
广东省樟树立木生长规律和生长模型研究
2018-07-07 03:47林丽平徐期瑚雷渊才
中南林业科技大学学报 2018年6期
林丽平,徐期瑚,罗 勇,雷渊才
(1.广东省林业调查规划院,广东 广州 510520;2.中国林业科学研究院 资源信息研究所,北京 100091)
樟树Cinnamomum camphora为樟科樟属常绿乔木,是亚热带常绿阔叶林的代表树种,也是亚热带地区重要的用材和特种经济树种。樟树生长速度中等,成型快,造林成活率高,既可作行道树、庭荫树,又是营建风景林、防护林的优良树种,适于在城镇绿化和乡村绿化中大力推广。樟树是广东优良的珍贵阔叶树种,也是绿化广东大行动开展碳汇造林工程的主要树种之一。目前对樟树良种选育[1-2]、扦插组培快繁[3-4]、逆境生理响应[5]、人工林生物量动态[6]、光合特性[7]及其对CO2浓度和温度升高、酸雨的响应[8-10]、病虫害防治[11]等方面已有许多研究,但对樟树的生长规律和生长模型的研究还较为缺乏。而掌握樟树立木的生长特征及规律,并建立樟树胸径、树高和材积的生长模型, 对科学经营、深入推广和培育樟树优质木材具有重要意义。本文拟针对广东省樟树资源现状,科学评估樟树不同年际的生长过程及其动态变化规律,为预估广东省樟树胸径、树髙、材积的生长,培育樟树大径材以及可持续经营管理提供技术支撑和理论依据。
1 研究区概况
研究区位于广东省全省境内,地处南海北部,有珠江三角洲平原与潮汕平原,其余为山地、丘陵, 地 理 坐 标 为 109°45′~ 117°20′E,20°09′~25°31′N,面积 17.97×104km2,该区属于东亚季风区,从北向南分别为中亚热带、南亚热带和热带气候,是全国光、热和水资源最丰富的地区之一,年平均气温21.8 ℃,最冷月1月平均气温为13.3 ℃,最热月7月平均气温为28.5 ℃,年平均降水量1 789.3 mm,年平均日照时数自北向南逐渐增加,由不足1 500 h增加到2 300 h以上,年太阳总辐射量在4 200~5 400 MJ/m2之间。广东省地带性森林植被的主要类型为中亚热带常绿阔叶林、南亚热带常绿阔叶林和少量热带季雨林。地带性土壤主要类型为赤红壤和红壤。
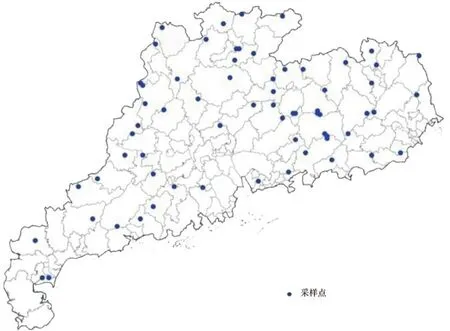
图1 广东省樟树分布情况及样地布设Fig.1 Study region and sample plots set in Guangdong province
2 研究方法
2.1 试验材料
在广东省内以连清样地中的樟树分布情况为基础,根据不同立地条件,采用单株伐倒法按10个径阶区间选取90株样木进行伐倒木测定。测定因子包括所处的立地条件(海拔、坡位、坡向、坡度)以及树种的起源、胸径、树高、枝下高、冠幅和坐标等。在此基础上,按径阶大小选取其中40株樟树标准木采用中央断面积区分法进行树干解析[12],计算平均生长量和连年生长量。
2.2 数据的采集与处理
解析木圆盘按一年一个龄级进行划分,先对圆盘的工作面进行抛光处理,对经抛光和标记南北直径线的圆盘40株樟树样品,用WinDENDRO测量每个圆盘东西南北四个方向的年轮宽度,数据整理之后使用Forstat2.2进行树干解析。各龄阶去皮直径取圆盘截面东西和南北向的均值,树龄与各圆盘的年轮数之差,即为林木生长到该断面高所需要的年数,各龄阶的材积采用梯形法进行计算。
(1)标准木单株材积应用平均断面积法(即梯形法)求得,材积计算公式为:
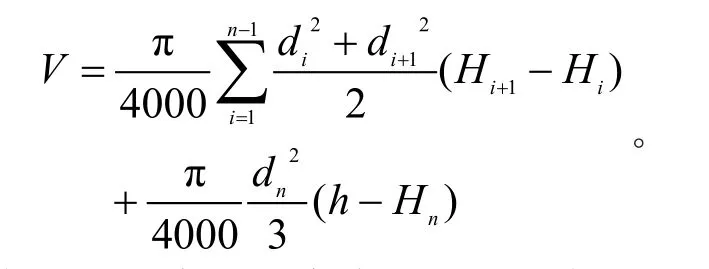
式中:Hi为离地面高度;di为带皮或和去皮直径;i=1,2,3,…,n;h为树木总高度。
(2)平均生长量计算公式:MAI=Yt-1/t。
式中:MAI为平均生长量,Yt是第t年直径、树高或材积的生长量。
(3)连年生长量计算公式:CAI=Yt-Yt-1。式中:CAI为连年生长量,Yt是第t年直径、树高或材积的生长量,Yt-1是第t-1年直径、树高或材积的生长量。
从90株伐倒木中随机抽取60株样木进行樟树林分胸径、树高和材积生长建模,其余30株样木数据作为验证数据。各样木调查情况如表1所示。
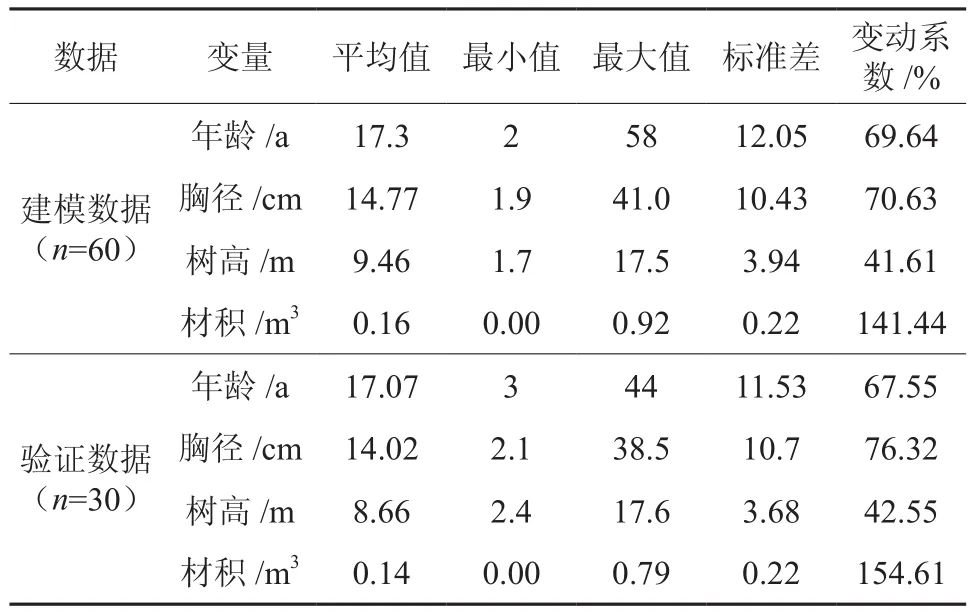
表1 广东省樟树样木调查因子统计量Table 1 Survey factor statistics of Cinnamomum camphora in Guangdong province
2.3 模型拟合与检验
模型参数估计使用了R软件的nlme过程对随机取抽的60株樟树伐倒木进行拟合,运用该函数算法来解决非线性最小二乘问题,具体公式见表2。
为了检验生长方程的拟合效果,采用赤池信息准则(AIC)、拟合优度(R2)、均方根误差(RMSE)、平均偏差(ME)、平均绝对偏差(MAE)对模型的拟合效果进行评价,模型评价公式如下:

表2 樟树7种生长方程模式[13-16]†Table 2 Seven kinds of growth equations of Cinnamomum camphora
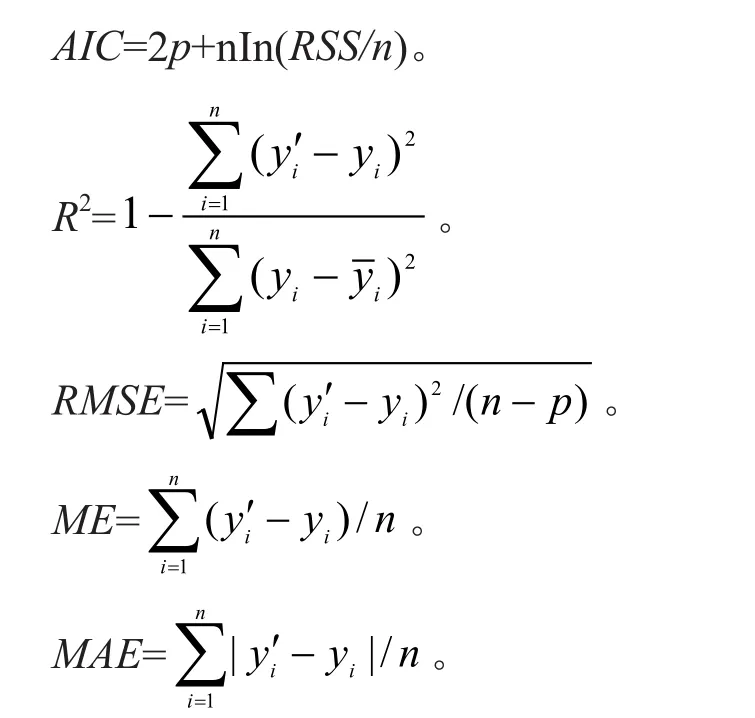
式中:yi是样本某因子的实测值;是yi的无偏估计值;是样本实测值的平均值;n是样本数量;p是参数的个数。通常认为AIC越小,RMSE、ME、MAE越接近0,R2的值越接近1,模型拟合效果越好。
3 结果与分析
3.1 生长过程分析
3.1.1 直径生长过程分析
樟树直径生长量见图2。从图2可以看出,(1)樟树生长初期比较缓慢,前3年直径总生长量仅有0.70 cm,前8年直径总生长量小于5 cm,到44 a去皮直径总生长量可达30.86 cm;林木个体的生长差异,前4年差异不大,5~20 a差异开始显现,20 a后随着年龄的增加而差异增大。(2)解析木在整个生长期内的生长过程符合树木生长的一般规律,即直径平均生长量的最大值比连年生长量的最大值出现的晚[17],在树龄11 a时,连年生长量达到高峰值,为1.03 cm,在树龄16 a时,直径平均生长量达到高峰值为0.74 cm,每10 a平均生长量为6.14 cm。(3)樟树在3~29 a平均生长量小于连年生长量,在树龄29 a时,平均生长量与连年生长量相等,在树龄29 a后平均生长量大于连年生长量。13~20 a樟树生长较快,直径平均生长量和连年生长量都在0.70 cm以上,表明樟树生长的速生期可能在13~20 a期间,此后平均生长量稍有下降后再呈平缓趋势。年轮波动较大的在进入速生期前,速生期和缓慢生长期波动幅度较小。

图2 樟树直径生长量Fig.2 Growth of diameter of Cinnamomum camphora
3.1.2 树高生长过程分析
樟树树高生长量见图3。从图3可以看出:(1)樟树树高初期生长比较缓慢,4~20 a树高生长速度较快且个体差异不明显,20 a后生长速度变慢,40 a后生长速度趋于稳定,樟树喜光,稍耐荫,适应性强,在生长初期有足够的光热资源,从而促进了树高的生长,树高一般能达到14~15 m。(2)樟树前18年树高以较大的速度增长,平均生长量均大于0.50 m。18 a后生长速度变慢,平均生长量为0.35 m。除苗期外,树高连年生长量和平均生长量随着年龄的增加总体上呈震荡式下降趋势,连年生长量波动范围比平均生长量要大,并在第5年时连年生长量达到峰值,为0.64 m。整个生长期连年生长量与平均生长量多次相交,呈多峰状。
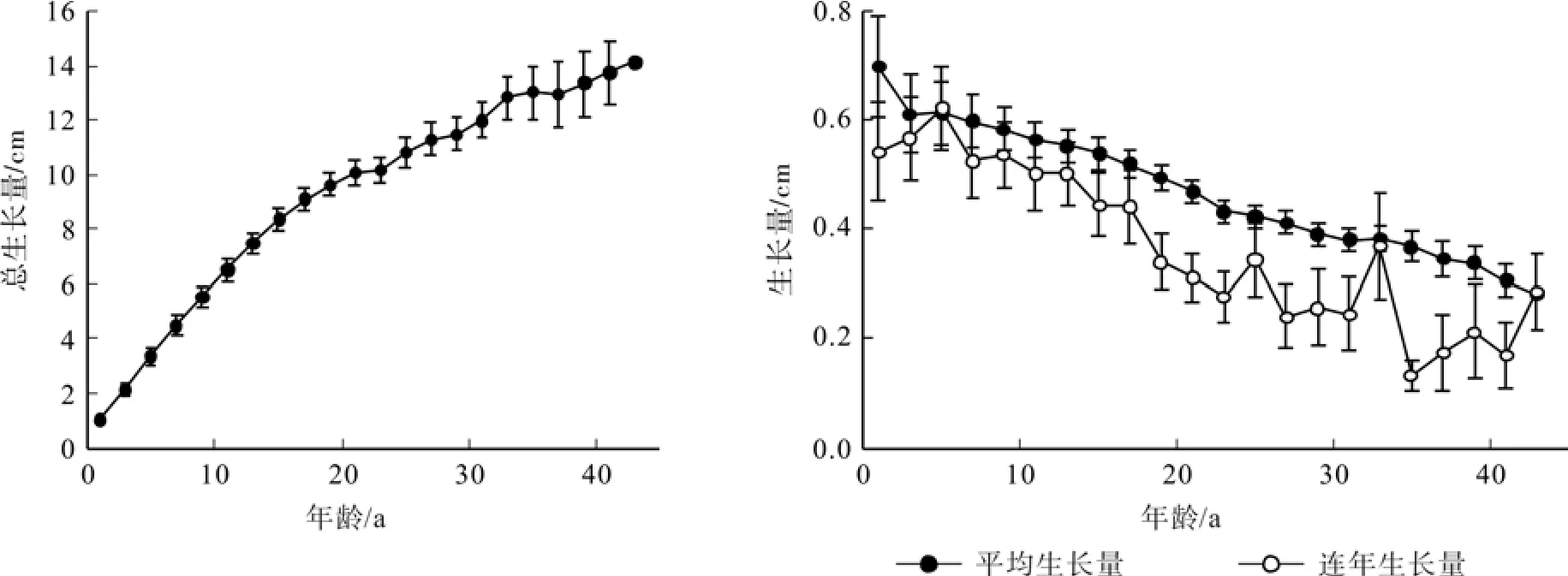
图3 樟树树高生长量Fig.3 Height growth of Cinnamomum camphora
3.1.3 材积生长过程分析
樟树材积生长量见图4。从图4可以看出,(1)前10年樟树单株材积的总生长量极小,且个体间差异不明显,10 a后随着年龄的增加生长变快,到44 a材积总生长量可达0.43 m3,且个体间差异逐步增大。(2)樟树材积平均生长量和连年生长量均随年龄的增加而增加,平均生长量和连年生长量高峰期出现的时间比较接近,平均生长量的高峰期在39 a,为0.009 m3,连年生长量的高峰期在36 a,为0.019 m3,40 a后平均生长量和连年生长量略有下降,这与直径和树高的生长高峰期出现的规律不一致,主要原因是由于随着直径的增加,材积呈几何级数迅速增加。第5年时,樟树的直径连年生长量为0.75 cm,材积连年生长量为0.001 m3;第15年时,直径连年生长量为0.89 cm,材积连年生长量为0.009 m3;第25年时,直径连年生长量为0.74 cm,材积连年生长量为0.013 m3;第35年时,直径连年生长量为0.60 cm,材积连年生长量为0.018 m3。随着年龄增大,直径增加,材积增长的速度也增加。材积的平均生长量和连年生长量没有相交点,说明樟树到44 a时还未达到成熟年龄,亦不能确定其主伐年龄。

图4 樟树立木材积生长量Fig.4 Growth of volume of Cinnamomum camphora
3.2 胸径、树高、单株材积的生长方程模拟及模型评价
3.2.1 胸径、树高、单株材积的生长方程模拟及评价
从表3分析可以看出,不论是对樟树的胸径、树高还是材积,Weibull生长方程都不收敛,对单株进行建模,也只有个别单株样木收敛。Richards和Korf生长方程对材积不收敛。

表3 樟树胸径、树高及材积的生长模型拟合Table 3 Growth model fitting of DBH, height and volume of Cinnamomum camphora
从总体上看,Gompertz、Mitscheerlich和Schumacher方程的拟合效果较好,从拟合优度检验来看,拟合决定系数R2差异不大,同一指标的R2相差0.01~0.03,立木胸径、树高和材积的最优拟合模型分别为Mitscheerlich、Schumacher和Gompertz方程。参数估计值表明:在现有立地水平下,广东省樟树直径、树高和材积的自然生长极值分别为45.91 cm、15.81 m和1.86 m3,在后续的森林经营管理中,唯有适时调整林分密度,加强土壤肥力,才可获得更高的单株立木生长量。
3.2.2 模型的检验
模型的检验采用樟树胸径、树高和材积最优模型的理论值与实际值进行线性拟合,最后利用R2、RMSE、MAE进行预测精度比较。验证结果分析表明,与实际观测值相比,模型拟合的樟树胸径、树高和材积结果都非常显著(见表4和图5,P<0.01),胸径的R2值为0.73,树高的R2值为0.74,材积的R2值为0.77。
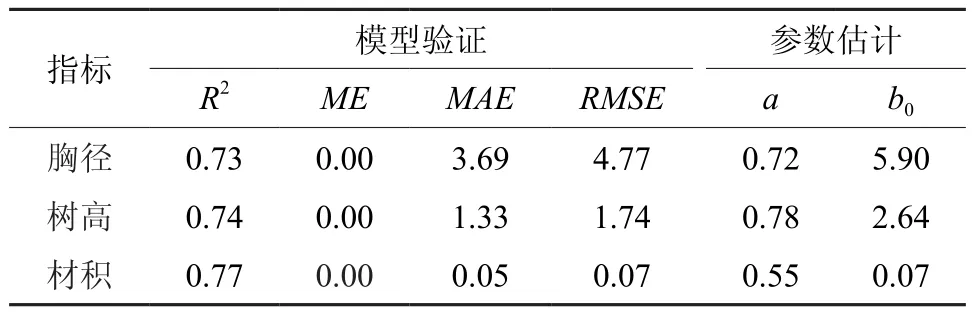
表4 樟树胸径、树高和材积实测值与理论值线性拟合检验结果Table 4 Evaluation of the actual-theoretical value linear fitting of DBH, height and volume for Cinnamomum camphora

图5 樟树胸径、树高和材积最优模型评估图Fig.5 Evaluation model of optimal model for DBH, height and volume of Cinnamomum camphora
4 结论与讨论
4.1 结 论
(1)44 a樟树(去皮)直径可达30.86 cm,树高可达14~15 m,材积可达0.43 m3。
(2)解析木在整个生长期内的生长过程符合树木生长的一般规律,即直径平均生长量的最大值比连年生长量的最大值出现的晚,在树龄29 a时,平均生长量与连年生长量相等。樟树前18年树高平均生长量均大于0.50 m,18 a后平均生长量为0.35 m。樟树树高整个生长期连年生长量与平均生长量多次相交,呈多峰状。樟树材积平均生长量和连年生长量均随年龄的增加而增加,平均生长量和连年生长量高峰期出现的时间比较接近,这与直径和树高的生长高峰期出现的规律不一致,樟树到44 a时还未达到成熟年龄,亦不能确定其主伐年龄。
(3)拟合出樟树胸径、树高和材积的最优生长模型分别为Mitscheerlich、Schumacher和Gompertz方程模型,其表达式依次为:,表明樟树胸径、树高、材积的自然生长极值分别为45.91 cm、15.81 m、1.86 m3。
(4)模型的检验采用樟树胸径、树高和材积最优模型的理论值与实际值进行线性拟合,验证结果分析表明,与实际观测值相比,模型拟合的樟树胸径、树高和材积结果都非常显著,胸径的R2值为0.73,树高的R2值为0.74,材积的R2值为0.77。
4.2 讨 论
(1)44 a樟树(去皮)直径可达30.86 cm,树高可达14~15 m,材积可达0.43 m3。这与龙汉利[18]研究的四川香樟人工林研究结果略有差异,原因可能是由于地理差异、解析木大年龄样本量较少或是解析木数据量的大小影响了研究结果。
(2)拟合出樟树胸径、树高和材积的最优模型,得出在广东现有立地水平下自然生长极值分别为45.91 cm、15.81 m、1.86 m3,这与广东二元立木材积表计算出的结果差异不大。由于采样受局限性,自然生长极值可能不能完全反映特大年龄樟树的情况。模型模拟精度除了受调查样本数据量的影响外,模拟对象、调查时间与方程的形式也会对模拟精度产生非常重要的影响[19]。为了提高本研究模型拟合的精度,可通过增加调查平行样本量、增加年龄较大调查样本量的模拟数据,达到提高模拟精度的目的。已经有研究显示,同一方程在不同林木生长阶段模拟的精度不同,当方程的模拟精度很高时,适用范围却非常小。因此,研究稳定性比较好、模拟精度较高的方程仍将是未来研究的热点。
(3)本研究成果可对广东省樟树单株生长过程及生长规律提供参考,但在实际情况中由于受到其它各种因素的影响,如立地条件、土壤条件、经营措施、经济条件或其它人为因素等,结果可能会出现差异,所以应因地制宜进行调整与分析。由于大径级样本少等建模局限性,本研究建立的樟树胸径、树高、材积模型可能会存在误差,在使用过程中要注意模型的外延问题。
[1]李 芳,黎祖尧,金志农.樟树育苗技术研究进展[J].江西科学, 2015,33(3):330-334.
[2]梁 机,王志勇,覃 鹄,等.不同贮藏和浸种方式对香樟种子发芽率的影响[J].安徽农业科学,2010,38(6):3231-3233.
[3]张建忠,姚小华,任华东,等.香樟扦插繁殖试验研究[J].林业科学研究,2006,19(5):665-668.
[4]陈东阳.芳樟穗条性状对扦插成活率的影响[J].亚热带植物科学,2012,41(3):69-72.
[5]闫 明,钟章成.铝胁迫对感染丛枝菌根真菌的樟树幼苗生长的影响[J].林业科学,2007,43(4):59-65.
[6]姚迎九,康文星,田大伦.18年生樟树人工林生物量的结构与分布[J].中南林学院学报,2003,23(1):1-5.
[7]黄志宏,田大伦,闫文德,等.城市樟树人工林冠层光合作用的时空特征[J].中南林业科技大学学报,2011,31(1):38-46.
[8]田大伦,罗 勇,项文化,等.樟树幼树光合特性及其对 CO2浓度和温度升高的响应[J].林业科学,2004,40(5):88-92.
[9]田大伦,付晓萍,方 晰,等.模拟酸雨对樟树幼苗光合特性的影响[J].林业科学,2007,43(8):29-35.
[10]张静芬,千怀遂,赖纯佳.我国亚热带樟树的温度适宜性及其变化趋势[J].生态学杂志,2010,29(8):1482-1487.
[11]季 荣,季晓波.香樟主要虫害的发生与防治[J].上海农业科技,2006(1):94-95.
[12]孟宪宇.测树学[M].北京:中国林业出版社, 1986.
[13]唐继新,麻 静,贾宏炎,等.南亚热带珍稀濒危树种格木生长规律研究[J].中南林业科技大学学报,2015,35(7):37-44.
[14]段爱国,张建国,童书振.6种生长方程在杉木人工林林分直径结构上的应用[J].林业科学研究,2003,16(4):423-429.
[15]彭舜磊,王得祥.火地塘林区铁杉生长规律研究[J].西北农林科技大学学报(自然科学版),2008,36(4):83-88.
[16]Pukkala T, Lahde E, Laiho O.Using optimization for fitting individual-tree growth models for uneven-aged stands[J].European Journal of Forest Research,2011,130(5):829-839.
[17]龚直文,亢新刚,顾 丽,等.长白山杨桦次生林生长过程与演替动向分析[J].林业科学研究,2009,22(3):379-384.
[18]龙汉利,梁国平,辜云杰,等.四川香樟人工林生长特性研究[J].四川林业科技,2011,32(4):1-4.
[19]祖笑锋,倪成才,Gorden Nigh,等.基于混全效应模型及EBLUP预测美国黄松林分优势木树高生长过程[J].林业科学,2015, 51(3):25-33.
猜你喜欢
广西林业科学(2022年3期)2022-07-08
林业科学研究(2021年6期)2022-01-05
内蒙古林业调查设计(2021年5期)2022-01-05
林业科技情报(2021年2期)2021-07-13
农民致富之友(2017年4期)2017-04-10
现代农业科技(2017年4期)2017-04-10
绿色科技(2016年23期)2017-03-15
绿色科技(2017年1期)2017-03-01
湖北农业科学(2014年3期)2014-07-21
卷宗(2012年5期)2012-10-21
