
桃蛀果蛾对不同品种苹果的适生性研究
2018-07-10 11:36张志伟门丽娜吴志毅孟廷玲马瑞燕
山西农业大学学报(自然科学版) 2018年7期
张志伟,门丽娜,吴志毅,孟廷玲,马瑞燕
(1.山西农业大学 林学院 山西 太谷 030801;2.山西农业大学 农学院,山西 太谷 030801;3.浙江出入境检验检疫局,浙江 杭州 310016)
桃蛀果蛾CarposinasasakiiMatsumura,隶属于鳞翅目(Lepidoptera)蛀果蛾科(Carposinidae),是我国北方果树产业危害较为普遍的蛀果类害虫,严重为害苹果、枣、梨、山楂等10多种果树的果实[1, 2]。早在20世纪50年代我国学者就曾对苹果、梨、山楂以及枣等果树上的桃蛀果蛾危害状况进行了研究。在东北地区,桃蛀果蛾严重为害苹果[3],使用广谱杀虫剂后蛀果率大大降低[4],但广谱杀虫剂的过度使用却给果品质量安全和生态环境带来了挑战。近年来,随着人均收入水平的增加和消费结构的转变,人们对食品安全重视程度不断提高,果品供给主体面临日益增长的食品质量安全需求[5],化学杀虫剂施用量呈下降趋势,桃蛀果蛾的无公害防控备受果农青睐。
由于果树产业迅速发展,果树栽培、管理模式明显改变,果树蛀果类害虫的危害大幅回升[6]。研究昆虫对寄主植物不同品种的选择性和适生性,可以为昆虫的种群动态监测和防控、寄主品种选育提供重要理论依据[7~9]。由于在不同寄主植物间[10~13]、在同种寄主植物的不同品种间[14]、甚至不同的采果期[15],桃蛀果蛾的生长发育和繁殖均存在显著差异,导致其在果园的发生情况极其复杂,增加了种群监测和防控难度。在苹果园中,桃蛀果蛾是重要的果实害虫,为保证果品质量,需要对不同品种苹果园的桃蛀果蛾种群进行管理;而苹果经济价值高、品种地域性强[16],研究桃蛀果蛾对不同苹果品种的适生性,可以为桃蛀果蛾种群监测和苹果抗虫品种选育提供参考资料,具有重要的科学意义。
目前,从自然条件下获得大量发育整齐一致的虫源难度很大,很难满足科学实验研究,导致桃蛀果蛾对不同苹果品种选择性和适生性、生物生态学特性和人工饲养技术等研究仍不充分,制约了其防治工作。通过人工饲养获得大量整齐一致的试虫成为开展桃蛀果蛾相关科学研究的重要保证。用寄主饲养桃蛀果蛾取得了可喜的进展[17~19],能够为各自实验室的部分研究提供试虫。研究实验条件下桃蛀果蛾对不同品种苹果的适生性,组建实验种群生命表,可以为深入系统地研究桃蛀果蛾的生物生态学特性及人工饲养提供科学依据。
1 材料与方法
1.1 试验材料
1.1.1供试虫源
在2011、2012年7-8月,把采自山西省农业科学院果树研究所疏于管理的苹果园和枣园的虫果带回实验室,待老熟幼虫自然脱果后,从中收集桃蛀果蛾;因种群个体数量不足,以沈阳农业大学植物保护学院和中国农业科学院植物保护研究所馈赠桃蛀果蛾作为补充。所有桃蛀果蛾作为一个大种群,在实验室内连续传代饲养10个生命周期后,用于本试验研究。
1.1.2饲养条件
在微电脑光照培养箱内饲养桃蛀果蛾。饲养条件为T=(25.0±0.5) ℃,RH=75%±5%,光照时长15 h/24 h,光照强度4 800 lx,以市售不同品种苹果的果实数量性状[20]、货架期、价格,和山西中南部、陕西等黄土高原区苹果主栽品种及近年来种植面积不断扩大的加工品种为综合依据[21,22],选择红富士、华冠、金冠、澳洲青苹4个品种苹果饲喂幼虫。
1.1.3试验主要仪器、药品及耗材
微电脑光照培养箱(SPX-250B-G型,上海博讯)、体视镜(Olympus SZX7,日本Olympus);75%乙醇、琼脂;指形管(d=1 cm,h=10 cm)、养虫盒(d=10 cm,h=5 cm)、成虫交尾产卵盒(d=15 cm,h=20 cm)、镊子、剪刀、滤纸;苹果(果实直径60~70 mm)。
1.2 试验方法
1.2.1幼虫的饲养与观察
自产卵日起,每天收集卵卡,剪成带卵的小卡片,放入经75%乙醇消毒的养虫盒培养。待卵接近孵化时,用镊子小心地把经过培养的小卵卡用琼脂粘贴于新鲜、成熟、洗净的市售红富士、华冠、金冠、澳洲青苹的萼洼处,每枚苹果接卵20粒,用喷壶喷雾保湿,每个品种苹果10枚重复。每枚苹果均单果放入养虫盒、置于光照培养箱中培养。每隔12 h观察一次卵孵化状况,记录孵化数量,直至不再孵化。在体视镜下观察并统计每枚果实上的蛀孔数、未孵化卵数、死亡幼虫数,统计卵的孵化率和幼虫的蛀果率。饲养期间,每日晨、午、昏各检查一次是否有幼虫脱果,称量脱果的幼虫体重、记录幼虫发育历期。若果实连续5 d不再有幼虫脱出,则解剖、查看其中是否还有幼虫,统计幼虫存活率及脱果率。
1.2.2成虫的饲养与观察
当第一头老熟幼虫自然脱果后,每隔6 h查看一次幼虫脱果情况,将同日脱果的末龄幼虫置于同一个底部铺有湿沙的养虫盒中,记录结茧时间。一旦成虫开始羽化,根据桃蛀果蛾的羽化习性[23],当日16:00~21:00,每60 min观察1次存活情况,翌日8:00再次检查成虫是否死亡;记录羽化时间以及死亡情况。选取雌、雄成虫1∶1配对,用指形管小心移入成虫交尾产卵盒,底部铺滤纸供其产卵。在相同饲养条件下,统计产卵时间、每日落卵量、单雌产卵量,直至所有成虫全部死亡,并记录死亡时间。
1.3 数据统计
生命表的组建按照张孝羲[24]的方法,各参数计算公式如下:
净增殖率R0=∑(lxmx)
平均世代周期T=(∑xlxmx)/R0
内禀增长率rm=(lnR0)/T
种群加倍时间Dt=(ln2)/rm
周限增长率λ=erm
式中:x表示以天(d)为单位的时间期限;lx表示任一个雌性个体在x期间的存活率;mx表示在x期间平均每个雌虫繁殖的雌性后代数。
利用Microsoft Excel 2013软件记录、整理所得数据,SPSS 19.0软件进行单因素方差分析,百分数和比率数据经过反正弦平方根转换后再进行统计分析,Fisher’s LSD法多重比较检验。
2 结果与分析
2.1 桃蛀果蛾在不同品种苹果上的发育历期
桃蛀果蛾在不同品种苹果上的发育历期明显不同(表1)。卵历期在所选的4个苹果品种之间无显著差异(4.96~5.09 d,P>0.05)。桃蛀果蛾在金冠苹果上的幼虫历期(17.81 d)显著短于其他品种(P<0.05),发育最快;在华冠上桃蛀果蛾的幼虫历期(20.32 d)最长,但与红富士上的幼虫历期差异不显著(P>0.05)。不同品种苹果上桃蛀果蛾的结茧前期(1.03~1.18 d)、蛹期(9.57~9.78 d)和产卵前期(1.77~2.03 d)均无显著差异(P>0.05)。桃蛀果蛾在华冠苹果上的世代历期(39.45 d)极显著长于其他品种(P<0.01),在红富士、金冠和澳洲青苹上世代历期无显著差异(36.28~37.59 d,P>0.05)。
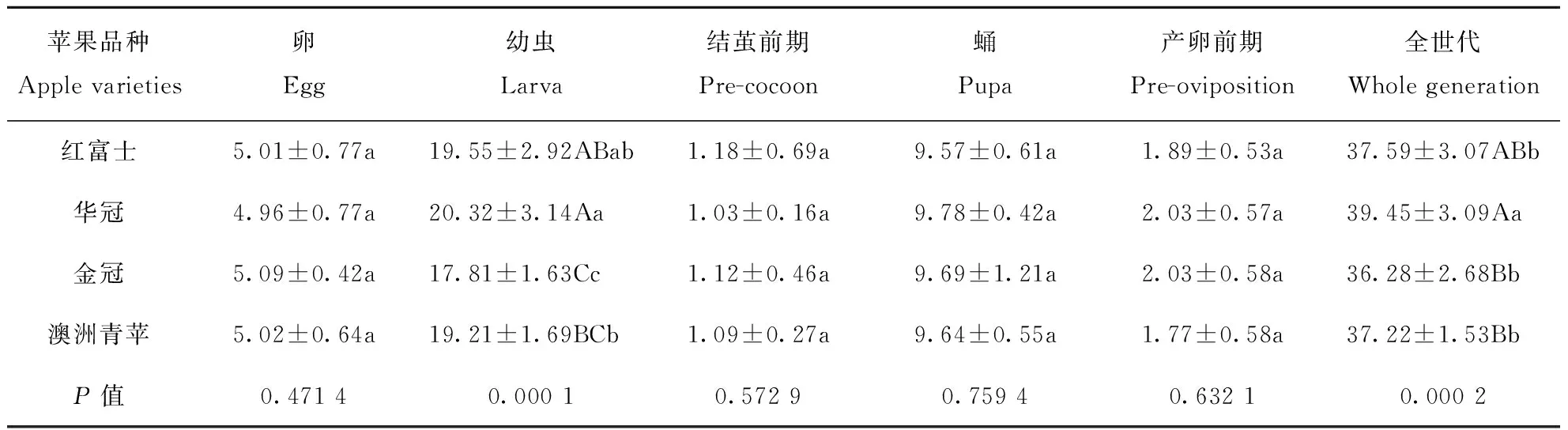
表1 桃蛀果蛾在不同品种苹果上的发育历期/dTable 1 Developmental duration of C. sasakii on different apple varieties
注:表中数据为(平均数±标准差);同一列数据后的不同大、小写字母表示有极显著差异(P<0.01,LSD)和显著差异(P<0.05,LSD);下表同。
Note:Remarks Mean±SD followed by different capital and small letters within the same column mean that extremely significantly different (P<0.01, LSD), significantly different (P<0.05, LSD), respectively. Same for the following tables.
2.2 桃蛀果蛾在不同品种苹果上的存活率
桃蛀果蛾的发育,包括孵化,幼虫蛀果、果内生长发育和老熟幼虫脱果,结茧及羽化等系列过程。桃蛀果蛾的这些关键生命节点在不同品种苹果上存有差异(表2)。桃蛀果蛾在红富士上的孵化率(75.50%)明显高于其他品种,但无显著差异(P>0.05)。在金冠和红富士上的蛀果率分别为91.25%和88.36%,极显著高于华冠(P<0.01);在澳洲青苹上的蛀果率仅为26.20%,极显著低于其他品种(P<0.01)。桃蛀果蛾在4个不同品种苹果上的脱果率无显著差异(P>0.05)。在澳洲青苹上的结茧率显著低于红富士和金冠(P<0.05),与华冠无显著差异(P>0.05)。羽化率在红富士上最高(80.42%),但和其他品种上的羽化率无显著差异(P>0.05)。幼期存活率以金冠上的为最高(42.50%),红富士次之(36.00%),澳洲青苹最低(13.00%)。
昆虫不同年龄-龄期的存活率高低及存活数量多寡决定其种群的兴衰,也影响昆虫对寄主植物的危害程度[7]。桃蛀果蛾在金冠苹果上的蛀果率和幼期存活率最高,故危害严重,即桃蛀果蛾对金冠的适生性最好,金冠对其抗性最差;而在澳洲青苹上蛀果率和幼期存活率最低,危害较轻,即桃蛀果蛾对澳洲青苹的适生性较差,澳洲青苹对其有较强的抗性。

表2 桃蛀果蛾在不同品种苹果上的存活率/%Table 2 Survival rate of C. sasakii on different apple varieties
2.3 不同品种苹果对桃蛀果蛾生长和繁殖的影响
桃蛀果蛾的幼虫重、成虫交配率、单雌产卵量和次代孵化率都是评价其生长和繁殖的常用指标。本实验条件下,不同品种苹果上的桃蛀果蛾在这些指标上也存有差异(表3)。桃蛀果蛾在澳洲青苹上的幼虫重(27.50 mg)极显著高于其他品种(P<0.01),其他苹果品种之间,桃蛀果蛾幼虫重差异不显著(P>0.05)。不同品种苹果作为寄主,桃蛀果蛾成虫交配率由高到低依次为金冠>红富士>澳洲青苹>华冠,金冠为寄主时,桃蛀果蛾成虫交配率最高,为79.80%。澳洲青苹为寄主时,桃蛀果蛾单雌产卵量(90.63粒)最大,但与其它品种无显著差异(P>0.05);澳洲青苹为寄主时,次代孵化率最高(97.20%),华冠为寄主时次之(95.60%),红富士为寄主时次代孵化率最低(91.50%)。
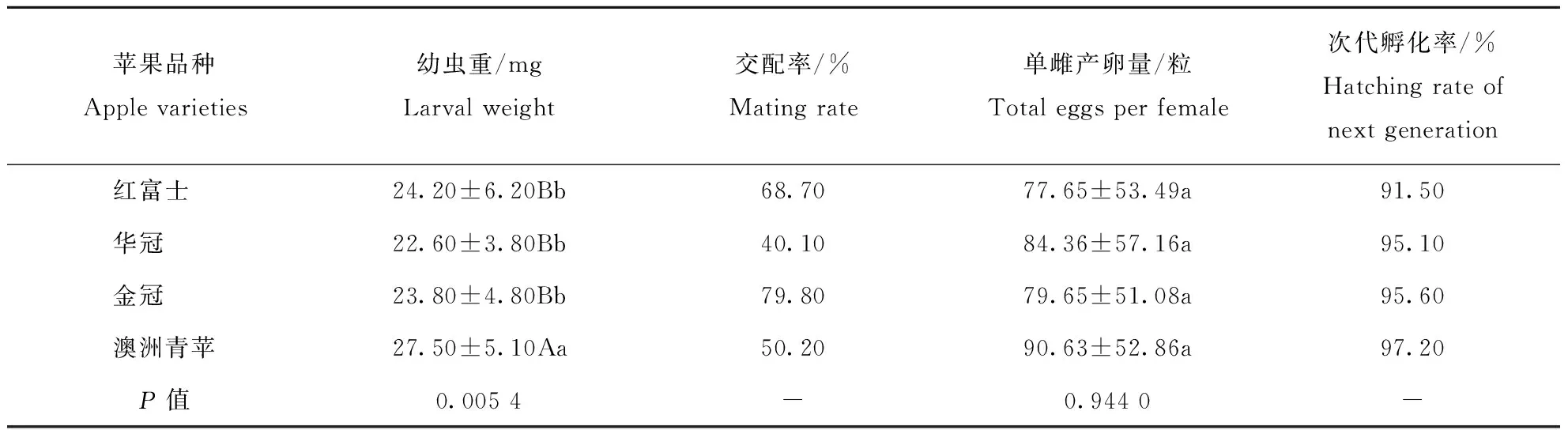
表3 桃蛀果蛾在不同品种苹果上的幼虫重和繁殖能力Table 3 Larval weight and fecundity of C. sasakii on different apple varieties
2.4 桃蛀果蛾在不同品种苹果上的实验种群趋势
根据相同试验条件、4个不同品种苹果上的桃蛀果蛾各发育期的存活率及成虫生殖力资料,以实际接卵量作为起始卵数,各发育阶段的死亡虫数和性比均用实际观察值,分别组建了不同品种苹果上的桃蛀果蛾实验种群特定年龄生命表。桃蛀果蛾在所选的4个苹果品种上的种群趋势指数>1(表4),即下一代种群数量为增长趋势。金冠上种群趋势指数I最大,为12.35,即:经过一个世代,种群数量达到原来的12.35倍;红富士上的种群趋势指数次之,为10.87;澳洲青苹上的种群趋势指数最小,仅为4.98。
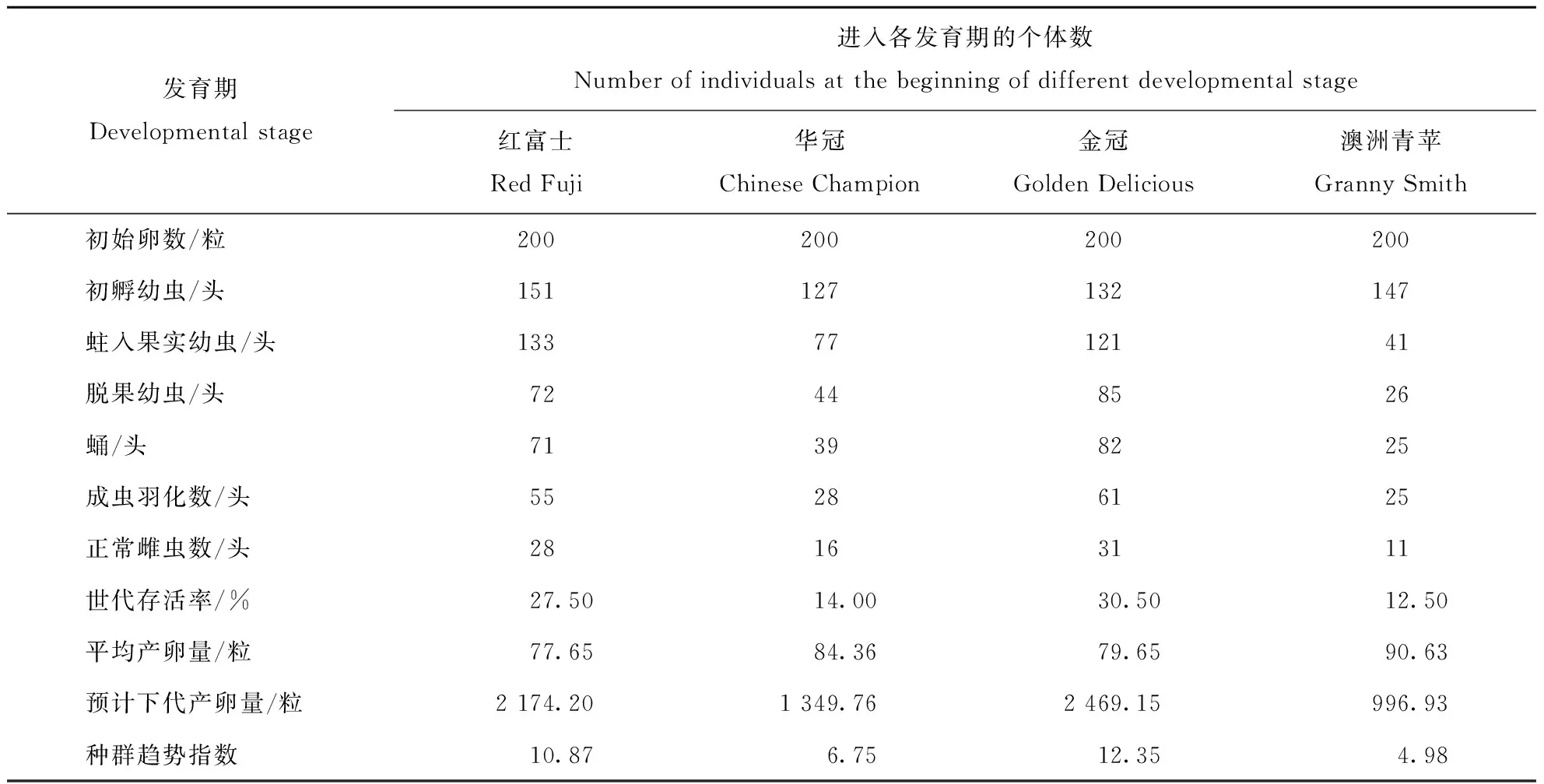
表4 桃蛀果蛾在不同品种苹果上的实验种群生命表Table 4 Life table of the laboratory population of C. sasakii on different apple varieties
根据桃蛀果蛾在所选4个品种苹果上的实验种群生殖力数据,计算其在不同品种苹果上的种群净增殖率(R0)、内禀增长率(rm)、周限增长率(λ)、平均世代周期(T)和种群加倍时间(Dt)。净增殖率(R0)以金冠为寄主时最高(7.56),红富士次之,澳洲青苹最小(4.53);内禀增长率(rm)在不同苹果品种之间的变化规律和净增殖率(R0)相似,也是以金冠为寄主时最高(0.0581),红富士次之,澳洲青苹最小(0.0417);不同品种苹果做寄主,周限增长率(λ)>1,即桃蛀果蛾实验种群呈几何级数增长;平均世代周期(T)以金冠为寄主时最短(34.86 d),红富士次之,华冠最长(37.61 d);种群加倍时间(Dt)和净增殖率(R0)、内禀增长率(rm)的变化规律相反,以金冠为寄主时最短(11.94 d),红富士苹果次之,澳洲青苹最长(16.63 d)(表5)。
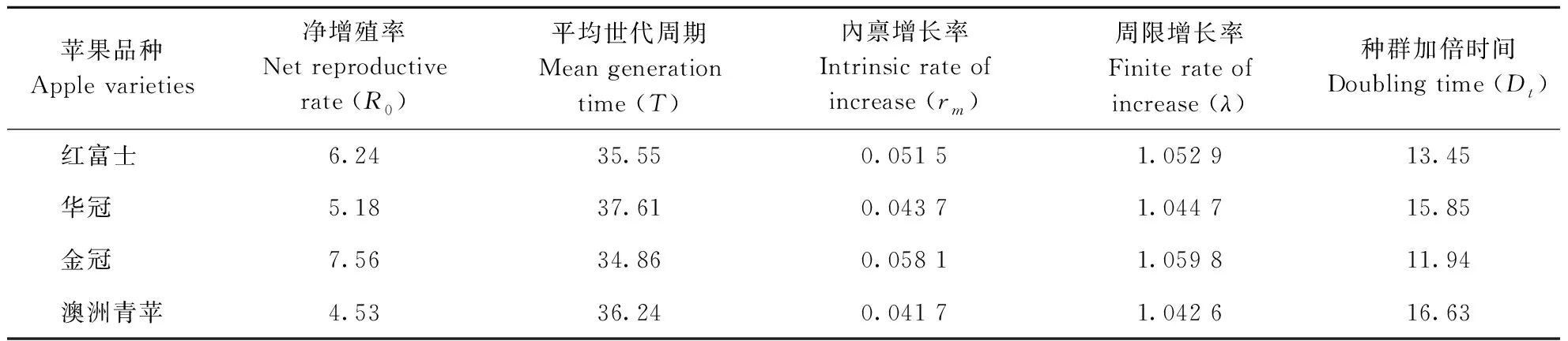
表5 桃蛀果蛾在不同品种苹果上的实验种群繁殖特征参数Table 5 Laboratory population parameters of C. sasakii on different apple varieties
3 讨论与结论
昆虫与其寄主植物间的关系十分复杂,寄主植物表面组织结构、寄主植物体内化学物质以及次生代谢物等均能影响昆虫的个体生长行为和繁殖行为,以及与之相关的一系列生命活动[7~9]。桃蛀果蛾为害多种果树的果实,但其对不同果树果实的嗜好程度存有差异,不同寄主植物通过影响害虫生长发育和繁殖进而对其生活史造成显著影响[12]。因此,选育抗桃蛀果蛾或忍耐桃蛀果蛾危害的果树品种是其治理的重要环节。本研究在相同试验条件下,详细比较了桃蛀果蛾在4个不同品种苹果上的生长发育和繁殖能力,发现桃蛀果蛾对不同苹果品种的适生性有明显差别,这些差别主要表现在桃蛀果蛾在金冠上幼虫发育历期最短(17.81 d)、蛀果率最高(91.25%)、幼期存活率最高(42.50%),实验种群的净增殖率(R0=7.56)和内禀增长率(rm=0.0581)最高,平均世代周期(T=34.86 d)和种群加倍时间(Dt=11.94 d)最短、发育最快,表明桃蛀果蛾对金冠苹果的适生性最好;这在国光、新红星和金冠等苹果品种对桃蛀果蛾生长发育和繁殖的影响研究中也得到了证实[14]。
本研究发现:桃蛀果蛾在不同品种苹果上实验种群的周限增长率(λ)均大于1,即桃蛀果蛾的实验种群均以几何级数增长;李定旭等[12]研究不同物种寄主植物上的桃蛀果蛾实验种群,也证实存有这个规律。
桃蛀果蛾在金冠和红富士上的存活率明显高于华冠和澳洲青苹,而世代历期相对较短、发育快,说明桃蛀果蛾对金冠和红富士的适生性较好,生长发育和存活指标均优于华冠和澳洲青苹,同时也反映出金冠和红富士对桃蛀果蛾的抗性较差。虽然桃蛀果蛾的幼虫体重在澳洲青苹上显著高于其他品种,但是存活率却最低(12.0%),蛀果率也最低(26.20%),说明桃蛀果蛾对澳洲青苹的适生性较差,即澳洲青苹对桃蛀果蛾有较强的抗性。澳洲青苹上桃蛀果蛾蛀果率低、幼虫体重高,可能由于澳洲青苹的果皮厚度大[25],导致每枚苹果实际蛀入的幼虫数量较少,幼虫个体之间竞争小[26],可能使幼虫生长发育状况优于蛀入果实数量多的处理。
桃蛀果蛾在本研究所选择的4个不同品种苹果上幼虫发育历期在17~20 d之间,短于张乃鑫等[27]的研究结果,主要原因可能是试验条件和寄主的成熟度有所不同:张乃鑫等[27]用未采摘的苹果接卵、在果园试验得出结果,果园温度不恒定、昼夜温差较大、苹果的成熟度和非离体状态都可能会影响桃蛀果蛾幼虫的发育历期[15, 28]。本研究采用不同品种的成熟苹果采摘后饲养桃蛀果蛾,采摘后的成熟果实中桃蛀果蛾幼虫的发育历期变短[15],且存活率高,也可能是我们试验结果和张乃鑫等[27]有差异的原因之一。本研究桃蛀果蛾的幼虫发育历期长于Toyoshima等[29]和张怀江等[14]所报道的结果,可能是所应用的苹果成熟度不同,且不同产地、不同生态区的富士苹果之间存有差别[30],以及试验条件不同,导致研究结果有差别,需要进一步研究。有研究表明,寄主物种不同会导致桃蛀果蛾幼虫发育历期有显著差异[12, 13],我们的研究进一步发现:同一种寄主的不同品种、同一品种寄主的不同成熟度也会影响桃蛀果蛾幼虫的发育历期。桃蛀果蛾为害后,寄主的蛋白质含量和应激保护性酶活性发生变化,不同品种苹果的保护酶活性差异显著[31],可能是导致桃蛀果蛾在不同品种苹果间个体生长发育产生差异的重要原因,进而导致桃蛀果蛾的种群生物学指标明显不同。
评价桃蛀果蛾对不同苹果品种的适生性,可以用幼虫发育历期、蛀果率、幼期存活率,实验种群的净增殖率、内禀增长率、平均世代周期和种群加倍时间作为参考指标,这些指标也可以用来评价人工饲养桃蛀果蛾时选用的寄主是否适宜。用不同品种苹果饲养桃蛀果蛾,孵化率、脱果率、结茧率和羽化率都较高,而蛀果率和幼期存活率差别明显,说明桃蛀果蛾幼期的发育状况是其整个世代的关键环节。本研究发现以金冠苹果饲养的桃蛀果蛾幼期发育状况最好,李定旭等[12]、Lei等[13]、张怀江等[14]的研究也发现金冠是饲养桃蛀果蛾的适宜寄主。澳洲青苹作为寄主虽然可以获得生长发育状况很好的桃蛀果蛾幼虫,但是蛀果率却显著低于其它品种,因此要获得相同数量的后代就要增加饲养用苹果的使用量,增加人工饲养的材料成本。本研究设置的试验条件,桃蛀果蛾卵期和蛹期明显短于李定旭等[12, 32],Lei等[13]和张怀江等[14]的结果,而幼虫期(包括结茧前期)明显较长,可能是所选苹果品种不同和试验条件有差异所致;但世代历期大致相同,说明试验条件不同虽然会导致桃蛀果蛾不同虫态的发育历期有差异,但对整个世代历期和幼期存活率基本没有影响。由此可见,人工饲养桃蛀果蛾,合理设置和调整饲养条件,可以在一定程度上调控桃蛀果蛾不同虫态的发育历期。
猜你喜欢
小学生学习指导·高年级(2022年2期)2022-02-16
湖北农业科学(2021年22期)2021-12-13
湖北农业科学(2020年17期)2020-11-09
农家科技下旬刊(2020年5期)2020-06-19
民主与法制(2018年35期)2018-09-19
当代教育(2018年2期)2018-08-20
中国果业信息(2018年11期)2018-01-17
幸福·悦读(2016年8期)2016-10-21
中国茶叶(2016年7期)2016-01-18
中国当代医药(2015年32期)2015-03-01
