
Developmental competence of bovine oocytes with increasing concentrations of nano-copper and nano-zinc particles during in vitro maturation
2018-07-28 08:50:44BakarAbdelHalimWalaaMoselhyNermeenAtefHelmy
Bakar R Abdel-Halim, Walaa A Moselhy, Nermeen Atef Helmy
1Department of Theriogenology, Faculty of Veterinary Medicine, Beni-Suef University, Beni-Suef, Egypt
2Department of Forensic Medicine and Toxicology, Faculty of Veterinary Medicine, Beni-Suef University, Beni-Suef, Egypt
3Department of Physiology, Faculty of Veterinary Medicine, Beni-Suef University, Beni-Suef, Egypt
Keywords:Nanocopper-oxide particle Nanozinc-oxide particle Toxicity IVM In vitro embryo production Cattle
ABSTRACT Objective: To evaluate copper and zinc concentrations in plasma and follicular fluid from cattle ovaries, and estimate the impact of rational concentrations of copper and zinc oxide nanoparticles (CuO-NPs & ZnO-NPs) supplemented during in vitro maturation (IVM)against DNA damage of cumulus cells, glutathione content in oocytes and on consequent embryo development. Methods: Oocytes were obtained from 2 to 8 mm follicles by aspiration method for IVM. Replicates of experiments were performed on different days,with a separate batch of cumulus oocytes complex for each day. Results: The DNA damage of oocytes and cumulus cells significantly decreased with supplemental CuO-NPs or ZnONPs concentrations of 0.7 and 1.0 µg/mL in the IVM medium compared to medium without CuO-NPs or ZnO-NPs (P < 0.01). Total glutathione concentrations in oocytes and cumulus cells significantly increased following supplementation with both 0.7 and 1.0 µg/mL CuO-NPs or ZnO-NPs in comparison with 0 and 0.4 µg/mL CuO-NPs or ZnO-NPs supplemented groups (P < 0.01). Supplementation of CuO-NPs or ZnO-NPs during IVM medium at any concentration had no significant effect on cleavge rate. Both CuO-NPs and ZnO-NPs significantly increased blastocyst rates when oocytes were matured with 0.7, 1.0 µg/mL CuO-NPs concentrations (P < 0.01). In contrast, addition of 1.5 µg/mL of CuO-NPs or ZnO-NPs to the maturation media resulted in detrimental effects on the developmental competence of bovine oocytes confirming toxicity induced by CuO-NPs and ZnO-NPs in high concentrations.Conclusions: CuO-NPs or ZnO-NPs-treated bovine oocytes during IVM show low level of DNA fragmentation and increased intracellular glutathione content of cumulus cells. In vitro embryo development is improved by supplementation of rational concentrations of CuO-NPs or ZnO-NPs to culture media. Toxicity induced by CuO-NPs and ZnO-NPs is confirmed in high concentrations.
1. Introduction
Nanoparticles are widely used in various fields including reproduction and immunity due to their physical characters[1,2]. The metal nanoparticles have an effective antioxidant activity and bactericidal effect due to its nanometer small size and surface/volume ratio. The nature and mechanism of nano copper (Cu) biotoxicity allows for an explanation of the biological effects of nanosized metal admixtures in the composition of other nanomaterials, including carbon based nanomaterials[3]. Cu activates dopamineβ-monooxygenase through catalyzing hydroxylation of dopamine to an essential neurotransmitter nor adrenaline, which stimulates the secretion of gonadotropin releasing hormone that controls both luteinizing hormone and follicle-stimulating hormone secretion from the anterior pituitary under effect of gonadotropin releasing hormone[4,5]. Cu lowers endopeptidase pass to bonds of specific amino acid, resulting in GnRH alteration[6]. Cu deficiency increased DNA fragmentation as a result of free radicals production[7,8]. Both iron (Fe) and Cu improves the culture of embryo developmental stages; a deficiency of Fe or Cu resulted in high apoptotic blastomeres[9]. Sufficient zinc (Zn) can improve cellular growth duringin vitroembryo culture[10,11]; Zn constitutes a component of enzymes incorborated in both ova, embryo developmental stages[10]. Zn is considered as a co-factor of proteins bound DNA[12]. Mitochondrial Zn is released in a redox reaction by oxidized form of glutathione (GSH-GSSG)[13,14].Zinc oxide nanoparticles (ZnO-NPs) resistance to heat and radiation are the main causes which expand their vast applications[15]. The current study aimed to detect Cu and Zn levels in follicular fluid and to monitor the effect of various concentrations of copper oxide nanoparticles (CuO-NPs) and ZnO-NPs supplemented during maturation and embryo growth. Recently, higher concentrations of nanoparticles (NPs) have been found to cause a detrimental effect on bovine oocytes developedin vitro[16]. Toxic symptoms in male rats exposed to CuO-NPs were observed at a higher rate than females exposed to the same particle mass[17]. It has been observed that higher concentrations of both copper oxide (CuO) and CuONPs—treated oocytes reduce glutathione (GSH) content, and impair catalytic activity and superoxide dimutase, which leads to embryo oxidative damage and physiological changes in zebra fish, especially hatching failure, shortening body length, and low reproductive efficiency. There was a significant reduction in the cleavage rates in embryos subjected to 1-25 mg/dm3CuO-NPs[18].
2. Materials and methods
2.1. Chemicals
All required chemicals were from Sigma-Aldresh. Tissue culture media-199 (TCM-199) was used as maturation medium. The fertilization medium was total alkaline phosphatase media prepared as described previously by Parrish[19]. Modified synthetic oviduct fluid was used as a developmental media for embryo culture[20].
2.2. Preparation of nanoparticles
CuO-NPs were prepared as mentioned by Ramyadeviet al[21]and ZnO-NPs were synthesized as described by Ghorbaniet al[22].Characterization of both CuO-NPs and ZnO-NPs was done by using transmission electron microscope.
2.3. Recovery of oocytes
Ovaries were collected randomly from local abattoir in Beni-Suef Governorate, Egypt and transported to the laboratory in sterile phosphate buffer saline (PBS) with antibiotics (streptomycin)at 37 ℃ within 1 h. Cumulus oocytes complexes (COCs) were harvested by slicing method. Oocytes with intact cumulus cells with granular cytoplasm were collected by using a stereomicroscope.Four to six replicates were conducted all over the period of the experiment.
2.4. Maturation of oocytes in vitro
TCM having 5% fetal calf serum was used for washing the oocytes complex twice. The oocytes were then washed in the media of maturation, divided into groups with 10 cells of each, transported toin vitromaturation (IVM) (50 µL) covered by mineral oil, cultured at 39 ℃ in 5% CO2incubator for 24 h, and then examined for cumulus expansion. Expanded oocytes were selected for fertilisation.
2.5. Comet assay
The procedure and classification of the cells were done according to Collins[23]. Cells were classified according to their tail length into five grades, namely, Grade 0 (comet with no visible tail), Grade 1(comets showing tiny tail), Grade 2 (including comets with a dim tail), Grade 3 (involving comets showing a clear tail), and Grade 4(comets with a reduced diameter of the head and showing a clear tail)
2.6. GSH-GSSG assay
The GSH-GSSG assay was carried out according to Krisher[24].GSH content within the cells was measured from a standard curve of GSH.
2.7. Fertilization and culture of matured oocytes in vitro
Motile spermatozoa were selected using swim-up technique (Parrish 2014). Frozen semen was obtained from Artificial Insemination Centre, Beni-Suef Governorate, Egypt. 2 × 106spermatozoa/mL was the final sperm concentration inin vitrofertilization, then incubated at 39 ℃ in 5% CO2for 24 h. After fertilization, denuded zygotes were washed twice in PBS, then transported to 25 µL modified synthetic oviduct fluid (1 embryo/µL) supplemented with fetal calf serum (10%, v/v) under mineral oil. The dishes were incubated at 38.5 ℃ in a 5% CO2for 8 days[25]. Cleavage rates were recorded at 48 h post-insemination and blastocyst number was determined on day 8 post-insemination.
2.8. Blastocyst staining for total cell number
At day 8 (day 0 = day of fertilization), blastocysts were washed twice with 1% polyvinyl pyrrolidone in PBS, and then fixed with 4%formaldehyde. The blastocysts were remained in 1% Triton, stained with Hoechst 33342 overnight (2.5 µg/50 µL mSOF) on slides, and then covered with a cover slip. The number of nuclei of blastocysts cells (8 d, Grade 1)(Grade Ⅰ: blastocyst with evenly granulated cytoplasm and intact cell membrane; Grade Ⅱ: blastocyst with uneven granular cytoplasm and intact cell membrane; Grade Ⅲ: blastocysts that were degenertated) could be counted under an epifluorescent microscope (Nikon; Tokyo, Japan).
2.9. Estimation of Cu concentrations
Blood (10 mL) were collected from jugular vein from cows(few minutes prior to slaughter), then the samples underwent centrifugation at 15 000 rpm for 10 min for separation of plasma.At the same time, ovaries of these cows were harvested randomly to estimate Cu concentration in different classes of follicles. An insulin syringe was used to aspirate follicular fluid from each follicle. Samples were pooled (usually between 7 and 10 small sized follicles) within a given ovary. Concentration of Cu was estimated by spectrophotometer (GBC 902)[26].
2.10. Experimental design
2.10.1. DNA integrity with CuO-NPs or ZnO-NPs treated oocytes during IVM medium
In experiment 1, the impact of CuO-NPs or ZnO-NPs on DNA fragmentation of oocytes following the addition of 0, 0.4, 0.7, 1.0 or 1.5 µg/mL CuO-NPs or ZnO-NPs to IVM medium was evaluated.DNA damage was evaluated in the matured COCs. In this regards,three replicates of 450 oocytes from different days were maturedin vitrowith increasing concentrations of CuO-NPs and in a similar manner for ZnO-NPs.
2.10.2. Intracellular GSH level with CuO-NPs or ZnO-NPs treated oocytes during IVM medium
Experiment 2 was conducted to detect the impact of adding 0, 0.4,0.7, 1.0 or 1.5 µg/mL Cuo-NPs or ZnO-NPs to IVM on GSH-GSSG levels in cumulus cells. After maturation of oocytes for 24 h, the total GSH concentrations were investigated. In this regards, four replicates of oocytes (grade 1) from different days, with 250 oocytes, 50 cells per treatment in each replicate (oocytes’ total number of grade 1:n=1 000) were maturedin vitrowith increasing concentrations of CuO-NPs. Similar protocols were conducted for ZnO-NPs.
2.10.3. Effect of CuO-NPs or ZnO-NPs during IVM on embryo developmental stages
Experiment 3 was designed to follow up the growth of cleaved embryos from matured oocytes; IVM medium treated with 0, 0.4,0.7, 1.0 or 1.5 µg/mL CuO-NPs or ZnO-NPs was investigated. In this respect, 1 000 COCs were matured in four replicates with CuONPs; the same number was used for ZnO-NPs. Cleavage rates were investigated 48 h post- insemination. Development to the blastocyst stage was monitored, either from oocytes or cleaved embryos.
2.11. Statistical analyses
Data in the current study were analyzed by using one way analysis of variance (ANOVA) and Student-Newman-Keuls multiple comparisonpost-hoctest (Tukey’s test), using SPSS (Version 16;SPSS Inc., Chicago, IL, USA). Data were expressed as Mean ±standard error (Mean ±SE).
3. Results
3.1. Cu concentration in plasma, follicular fluid, and IVM medium
Cu concentrations were (1.12±0.21) µg/mL Cu in 20 plasma samples, (0.88±0.17) µg/mL in follicular fluid from large follicles,while the concentration of Cu in follicular fluid from small follicles was (0.77±0.19) µg/mL, and (0.07±0.01) µg/mL in the maturation medium. Cu concentrations in plasma and follicular fluid had no significant difference (P> 0.05).
3.2. Zn concentration in plasma, follicular fluid, and IVM medium
In plasma, Zn level was (1.26±0.70) µg/mL, while in follicular fluid from large follicles the Zn concentration was (1.38±0.22)µg/mL compared to (1.42±0.41) µg/mL in follicular fluid from small follicles, while Zn concentration was (0.20±0.01) µg/mL in maturation medium. Zn concentrations in plasma and follicular fluid had no significant difference (P> 0.05).
3.3. Effects of CuO-NPs or ZnO-NPs on integrity of DNA in cumulus cells
The damage degree of DNA in oocytes cultured with 0.7 and 1.0 g/mL CuO-NPs during IVM showed a significant (P< 0.01)decrease. Damage was lower in oocytes matured in medium with 0.7 or 1.0 µg/mL CuO-NPs compared with those matured in the medium without CuO-NPs. However, damage degree of DNA rose again with increased concentration of CuO-NPs (1.5 µg/mL), which confirmed toxicity was induced by higher concentrations of CuONPs in oocytes during IVM (Table 1).
The damage degree of DNA in oocytes cultured with 0.7 and 1.0 µg/mL ZnO-NPs during IVM showed a significant (P< 0.01)decreas. The oocytes matured in 0.7 or 1.0 µg/mL ZnO-NPs showed lower damage compared with those matured in the medium without ZnO-NPs. However, damage degree of DNA rose again with increased concentration of ZnO-NPs (1.5 µg/mL), which confirmed toxicity was induced by higher concentrations of ZnO-NPs in oocytes during IVM (Table 2).
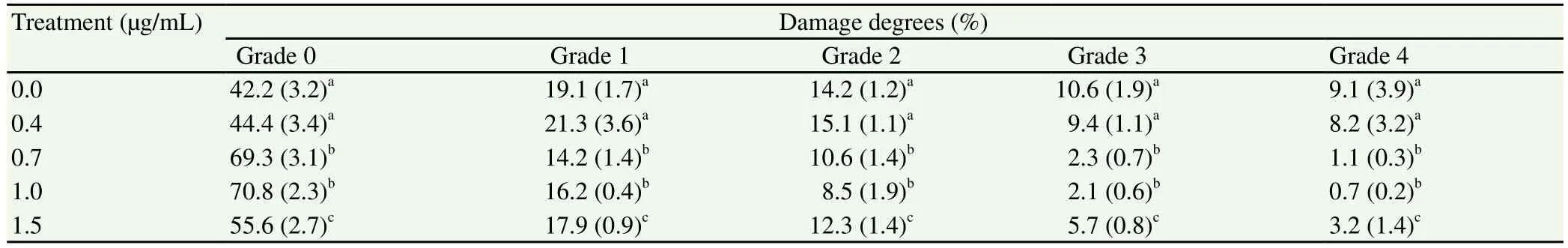
Table 1 DNA fragmentation in cumulus cells during IVM with various concentrations of CuO-NPs.
3.4. Effects of CuO-NPs or ZnO-NPs on intracellular GSH concentration
Intracellular concentrations of GSH-GSSG were significantly(P< 0.01) increased with the addition of 0.7 and 1.0 µg/mL CuONPs. On the contrary, the total GSH decreased in oocytes and cumulus cells supplemented with 1.5 µg/mL confirming toxicity exerted by higher concentrations of CuO-NPs during IVM (Table 3).Moreover, in the same experiment intracellular concentrations of GSH-GSSG were significantly (P< 0.01) increased with 0.7 and 1.0µg/mL ZnO-NPs. However, GSH-GSSG concentration did not differ significantly with 0 or 0.4 µg/mL ZnO-NPs. Besides, total GSH decreased in oocytes and cumulus cells supplemented with 1.5 µg/mL,which confirmed toxicity was exerted by higher concentrations of ZnO-NPs during IVM (Table 3).
3.5. Effects of CuO-NPs or ZnO-NPs on the developmental stages of embryos
The results revealed that addition of CuO-NPs to IVM at any concentration had no significant difference in cleavage rate. CuONPs significantly (P< 0.01) increased blastocyst rates when oocytes were matured with 0.7, 1.0 µg/mL CuO-NPs concentrations. At the same time, CuO-NPs significantly increased the percentages of blastocyst calculated from cleaved embryos (P< 0.01) (Table 4).
In the same experiment, the results showed that there was no difference in cleavage rate when ZnO-NPs were added to IVM medium. While, there was an significant (P< 0.01) increase in the blastocyst rate in oocytes with 0.7 and 1.0 µg/mL ZnO-NP (Table 4).In contrast, supplementation of IVM with 1.5 µg/mL CuO-NPs or ZnO-NPs resulted in significant decrease in cleavage and blastocyst rates.
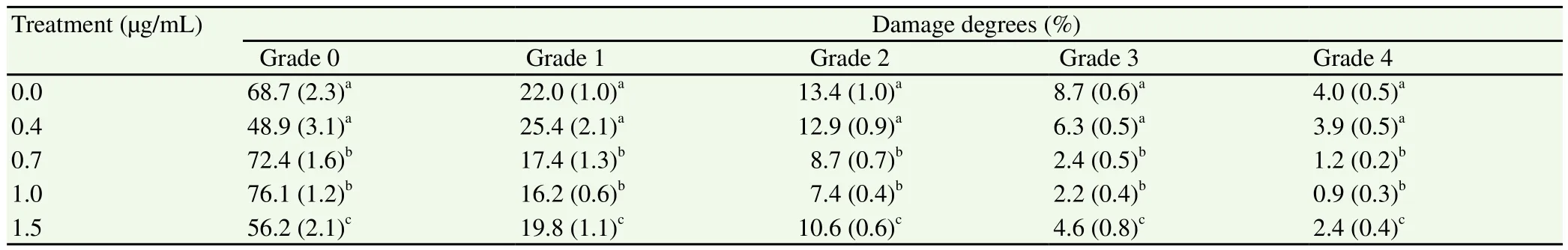
Table 2 DNA fragmentation in cumulus cells matured in vitro with various concentrations of ZnO-NPs.

Table 3 Total GSH concentrations in oocytes and cumulus cells during IVM with increasing concentrations of CuO-NPs and ZnO-NPs.

Table 4 Embryo development from oocytes during IVM with increasing concentrations of CuO-NPs and ZnO-NPs.
4. Discussion
In the present study, both Cu and Zn concentrations in follicular fluid and plasma of cows were similar. The addition of Cuo-NPs or ZnO-NPs during IVM of bovine oocytes minimized DNA fragmentation and increased total GSH content in oocytes and cumulus cells when supplemented at 0.7 and 1.0 µg/mL. In addition,when the Cuo-NPs or ZnO-NPs concentration was 0.7 and 1.0 µg/mL during IVM, the developmental competence (embryo development and embryo quality) had improved.
Cu status in cattle is defined as deficient, marginal, and adequate for plasma concentrations of ≤ 30, 31—60, and ≥ 60 µg/dL Cu respectively[27]. In the present study, Cu concentrations in cows were similar in plasma and follicular fluid from large and small follicles. Therefore, plasma Cu status might be useful to predict Cu status in the follicular environmentin vivo. Concentrations of Cu,Zn, and selenium in serum and follicular fluid were decreased in women subjected toin vitrofertilization, which could be adjusted by giving mineral supplements, which had a positive impact on lipid peroxidation, reduced glutathione in serum and follicular fluid,augmenting the antioxidant defense system by decreasing oxidative stress[28].
In mammalian cells, Cu and Zn operate as a free radical scavenger.Addition of Zn sulfate to cell culture can protect against DNA damage and prevent cell deathin vitro[29]. The antioxidant role of CuO-NPs and ZnO-NPs preserves DNA integrity in oocyte by inhibiting DNA fragmentation. In the current study, IVM medium with 0 or 0.4 µg/mL CuO-NPs or ZnO-NPs had no protective effect against the DNA damage of the cells. However, DNA integrity in oocytes was improved when CuO-NPs or ZnO-NPs were added to IVM medium at 0.7 and 1.0 µg/mL concentrations.
The increase of DNA damage found in cows with Cu deficiency is attributed to higher oxidative stress[30]. Cu deficiency in cattle increased the frequency of chromosomal aberrations and DNA migration. Intracellular glutathione concentration (GSH-GSSG)was increased in 0.7 and 1.0 µg/mL CuO-NPs or ZnO-NPs IVM supplemented media, in both oocytes and cumulus cells. It has been recorded that reduction of intracellular Cu or Zn resulted in a reduction in GSH level in Chinese hamster ovary cells culturedin vitro[31].
The relation between CuO-NPs or ZnO-NPs and GSH has been explained; Cu is a biological ligand forming binary complexes with GSH[32]. Moreover, formation of a stable Cu-GSH complex to the presence of some Cu-chelators lacks all thiol reactivity, but fully serves the free-radical scavenging properties of GSH[32]. Higher concentrations of CuO-NPs or ZnO-NPs is toxic in its unbound form, and causes redox imbalance attributed to its highly redox active nature[33].
Furthermore, high GSH store during IVM improved developmental competence of bovine oocytes, producing more embryos and reaching the blastocyst stage[34]. In the current study, regardless of Cu or Zn status in the animal, various concentrations of Cuo-NPs(0.4, 0.7 and 1.0 µg/mL) supplemented to IVM medium and the embryo development of these oocytes till the blastocyst stage did not increase cleavage rates. While, ZnO-NPs (0.4, 0.7, and 1.0 µg/mL) added to IVM medium were significantly increased cleavage rates. However,the blastocyst rate was significantly increased when oocytes were matured with CuO-NPs or ZnO-NPs (0.4, 0.7, and 1.0 µg/mL)during IVM. Furthermore, the blastocyst rate was significantly higher when 0.7 and 1.0 µg/mL CuO-NPs or ZnO-NPs were used.These results were in agreement with a previous report that Cu during IVM had an effect on bovine oocyte maturation[9]. However,in our study, the improvement of blastocysts rates was a consequence of the addition of CuO-NPs during oocyte maturation, whereas Gaoet al[9] found that the supplementation of zygote medium with Cu improved embryo development. Furthermore, the addition of 0.68 and 0.46 mg/L Cu to zygote culture media improved the yield of bovine embryo[9].
On the contrary, the current study revealed that higher concentrations of CuO-NPs or ZnO-NPs (higher than 1.0 µg/mL) are toxic to oocytes maturedin vitroand deteriorate embryo development.Several mechanisms have been suggested to explain the toxic effect of Cu in living cells; Cu acts as catalyst in the production of superoxide radicals, hydroxyl radicals and hydrogen peroxide (Haber-Weiss reaction) leading to oxidative damage[35]. High concentrations of CuO-NPs or ZnO-NPs- supplemented oocytesin vitrolead to high DNA fragmentation. Recently, it has been found that CuO-NPs are toxic to skin-associated cells and that extracellular signal-regulated kinase and p53 is the key factors controlling the mechanism of cytotoxicity[36]. CuO-NPs also induced apoptosis and oxidative damage in HaCaT human keratinocytes[37].
In conclusion, Cu and Zn levels were similar in plasma and follicular fluid of cows. Antioxidant parameters which reflect the antioxidant role of CuO-NPs and ZnO-NPs supplementation duringin vitromaturation of bovine oocytes had been proved.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Acknowledgments
Our sincere gratitude goes to Department of Nanotechnology,Faculty of Science, Beni-Suef University, for their kind supervision and helpfulness during this study and for their kind cooperation,faithful advice and careful revision of this work.
 Asian Pacific Journal of Reproduction2018年4期
Asian Pacific Journal of Reproduction2018年4期
- Asian Pacific Journal of Reproduction的其它文章
- In vitro fertilization: Facts in medical sciences
- Functions of follicular and marginal zone B cells in pregnancy
- Treatment of cows with clinical endometritis Ⅲ as cows affected by pyometra-Non antibiotic treatment of severe clinical endometritis
- Effect of foot and mouth disease vaccination on seminal antioxidant profiles of mithun(Bos frontalis)
- Consequences of unilateral cryptorchidism on semen and sperm characteristics in West African Dwarf Goats
- Increase in reproductive ability of high-producing cows, and qualitative parameters of their offspring, under conditions of intensive milk production