
中国季节性昼夜增温的不对称性及其对植被活动的影响
2018-07-18 10:10杜自强张霄羽
生态学报 2018年11期
赵 杰, 杜自强,*, 张 红, 张霄羽, 赵 虹
1 山西大学黄土高原研究所, 太原 030006 2 山西大学环境与资源学院, 太原 030006
植被作为地球系统物质循环和能量流动的主要载体,在调节全球碳平衡、降低温室气体浓度和维持气候稳定方面扮演着不可替代的角色[1- 2]。归一化植被指数(Normal difference vegetation index, NDVI) 对地表植被覆盖状况反映敏感,是监测和指示植被活动和生产力变化情况的常用指标[3- 5]。植被的诸多生理生态特征,如光合作用能力、生长周期的持续时间以及群落变化等对气候变化反映敏感[6- 7],植被动态变化及其对气候因子变化的响应研究成为研究全球变化的关键问题之一[8- 9]。目前,越来越多的证据表明全球气候变暖普遍存在昼夜不对称(即增温速率不一致)以及季节不对称的趋势特征[10- 12]。例如,全球大部分地区夜间增温速率快于白天增温速率[10],北半球中高纬度地区普遍存在夏季增温速率快于春季和秋季的趋势特征[12]。由于大多数植物的光合作用在白天进行,而呼吸作用贯穿整天,昼夜增温速率的差异势必对植被的碳吸收和碳消耗产生重要影响[13- 15]。同样,在温带地区,植被的物候特征和光合作用特性具备明显的季节性周期,植被活动对温度的响应在不同季节存在差异[15- 17]。因此,从季节尺度上研究昼夜不对称增温与植被动态的相关性有助于厘清全球变化背景下增温对植被的影响。
近年来,国内外部分学者基于气象站点数据及卫星观测数据在宏观尺度上开展了不同季节昼夜增温对植被活动影响的相关研究。例如,Shen 等[18]发现,夏季白天增温利于青藏高原地区的植被生长,而该季节夜间温度的上升对植被的生长起到消极作用。Tan 等[7]在季节尺度上分析北半球中高纬度地区植被对不对称昼夜增温的响应时发现,在春季和夏季夜间增温利于植被生长,秋季夜间温度的升高对大多数地区的植被生长不利;在夏季,白天增温不利于温带干旱地区植被的生长,但春季白天增温却能够利于寒带地区植被NDVI的提高。Shen 等[19]的研究结果同样表明,不同季节的昼夜增温对北半球中高纬度地区植被光合能力的影响程度各有不同。尽管部分学者通过遥感数据和气象数据分析了北半球植被对昼夜增温的季节性响应特征[7, 18-19],但是关于季节性昼夜不对称增温对植被活动的影响仍旧缺乏全面的认识。因此,有必要在基于遥感数据和气象站点数据研究的基础上,结合昼夜模拟增温对植被生长影响的相关结果,继续深入探索季节性昼夜不对称增温对植被活动的影响。
中国位于欧亚大陆的东部,受东部季风气候与西北干旱气候的强烈影响,是全球气候变化最为敏感和复杂的地区之一[20],气候变化对我国植被动态的影响十分显著[21- 22]。另外,我国地域面积广大、地形复杂多样,水热条件悬殊,各地区植被对气候变化的响应程度和响应时间存在明显的区域差异,适于进行大尺度、长时间序列的植被对气候变化响应关系研究[1, 23]。基于上述认识,本文利用近34年新一代NDVI数据集以及中国植被区划数据、气象数据,辅以线性回归分析、GIS空间分析等手段,分析并探讨中国季节性昼夜增温的变化趋势及其对不同类型植被活动的影响,以期增强全球气候变化背景下季节性昼夜不对称增温对中国陆地植被动态影响的认识。
1 数据与方法
1.1 数据来源
1.1.1遥感数据
植被NDVI数据来自美国国家航空航天局的全球观测模拟与制图研究组生产的第3代遥感数据(GIMMS-NDVI3g),该数据集由美国宇航局艾姆斯生态预测实验室提供(http://ecocast.arc.nasa.gov/data/pub/gimms/3g.v1/)。 其空间分辨率为8km,时间分辨率为15d,时间跨度是1982年1月—2015年12月。该数据集是目前时间序列最长的NDVI数据集,已被广泛应用于大区域尺度的植被动态变化[24- 25]、植被净初级生产力评估[26]以及生物量估测等研究中[4]。在数据处理过程中,采用最大值合成法重建中国各月NDVI数据集,以进一步消除云层、大气等噪声的干扰[21]。由于NDVI值在低植被覆盖度地区受下垫面的影响较大,借鉴相关研究[1, 25],本文将多年NDVI年均值大于0.1的区域定义为植被区域。
1.1.2气象数据
1982—2015年中国月极端最高气温(Tmax)、极端最低气温(Tmin)、降水量等气象数据来源于中国气象科学数据共享服务网的中国地面气候资料月值数据集。通过ArcGIS10.2,采用反距离权重内插法(Inverse Distance Weighted, IDW)对各个气象因子进行空间插值,获得整个研究区的气象数据和栅格图像[24]。结合相关研究[27],将3—5月、6—8月、9—11月及12—次年2月分别定义为春季、夏季、秋季和冬季,并且将各个季节中的气候要素和NDVI季均值作为该季节的计算数据。
1.1.3植被区划数据

图1 中国植被区划分区图Fig.1 Spatial distribution of studied sub-regions across China R1:寒温带针叶林区域;R2:温带草原区域;R3:温带荒漠区域;R4:青藏高原高寒植被区域;R5:暖温带落叶阔叶林区域;R6:温带针叶、落叶阔叶混交林区域;R7:亚热带常绿阔叶林区域;R8:热带季风雨林、雨林区域
植被区划数据来源于中国科学院资源环境科学数据中心。该数据依据《1∶100万中国植被图集》及各植被类型区的地理分布特征将中国划分出8个彼此有区别,但内部有相对一致性的植被组合分区。据此植被区划数据,本研究将中国植被划分为8大植被分区(图1):寒温带针叶林区域(R1),温带草原区域(R2),温带荒漠区域(R3),青藏高原高寒植被区域(R4),暖温带落叶阔叶林区域(R5),温带针叶、落叶阔叶混交林区域(R6),亚热带常绿阔叶林区域 (R7),热带季风雨林、雨林区域(R8)。
由于除亚热带和热带植被以及少数人工植被以外,其他植被在冬季几乎停止生长,因此,在研究冬季昼夜气温的变化趋势以及NDVI和昼夜气温之间的相关性时仅考虑亚热带常绿阔叶林区域和热带季风雨林、雨林区域。
1.2 研究方法
1.2.1一元线性趋势分析法
在区域尺度和像元尺度对昼夜气温和相应年份进行最小二乘法线性回归分析,以线性回归系数来表征昼夜气温的变化趋势。线性回归系数的显著性检验采用t检验,其中:P<0.05表示回归系数显著,P<0.01表示回归系数极显著。通过逐像元计算各季节昼夜温度变化速率,分析中国植被覆盖地区在不同季节昼夜温度变化趋势的时空格局。
1.2.2偏相关分析法
通过偏相关分析法,可消除其他变量的干扰来研究昼夜增温的不对称性对植被NDVI动态变化的影响[27]。本文通过基于像元的空间分析法,逐像元计算NDVI和Tmax、Tmin的二阶偏相关系数。其中,通过控制Tmin和降水量计算各季节NDVI与Tmax的偏相关系数;控制Tmax和降水量计算各季节NDVI和Tmin的偏相关系数。采用t检验法进行偏相关系数的显著性检验,其中,P< 0.05表示相关性显著,P< 0.01表示相关性极显著。
2 结果与分析
2.1 季节性昼夜增温的时空格局
2.1.1区域尺度
通过线性回归方程计算各个季节昼夜增温的变化趋势后发现(表1):Tmax在4个季节均表现为显著上升,其中春季(0.61℃/10a)和冬季(0.49℃/10a)上升速率最快,秋季(0.35℃/10a)上升速率最慢;Tmin在各个季节同样均表现为显著增长趋势,其中秋季(0.48℃/10a)和夏季(0.43℃/10a)增长速率最快,冬季(0.29℃/10a)增长速率最慢;从昼夜增温速率的对比来看,冬季和春季Tmax的增长速率分别为Tmin增长速率的1.69倍和1.49倍,而秋季和夏季Tmin的增长速率分别为Tmax增长速率的1.35倍和1.04倍。总的来说,昼夜增温在不同季节均表现出不对称变化特征,且不对称变化特征存在明显的季节性差异,其中春季和冬季白天增温速率快于夜间,秋季和夏季夜间增温速率快于白天。
由于各植被分区存在纬度、海拔、海陆位置等存在较大差异,其昼夜增温趋势可能存在显著不同。为进一步理清我国昼夜增温的季节性差异,对不同植被分区在不同的季节进行了昼夜增温趋势的计算。由表1可见,各植被分区在各个季节上昼夜增温速率存在较大差异。例如,春季,R3、R5、R7白天增温快于夜间,R4、R8夜间增温快于白天;夏季,R2、R5、R7白天增温快于夜间,R3、R4、R8夜间增温快于白天;秋季,R3、R5、R7白天增温快于夜间,R4、R8夜间增温快于白天;冬季,R7及R8均表现出白天增温快于夜间增温的趋势。由此可见,各植被分区在各季节均表现出昼夜增温的不对称性,但不同植被分区季节性昼夜增温速率的不对称性存在明显差异。

表1 各季节Tmax和Tmin变化趋势统计/(℃/a)
注:R1:寒温带针叶林区域; R2:温带草原区域; R3:温带荒漠区域;R4:青藏高原高寒植被区域;R5:暖温带落叶阔叶林区域;R6:温带针叶、落叶阔叶混交林区域;R7:亚热带常绿阔叶林区域;R8:热带季风雨林、雨林区域。“研究区”在春季、夏季和秋季指除台湾省以外的整个中国植被覆盖区,冬季只涉及除台湾省以外的R7,R8地区;*表示通过P<0.05显著性检验,**表示通过P<0.01显著性检验
2.1.2像元尺度
空间平均值可以从整体上表征昼夜气温的变化趋势,但由于气温变化特征往往存在空间异质性[27],甚至不同区域的气温可能存在相反的变化趋势,所以基于空间平均值得到的结果不能很好地描述不同区域的变化特征[21]。鉴于此,本文采用最小二乘法线性回归模型,逐像元进行昼夜增温趋势的计算,并对所得结果进行显著性检验。结果如下:
春季,中国大部分地区昼夜气温均呈现上升态势(图2)。其中,Tmax呈现显著上升趋势的地区约占中国面积的68.36%,主要位于新疆北部、青海西部、内蒙古高原中部、黄土高原、东北平原南部、华北平原、长江中下游平原及东南沿海地区。Tmax呈现显著下降的地区面积较小,仅占中国的0.05%。约52.65%的区域Tmin呈现为显著上升趋势,其主要位于新疆西部、青藏地区、华北平原西部、云贵高原、四川盆地及江南丘陵地区。与Tmax类似,Tmin呈现显著下降的区域面积较小,仅占中国的0.21%。

图2 春季Tmax、Tmin增温速率分布图Fig.2 Spatial patterns of warming rate of Tmax and Tmin in spring
与春季类似,夏季中国大部分地区昼夜气温均呈现上升态势(图3)。其中,约有67.20%的地区Tmax呈现显著上升趋势,主要集中在新疆北部、青藏高原东部、内蒙古高原中西部、大兴安岭、华北平原南部以及我国南方地区的大部分区域。Tmax呈现为下降趋势的地区较少,通过显著性检验的地区仅占研究区的0.51%。Tmin呈现为显著上升的地区占研究区的比值高达73.16%,主要分布于西北地区西北部、青藏地区、四川盆地、内蒙古中部、东北平原北部、华北平原西部以及我国东部和南部的沿海地区。Tmin呈现为显著下降趋势的地区仅占研究区的0.22%,且集中分布在黑龙江西北部。
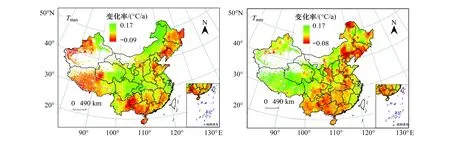
图3 夏季Tmax、Tmin增温速率分布图Fig.3 Spatial patterns of warming rate of Tmax and Tmin in summer
秋季昼夜增温的空间分布(图4)显示,Tmax呈现为显著上升趋势的地区占研究区的41.31%,主要位于青藏高原及我国西南地区,Tmax呈现下降趋势的地区仅占研究区的0.03%。Tmin呈现上升趋势的地区占研究区的57.84%,该地区主要位于新疆西部及40°N以南的大部分地区。Tmin呈现显著下降的地区较少,仅占研究区的0.27%。

图4 秋季Tmax、Tmin增温速率分布图Fig.4 Spatial patterns of warming rate of Tmax and Tmin in autumn
冬季昼夜增温的空间分布(图5)显示,Tmax呈现为显著上升趋势的地区占研究区的55.92%,其主要位于汉中盆地、四川盆地、云南高原、滇南谷地丘陵及西藏东部地区。Tmax呈现下降趋势的地区仅占研究区的0.05%。Tmin呈现显著上升趋势的地区占研究区的36.51%,该地区主要位于西藏东部、四川、云南及浙江中部地区,Tmin呈现为显著下降趋势的地区同样较少,仅占研究区的0.09%。
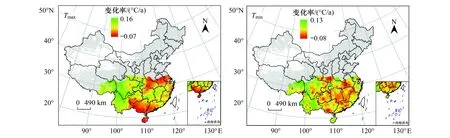
图5 冬季Tmax、Tmin增温速率分布图Fig.5 Spatial patterns of warming rate of Tmax and Tmin in winter
2.2 植被NDVI同季节性昼夜温度的相关性
2.2.1区域尺度
从植被NDVI与不同季节昼夜气温的偏相关系数(表2)来看,整个研究区各季节植被NDVI与Tmax的二阶偏相关系数均为正值。其中,春季和冬季NDVI值与Tmax呈现显著正相关(P﹤0.05)。而NDVI与Tmin的偏相关系数在各个季节均未通过显著性检验(P﹥0.05)。这表明,相比于夜间增温,春季和冬季的白天增温对植被NDVI值的影响更显著,且更多的表现为积极影响。
由于不同类型植被的生物学特性存在差异,可能导致不同植被类型对气候变化的响应存在差异[1,3-4,28]。因此本文对8个植被分区NDVI值与昼夜增温的相关性进行计算,以了解不同植被类型对昼夜不对称增温的响应。不同类型植被与季节NDVI和Tmax、Tmin的偏相关系数(表2)显示:季节性昼夜增温对各类型植被产生不同的影响程度不同。春季,R1、R2、R4、R5、R6、R7地区的植被NDVI与Tmax表现为显著正相关。各类型植被NDVI同Tmin之间的相关性水平较弱,仅R3地区植被NDVI与Tmin表现为显著正相关;夏季,R7地区植被NDVI与Tmax表现为极显著正相关,R1、R3和R8地区植被NDVI与Tmax表现为显著正相关。各类型植被NDVI同Tmin之间的相关性程度较弱,仅R3地区植被NDVI同Tmin通过显著性检验;秋季,R1、R3和R6地区植被NDVI与Tmax之间的相关性通过显著性检验,且皆为正值,R1和R5地区植被NDVI与Tmin的相关性通过显著性检验,前者显著负相关,后者显著正相关;冬季,R7地区植被NDVI与Tmax呈现显著正相关,植被NDVI与Tmin之间的相关性较弱,未能通过显著性检验。
2.2.2像元尺度
为了解研究区不同季节植被NDVI与昼夜气温相关关系的空间格局,通过逐像元计算偏相关系数,得到各个季节植被与昼夜气温的相关性空间分布(图6—9)。
春季植被与昼夜增温的偏相关系数空间分布(图6)显示,约85.90%的地区NDVI与Tmax呈现正相关,其中通过显著性检验的地区占研究区的37.51%,主要分布于青藏高原东部、四川盆地、大兴安岭及东北平原以东地区。NDVI同Tmax呈现显著负相关的地区仅占研究区的0.98%,主要分布在晋南关中盆地及新疆西北部的部分地区。约63.58%的地区NDVI同Tmin呈现正相关,但通过显著性检验的比值仅为8.33%,该地区主要分布在海南、江南丘陵、东北地区北部及陕甘宁地区。NDVI与Tmin呈现为显著负相关的地区占研究区的比值为4.09%,其主要位于青藏高原地区。总体上,春季昼夜温度的上升对我国大部分地区的植被生长均表现出积极的影响,分析原因可能与温度上升导致该地区植被生长季普遍提前有关[29]。进一步发现,NDVI与Tmax呈现为显著正相关的地区远高于NDVI与Tmin呈现为显著正相关的地区。其原因可能是相比于夜间增温,白天增温更易促使植被展叶期、返青期的提前[30]。

表2 各季节植被NDVI与Tmax、Tmin的二阶偏相关系数
R1:寒温带针叶林区域; R2:温带草原区域; R3:温带荒漠区域;R4:青藏高原高寒植被区域;R5:暖温带落叶阔叶林区域;R6:温带针叶、落叶阔叶混交林区域;R7:亚热带常绿阔叶林区域;R8:热带季风雨林、雨林区域。“研究区”在春季、夏季和秋季指除台湾省以外的整个中国植被覆盖区,冬季只涉及除台湾省以外的R7,R8地区;*表示通过P<0.05显著性检验,**表示通过P<0.01显著性检验
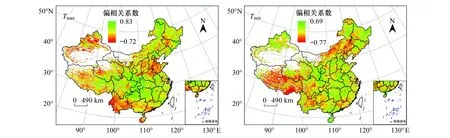
图6 春季NDVI与Tmax、Tmin偏相关系数空间分布图Fig.6 Spatial patterns of the partial correlation coefficients between spring NDVI and Tmax / Tmin
从夏季植被与昼夜气温的相关性空间分布(图7)可以看出,约60.05%的地区NDVI与Tmax呈现正相关,通过显著性检验的区域占研究区的9.98%,主要分布在大兴安岭北部、东北东部山地、西藏东北部、青海东部及我国南方地区。NDVI与Tmax呈现显著负相关的区域占研究区的比值仅为4.43%,主要零星分布于新疆西北部、青藏高原中部及东部、云南高原中部及东北平原地区。约50.56%的地区NDVI同Tmin呈现正相关,呈现显著正相关的地区占研究区的5.80%,主要分布在西北地区及东北地区。NDVI同Tmin呈现显著负相关的地区占研究区的4.70%,主要分布在西南地区、黄土高原南部及鲁中山地丘陵地区。由此可见,昼夜增温对植被产生积极影响的地区面积均高于产生消极影响的地区。与春季相比,夏季Tmax与NDVI呈现显著正相关的比值较低,其原因可能是由于夏季白天气温在大部分地区已接近植被生长的最适温度,导致该季节植被对白天增温的敏感程度较低[31]。相关结果表明,夏季白天温度的上升可能会降低水的可利用性,对植被的生长产生消极影响,这可能是造成部分地区NDVI与Tmax呈现为负相关关系的原因[18]。
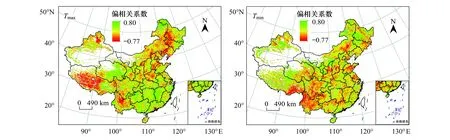
图7 夏季NDVI与Tmax、Tmin偏相关系数空间分布图Fig.7 Spatial patterns of the partial correlation coefficients between summer NDVI and Tmax / Tmin
分析秋季植被与昼夜气温的相关性空间分布(图8)后发现,约64.23%的地区NDVI与Tmax呈现正相关,呈现显著正相关的地区占研究区的11.51%,其主要位于新疆西部、云南高原、四川盆地及东北地区北部。NDVI与Tmax呈现显著负相关的地区仅占研究区的1.55%,其主要零星分布在我国东部沿海地带。 约63.30%的地区NDVI同Tmin呈现为正相关,呈现显著正相关的地区占研究区的8.21%,其主要位于新疆西部、华北平原、江南丘陵、贵州高原及大兴安岭南部地区。NDVI与Tmin呈现为显著负相关的地区占研究区的2.48%,主要零星分布在准噶尔盆地西侧、大兴安岭北部及云南高原地区。上述结果表明,秋季昼夜增温对大部分地区植被NDVI的提高产生积极影响。与春季相比,秋季植被对白天增温产生积极响应的地区面积较少,其原因可能因为秋季植被光合作用持续时间及植被生产力对温度的敏感程度低于春季[32]。

图8 秋季NDVI与Tmax、Tmin偏相关系数空间分布图Fig.8 Spatial patterns of the partial correlation coefficients between autumn NDVI and Tmax / Tmin
从冬季植被与昼夜气温的相关性空间分布(图9)来看,约84.18%的地区NDVI与Tmax呈现正相关,呈现显著正相关的地区占研究区的27.12%,其主要位于汉中盆地、云贵高原及江南丘陵地区。NDVI与Tmax呈现显著负相关的地区占研究区的1.26%,其主要零星分布于云南高原中部。约63.68%的地区NDVI与Tmin呈现正相关,呈现显著正相关的比值为8.23%,主要分布在云南高原、海南省及长江中下游平原地区。NDVI与Tmin呈现为显著负相关的地区仅占研究区的0.87%,主要零星分布在广西南部及广东北部。由此可见,冬季昼夜增温利于绝大部分地区植被的生长,其原因可能是由于温度作为该季节植被生长的主要限制因子,温度的升高可以进一步提升植物光合作用酶的活性。
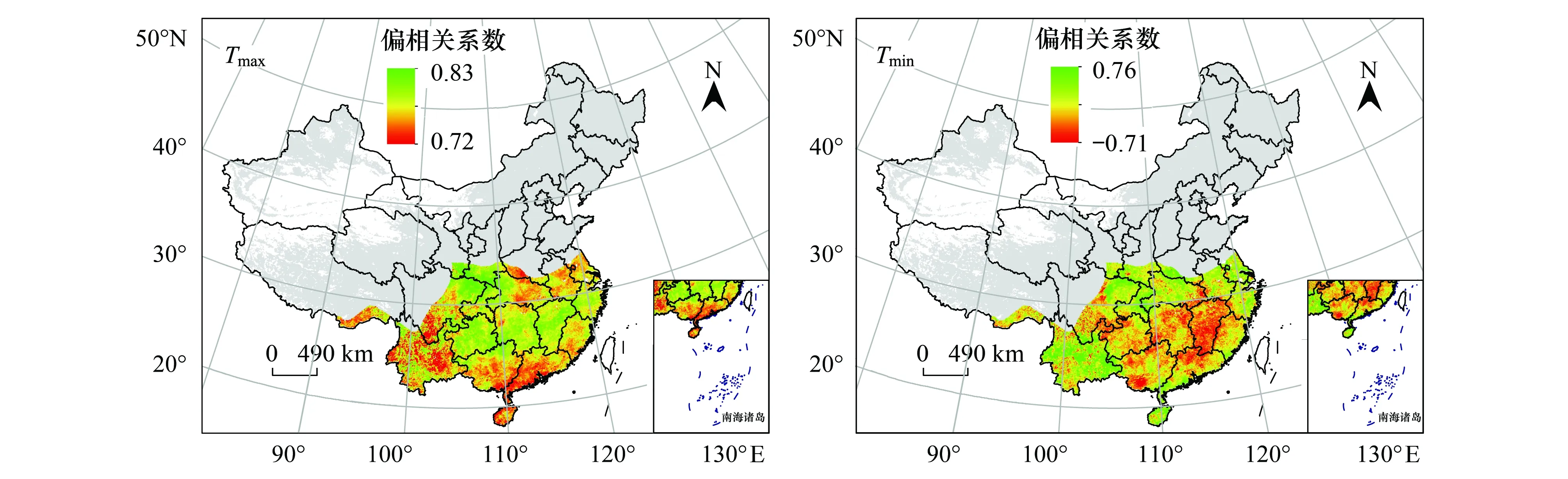
图9 冬季NDVI与Tmax、Tmin偏相关系数空间分布图Fig.9 Spatial patterns of the partial correlation coefficients between winter NDVI and Tmax / Tmin
3 结论与讨论
3.1 结论
本文利用中国1982—2015 年的气象及卫星遥感观测数据集,在季节尺度上分析了昼夜增温的变化趋势及其对植被影响。得出如下结论:
1)近34年来中国昼夜气温在各个季节均呈现为显著上升态势,但不同季节昼夜增温速率存在较大差异;昼夜增温在不同季节均表现出不对称变化特征,且不对称变化特征存在明显的季节性差异,其中春季和冬季白天增温速率快于夜间,秋季和夏季夜间增温速率快于白天;昼夜增温速率在空间格局上也存在明显差异。
2)从区域尺度来看,近34年来我国植被NDVI与Tmax在各个季节中均呈现为正相关关系,其中春季和冬季呈现为显著正相关关系,而各个季节中NDVI与Tmin的相关性均不显著,说明相对于夜间增温,白天增温对我国植被活动影响程度更大;从像元尺度来看,春季和冬季研究区植被NDVI与昼夜气温通过显著性检验的比例更高,并且更多地表现为正相关关系,这说明春季和冬季昼夜增温对我国植被活动的影响范围更广,且多利于植被NDVI的提升。
3)近34年来,我国各类型植被覆盖区在不同季节昼夜增温速率差异明显;不同类型植被对昼夜增温速率的不对称性产生了不同的响应,并且在各个季节上的响应程度存在差异。
3.2 讨论
本文通过遥感数据和气象数据分析了我国各季节昼夜气温的变化规律,并对我国植被对昼夜增温的季节性响应特征进行了探讨。研究发现,我国植被在不同季节对昼夜增温的响应存在明显差异。尽管大部分地区对昼夜温度的上升表现出积极的响应,但昼夜温度的上升对植被NDVI产生不利影响的地区在不同季节均有存在。在温度作为植被生长的主要限制因子的地区,Tmax的上升可能会通过提升光合作用酶的活性[33]、提高土壤氮的可利用性[34]以及延长植被生长周期[29],对植被NDVI的增加起到推动作用。然而,在受水分限制的干旱半干旱区,白天增温可能会通过提高植被的蒸腾作用[35]、加速土壤水分蒸发、降低土壤水含量,对植被生长造成不利影响[35-36]。尽管Tmin与植被NDVI之间通过显著性检验的地区较少,但类似于白天增温,Tmin同样可以通过以下两种方式对植被生产力产生不同的影响。一方面,夜间增温能够通过增加植被自养呼吸速率[37],降低植物成熟期胚乳细胞的体积[38]以及缩短植物灌浆期[39],对植被NDVI的提升产生消极影响。值得注意的是,夜间自养呼吸的增加也可能会通过补偿作用,刺激植被次日光合能力的提高[14,40]。另一方面,夜间增温能够通过降低霜冻灾害发生频率[41],提升植物对干旱的抵抗能力[42]以及调节植物叶片中碳水化合物含量[14],对植被生产力的提高带来积极影响。
植被的动态变化受到地理因子(地形、土壤条件、地表粗糙度等)、气候因子(气温、降水、云层覆盖度、太阳辐射量等)以及人为因素的共同干扰。不同区域因其地理条件、气候条件以及人类干扰程度的差异,决定植被动态变化的关键因子存在差异。由于数据的可获得性,本文未能综合考虑所有影响植被动态变化的控制因子;此外,由于植被对气候因子的响应往往存在一定程度的滞后性,且在植被生长的不同阶段对气候因子的滞后时间同样存在差异[43-44],这些因素势必对研究结果的准确性造成一定的影响。因此,在以后的研究中需要综合各种因素,辅以控制实验或者数学模型等手段,以厘清季节性昼夜增温对不同植被的影响机理[24]。
猜你喜欢
河北地质(2022年2期)2022-08-22
成都信息工程大学学报(2022年3期)2022-07-21
今日农业(2021年2期)2021-03-19
科学大众(2020年17期)2020-10-27
快乐作文(1.2年级)(2019年3期)2019-09-10
英语文摘(2019年5期)2019-07-13
幼儿画刊(2018年10期)2018-10-27
现代园艺(2017年23期)2018-01-18
小雪花·成长指南(2016年2期)2016-03-16
应用海洋学学报(2015年2期)2015-11-22
