提高,得益于改由Nimah and Hanks方程计算水分胁迫因子中的潜在根系吸水项,模拟的水分胁迫相较于WSI1历时短,较WSI3程度轻,作物累积蒸腾量最大,因而产量的模拟效果得到提高。D3、D4处理中,低水处理冬小麦产量被低估,WSI1的模拟精度最高,而高水处理下WSI3的模拟精度最高,这是因为3种胁迫因子均高估了冬小麦产量,而WSI3在WSI2的基础上增加了土壤蒸发项,造成模拟的水分胁迫程度增加,使各处理产量的模拟值最接近实测值。

表5 3种水分胁迫因子(WSI1、WSI2、WSI3)对不同受旱情境下冬小麦产量模拟效果比较Tab.5 Comparisons of measured and simulated winter wheat yields under different scenarios of water stress by using three stress indices of WSI1, WSI2 and WSI3
3 讨论
本研究利用RZWQM2模型对2 a冬小麦分段受旱试验进行模拟发现,同一年不同受旱条件下的冬小麦开花期和成熟期都一样且与CK处理相同,而田间试验表明,干旱胁迫发生得越早、胁迫程度越重,冬小麦的开花期和成熟期越提前。孙宏勇等[37]的研究也发现整个生育期干旱胁迫能使冬小麦的生育期缩短5~8 d。许多研究表明,模型无法模拟不同水分胁迫造成的作物物候期的差异,需要对模型进行相应的改进[38-40]。因此,模型对水分胁迫造成的作物物候期变化还需进一步研究。
经校正和验证后,RZWQM2模型可以很好地模拟CK处理下冬小麦的生物量和籽粒产量。而三者模拟的生物量和籽粒产量均表现为低水水平处理的模拟精度低于高水水平,前期受旱处理的模拟精度低于后期受旱处理。因此RZWQM2模型对前期受旱和严重受旱条件下的冬小麦生长过程的模拟还需进一步改进。陈晓远等[41]的研究表明,不同生育期复水对前期遭受水分胁迫冬小麦的生长及产量均有促进作用,复水后作物的生长受到激发,在茎秆伸长的同时绿叶面积增大,干物质积累增加且更多地向冠部分配,根冠比降低,作物进行补偿生长,其中以拔节期复水的补偿作用最大。ACEVEDO等[42]认为,轻度胁迫后复水,植物表现出补偿生长。刘春光[43]的研究表明,分蘖期适应水分胁迫能力强于拔节期,且重度胁迫受旱较中度胁迫严重,但复水后小麦生长及生理反应的补偿效应也相对明显。而本研究中RZWQM2模型对D1处理(越冬期和拔节期受旱,返青期复水)的模拟并没有体现出复水后冬小麦的补偿生长。因此,模型未考虑旱后复水对作物生长的补偿效应,这可能是导致冬小麦前期受旱情境下生物量和籽粒产量被低估的原因之一。
从冬小麦生物量和籽粒产量模拟效果来看,WSI2较WSI1在冬小麦前期受旱处理有所提高;而WSI3对冬小麦后期受旱处理的模拟较WSI1略有改善。整体来看WSI2模拟的各受旱处理下冬小麦的生物量和产量精度最高,其中2 a生物量的平均RRMSE降低了2.84个百分点,籽粒产量的平均ARE降低了1.43个百分点。水分胁迫对作物生物量和产量的影响主要取决于胁迫的轻重和持续时间。WSI2在WSI1的基础上改由Nimah and Hanks方程计算水分胁迫因子中的潜在根系吸水项,改变了水分胁迫历时和程度,模拟的作物累积蒸腾量最大,因而产量和生物量的模拟效果得到提高。而WSI3在WSI2基础上通过在SWFAC的分子分母中加入土壤蒸发项,考虑土壤水分亏缺对冠层温度的影响,进而导致作物遭受水分胁迫。本研究为两个连续生育期持续受旱,受旱时间长,前期受旱处理作物长势差,恢复灌水后水分主要用于土壤蒸发。因此本研究中前期受旱处理下选用WSI3计算出水分胁迫值必然偏大。实际模拟结果也是如此,WSI3模拟的SWFAC较WSI2和WSI1明显增大,生物量和籽粒产量的模拟效果最差。刘恩民等[44]的研究也表明,受旱麦田叶面温度的升高受受旱历时和作物生长状况的影响。长期受旱麦田植株发育较差,同等受旱条件下叶面温度升高值小,而受旱处理前一直正常供水麦田植株发育正常,同等受旱条件下叶面温度升高明显。因此,在今后的研究中需将作物受旱历时及作物生长状况对冠层温度升高导致的水分胁迫考虑进水分胁迫因子的计算中。
3种水分胁迫因子对20~40 cm土层土壤含水率的模拟均呈现抽穗期之前偏低,而抽穗期后偏高的趋势。土壤中水分的散失主要通过蒸发和植株蒸腾进行,这一趋势反映出模型对ET模拟的不准确性。DEJONGE等[45]认为,要改进CERES模型模拟水分胁迫条件下的作物生长过程,应从提高蒸发蒸腾量的模拟精度入手。而从三者模拟的累积蒸腾量来看,蒸腾量是影响水分胁迫条件下作物产量模拟的重要因素。因此,可以考虑通过改进ET的模拟精度提高模型对水分胁迫条件下作物生长过程的模拟。
此外,王艳哲等[46]的研究表明,在水分供应受限制条件下,增施氮肥会降低根冠比,更利于地上干物质的积累和经济产量形成。黄玲等[47]研究表明,在干旱胁迫初期可通过施氮来提高土壤贮水的利用率。孟兆江等[48]研究表明,水分调亏复水后有利于增强作物对土壤氮素的吸收利用能力。由于试验设置处理限制,本研究没有考虑氮素对作物生长发育的影响,在水、氮互作条件下模型对作物生长发育的模拟将会发生变化,还需要进一步研究。
4 结论
(1)现有RZWQM2模型能较好地模拟生育后期轻度受旱条件下冬小麦的生长发育过程,但随着受旱程度的加深和受旱时段的提前,模型的模拟精度降低。
(2)水分胁迫因子WSI2对冬小麦各受旱处理的模拟效果相对较好,突出表现为提高了冬小麦越冬期和返青期受旱条件下生物量、产量的模拟精度。如需利用现有模型对水分胁迫条件下冬小麦生长发育的过程进行模拟,建议水分胁迫因子选用WSI2。
猜你喜欢
中国农业信息(2022年5期)2023-01-14
现代园艺(2022年7期)2022-11-19
中国交通信息化(2022年7期)2022-10-27
林业科技(2022年5期)2022-10-08
长江科学院院报(2022年5期)2022-05-19
农业灾害研究(2022年1期)2022-05-07
今日农业(2021年4期)2021-06-09
今日农业(2020年19期)2020-12-14
安徽农学通报(2020年7期)2020-05-26
湖北农业科学(2019年22期)2019-12-23
