
非生长季糖槭树干液流特征及影响因子分析
2018-07-04 10:50李明丹王阿晴汤祖翔吴闰生周婧涵
四川林业科技 2018年3期
李明丹,王阿晴,汤祖翔,吴闰生,周婧涵,王 伟,刘 华
(安徽农业大学林学与园林学院 安徽 合肥 230036)
水分在大气—森林—土壤循环中的一个重要过程是植物的蒸腾作用。林分蒸腾量在森林水循环中占有很大比重[1],蒸腾作用消耗树木从土壤中吸收水分的 90%[2],随着环境因子的变化,树木的蒸腾速率在不同时段、不同季节、不同的天气条件下均会发生相应的改变。树干液流是植物体内由于叶片的蒸腾失水并带动树木体内水分通过木质部运输到叶片的过程[3]。热扩散法是在不影响树木自然生长的情况下可连续准确的监测树干液流的方法[4],通过测定树干液流,可以反映出林木蒸腾耗水的变化[5]。
糖槭(AcersaccharumMarsh.)是槭树科槭属多年生的落叶乔木,原产于北美地区。其树型雄伟,木质坚硬,加之秋季叶色变化丰富,在城市绿化中具有极高的观赏价值和生态价值,在我国东北及南方各省区作为行道树栽培已有近百年的历史。同时,糖槭还是一种木本糖料树种,一般在生长15a后就可以提取枝液,并可加工成糖浆,具有较高的经济价值[6]。目前,我国对糖槭的研究主要在光合生理[7]、引种技术[8]、叶片成分分析[9]等方面,在树干液流方面的研究还未见相关报道。目前有关木本植物树干液流的研究主要集中在生长季[10],对于非生长季的树干液流变化研究相对较少,不同生长季节的树干液流速率是否存在一定的相似性,非生长季的树干液流存在意义如何仍未明确。本文基于EMS 62包裹式植物径流测量系统,通过连续监测4 a生的糖槭在秋冬季节的树干液流速率,分析其日变化和季节变化规律,以及水分运移过程中的分配特征,明确太阳辐射、土壤湿度、空气湿度等环境因子对其的影响,为准确评估森林水分蒸腾量对引进的加拿大糖槭在城市森林水分循环的作用,并为该树种在培育管理及开发利用等方面提供一定的理论基础。
1 研究区概况
试验地位于安徽农业大学的农翠园内的糖槭苗圃地。该地属亚热带季风气候,年平均气温15.7 ℃,7月平均气温27.5 ℃,冬季月平均气温1.5 ℃~5 ℃,年平均降雨量近1 000 mm,多集中在5月~6月,年均日照2 100 h左右,年平均相对湿度约为77%,年平均无霜期约228 d。
2 试验材料与方法
2.1 试验材料
试验材料为生长良好、无病害的4a生糖槭苗木,其特征见表1。
表1试验样木情况
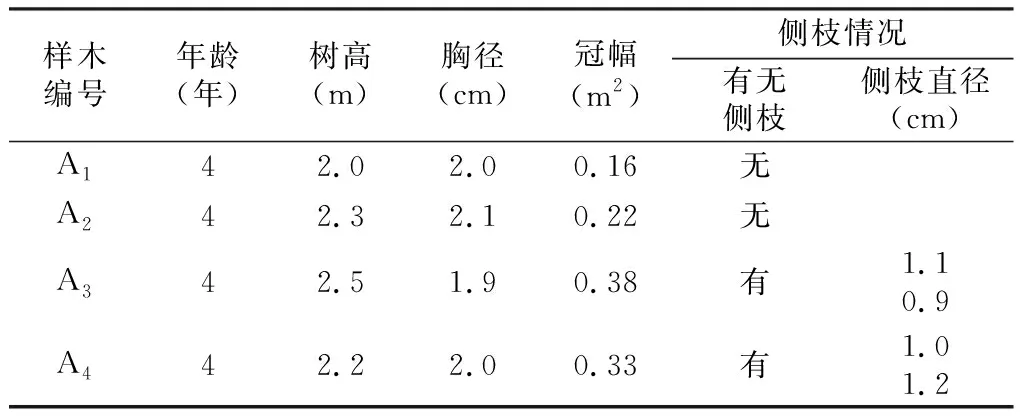
Tab.1 Characteristics of sample trees
2.2 试验方法
于2016年9月,对已选择的4株样树进行Sap Flow Set EMS 62包裹式植物径流测量系统(EMS Brno,Czech Republic)的安装。在树干的阳生面,根据热电偶探针的长度及距离,利用打孔器钻孔,然后将针垂直插入孔中,并用泡沫固定探针后,将其包裹紧实,然后再加上绝热罩,并用胶带密封两端(不要包得太紧,因为茎干在生长),避免水分沿树干流入;同时在绝热罩的下端辐射罩的下端,留一个空隙在线和茎干处,用于排水;然后将探针缆线与数据采集器(DT50)相应接口连接,接通电源。启动测定程序设定为每隔10 min记录一次数据。同时,利用样地内安装的AMS-1000自动气象站,监测大气温度、太阳有效辐射、空气相对湿度、风速、水汽压、土壤温度、土壤水分等指标。于2016年9月至2017年3月连续监测树干液流,每10 d下载一次数据。
2.3 数据处理与分析
选取每个月上、中、下旬任意一天的数据分析不同月份糖槭树干液流速率的动态变化;每个月每天连续监测的液流数据的平均值分析其月变化;以9月份为例,选取A1与A2的上中下旬的晴天、阴天和雨天数据对比分析不同天气条件下液流变化的差异;利用A3和A4的数据分析液流在主干和侧枝之间的分配。用 Excel 2007 整理数据制作图表,SPSS 17.0 进行树干液流和环境因子的相关性分析。
3 结果与分析
3.1 不同月份糖槭树干液流速率的动态变化
糖槭树干液流的动态变化呈多峰型(见图1)。不同月份日间液流和夜间液流发生时段、液流峰值及其液流最大值见表2。9月份到次年3月份,日液流的发生均在20:00时结束,但11月、2月和3月的液流启动时间较其他月份有滞后现象。日液流速率和夜间液流速率的变化基本同步。?其日液流的最大值和夜间液流速率平均值均呈现先下降再增加的趋势。其中,日液流速率在11月份液流速率下降明显,次年1月份的液流速率最大值最小,次年2月份开始液流速率快速增加。夜间液流速率在12月份下降明显,次年1月份最小,次年2月份开始增加。同时,11月和次年2月份的夜间液流持续时间最长,且无零液流时段出现;其他月份均有时间长短不一的零液流速率存在。

图1 不同月份糖槭树干液流的日变化Fig.1 Diurnal variation of sugar maple sap flow in different months
表2不同月份糖槭树干液流变化的关键参数
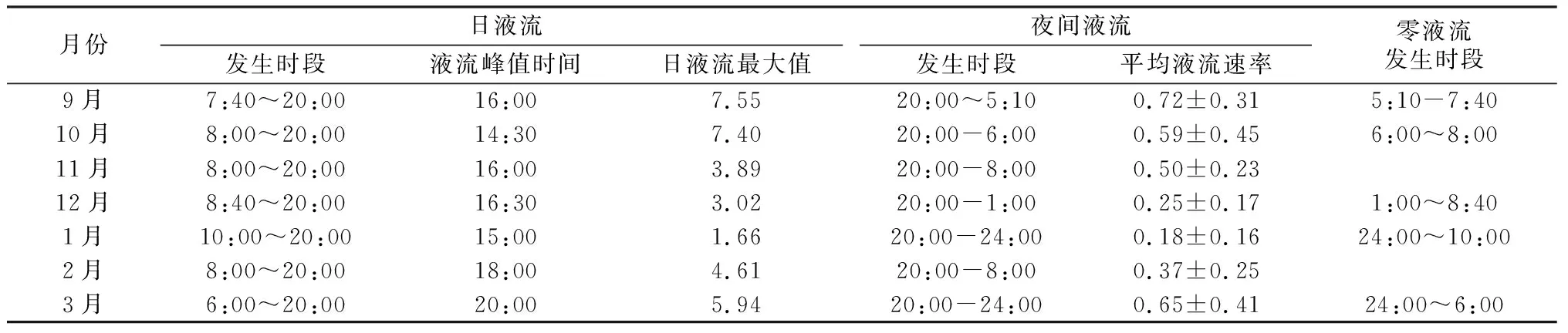
Tab.2 The key parameters of A.saccharum sap flow in different months
3.2 糖槭树干液流速率的月变化
4 a生糖槭树干液流在2016年9月至2017年3月期间的月变化呈现下降趋势(见图2)。其中,9月份糖槭的平均树干液流速率最大,其值为5.784 cm·h-1;10月份平均树干液流降为2.126 cm·h-1,较9月份减少了63%;11月份的平均液流速率为1.431 cm·h-1,较10月份减少了33%;12月份的平均树干液流分别为0.164 cm·h-1,较上个月减少了89%;1月份平均值为0.295 cm·h-1,较12月份增加了80%;2月份的平均树干液流速率为0.296 cm·h-1,较1月增加了3%;3月份的平均树干液流速率分别为0.421 cm·h-1,较2月增加了29.7%。
3.3 不同天气条件下糖槭树干液流速率的比较
3.3.1 不同天气条件下的糖槭树干液流日变化
以2016年的9月份为例,分析晴天、阴天和雨天三种典型不同天气条件下4 a生糖槭树干液流速率的日变化见图3。晴天树干液流速率变化最大,阴天次之,雨天最小。晴天、阴天、雨天时的液流速率变化曲线则呈单峰型,雨天时的液流日变化相对比较弱,但所有树干液流速率均呈“昼高夜低”的变化趋势。晴天条件下,4 a生糖槭树干液流在日间在7:40开始启动,8:00之后迅速升高,在11:00时达到最大值(5.539±1.002)cm·h-1,在11:00~13:00间保持较高的液流速率,之后有所下降,到16:00时树干液流迅速下降,19:00时的液流值很微弱,并在20:00~5:10时段内维持较弱的夜间液流,其液流速率在0.024 g·h-1~0.426 cm·h-1之间。阴天条件下,糖槭树干液流变化曲线均呈单峰型,夜间液流明显。其日间的液流速率较大的时段主要发生在11:00~15:00时之间,在12:00时左右的液流速率最大。雨天条件下,4a生糖槭树干液流明显降低,且无明显的夜间液流,日也留的变化主要发生在5:00~8:00时段内,7:00时达到最大值(1.037±0.201) cm·h-1。
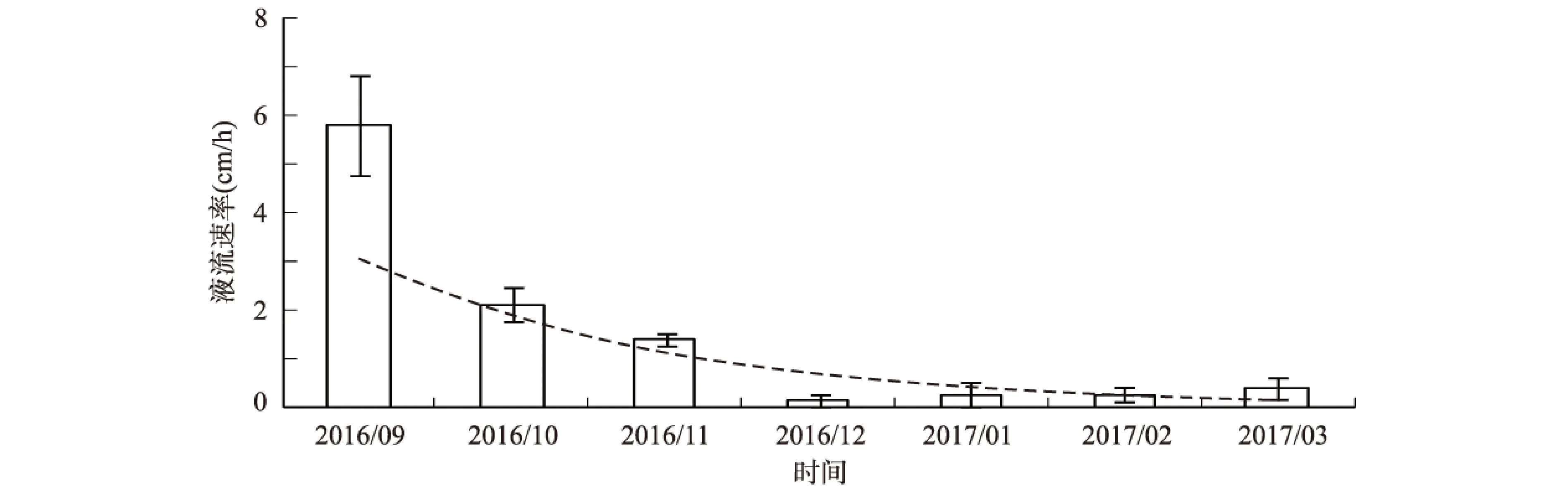
图2 糖槭树干液流的月变化Fig.2 Monthly change of A.saccharum sap flow
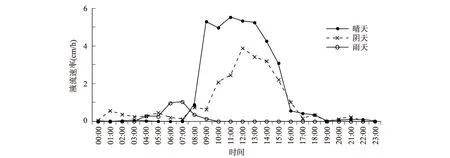
图3 不同天气条件下的糖槭树干液流日变化特征Fig.3 Characteristics of A.saccharum sap flow in sunny,cloudy,and rainy days
3.3.2 不同天气条件下不同树体位置对树干液流速率的影响
不同位置的树干液流分配及日变化见图4。晴天条件下,在树体的不同位置,其树干液流速率的峰值大小和到达峰值的时间均不相同;到达峰值的时间顺序为主干>小枝干>大枝干,峰值大小为主干>大枝干>小枝干。阴天条件下,不同树体位置的树干液流速率变化也呈单峰型,到达峰值时间为主干>大枝干=小枝干,峰值大小为大枝干>主干>小枝干。雨天条件下,主干树干液流速率变化呈单峰型,两侧枝树干液流为多峰型,其树干液流速率相对较低;其达到液流速率到达最大值的时间顺序为主干>大枝干>小枝干,峰值大小为主干>大枝干>小枝干。
3.4 不同环境因子对树干液流速率的影响
外界环境因子的波动在树体内部的液流变化上得到响应。整个非生长季内糖槭树干液流速率与环境因子的相关关系见表3。与太阳辐射强度、空气温度极显著正相关,与空气湿度、土壤湿度极显著负相关。影响糖槭树干液流速率的环境因子排序为:土壤湿度>空气温度>空气湿度>太阳辐射。
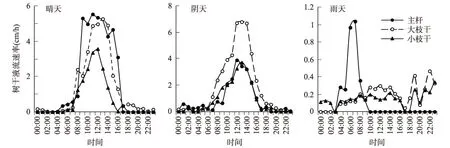
图5 不同树体位置树干液流分配及日变化Fig.4 Distribution and diurnal variation of the sap flow in different tree positions
表3糖槭树干液流速率与环境因子相关分析
Tab.3 Correlation analysis betweenA.saccharumthe sap flow and environmental factors

项目太阳辐射空气温度空气湿度土壤湿度液流速率Pearson Correlation0.050**0.065**-0.051**-0.293**Sig.(2-tailed)0.0010.0000.0010.000
注:**P<0.01;N=4185;Pearson Correlation相关系数;Sig.( 2-tailed) -P 值( 双侧)
4 讨论
糖槭树干液流速率昼夜变化特征都呈现“昼高夜低”。赵培强[11]等对华南地区常见树种的树干液流速率在干湿季变化其日变化格局均为“昼高夜低”变化格局。这主要是受光合有效辐射的影响,在不同的地域,不同树种的树干液流速率均呈现此种变化趋势。糖槭树体不同位置均存在大小不同的夜间树干液流速率,夜间液流主要用于夜间补水、夜间蒸腾和养分运输,以维持自身水分平衡[12,13]。晴天条件下不同树体位置糖槭树干液流速率均呈单峰曲线,这与柳杉[14]、枸杞[15]、木荷[16]、荔枝[17]和云杉[18]的晴天液流曲线变化趋势一致,均呈单峰曲线。糖槭树干液流在晴天,液流启动时间早,持续时间长,液流量大,变化幅度也大,阴天和雨天的液流量变化相差不大,启动时间晚,持续时间短,且雨天液流量变化幅度最小,液流量相对稳定。原因在于雨天时太阳辐射较弱,空气湿度相对较大,气温较低,糖槭蒸腾作用与晴天相比较弱,其树干液流量相对稳定,变化幅度较小。
糖槭月平均树干液流速率变化均为下降趋势,与叶面积指数的季节变化有很大关系[19]。9~11月变化较明显,主要是因为9~11月内糖槭落叶速度快,叶面积指数快速下降,同时日照时间变短,气温逐渐降低,雨雪天气增多,因而导致蒸腾作用变小,树干液流速率随之变小,减小到较低水平后开始进入休眠状态。12~2月份内基本保持不变,是因为糖槭叶片完全凋落进入休眠期,存在少量液流速率维持自身水分平衡及养分运输。1月份的液流出现回升,这为新芽萌发运输营养,也可能是1月份天气开始回暖,维持自身水分平衡的需求增加。
糖槭树干液流速率受太阳辐射强度、空气温度、空气湿度、土壤湿度等环境因子的显著影响。糖槭树干液流均与太阳辐射呈极显著正相关,因为太阳辐射强度增加叶片气孔导度增大,树木树冠蒸腾量增大,树干液流量也随之增大。糖槭树干液流与空气温度呈极显著正相关,空气温度升高,水分汽化速度加快,蒸腾速率加快,树干液流速率随之加快,而空气温度随太阳辐射强度的升高而升高。与空气湿度呈极显著负相关,空气湿度的增加,水气压随之增加,使叶片周围水气压与叶片内部水气梯度减小,水分汽化速度减弱,树干液流速率随之减弱。土壤湿度对植物生长的影响主要表现在影响其呼吸作用和蒸腾作用,当土壤湿度在一定合适范围内,植物正常生长;当土壤湿度过高时,会引起气孔关闭,迫使蒸腾作用停止,而光合速率不受影响;当土壤湿度过低时,植物根系水分吸收会受到抑制,引发植物水分亏缺,进而导致气孔关闭避免水分继续亏缺[15]。本研究中,非生长季内叶片凋落导致的树木蒸腾降低,因此糖槭树干液流速率也与根部土壤湿度表现出极显著的负相关关系。
参考文献:
[1] Jasechko S,Sharp Z D,Gibson J J,et al.Terrestrial water fluxes dominated by transpiration [J].Nature,2013,496(7445):347.
[2] Ortuno M F,Garciaorellana Y,Conejero W,et al.Stem and leaf water potentials,gas exchange,sap flow,and trunk diameter fluctuations for detecting water stress in lemon trees[J].Trees,2006,20(1):1~8.
[3] 赵春彦,司建华,冯起,等.树干液流研究进展与展望[J].西北林学院学报,2016,30(5):98~105.
[4] Ballester C,Castel J,Testi L,et al.Can heat-pulse sap flow measurements be used as continuous water stress indicators of citrus trees?[J].Irrigation Science,2013,31(5):1053~1063.
[5] Steppe K,Vandegehuchte M W,Tognetti R,et al.Sap flow as a key trait in the understanding of plant hydraulic functioning[J].Tree Physiology,2015,35(4):341~345.
[6] 许易梅.复叶槭(糖槭)的应用现状及发展前景的综合评价[D].东北林业大学,2004.
[7] 闻婧,张俊,孟力力,等.夏季遮光对糖槭光合特性和叶绿素荧光动力学特征的影响[J].江苏农业科学,2015(8):189~190.
[8] 马艳丽,王鹏.糖槭的应用价值与栽培技术[J].黑龙江农业科学,2013,(5):154~155.
[9] 赵宏.糖槭叶化学成分及抗氧化、抗炎活性研究[D].佳木斯大学,2008.
[10] 马履一,王华田,林平.北京地区几个造林树种耗水性比较研究[J].北京林业大学学报,2003,25(2):1~7.
[11] 赵培强,赵平,牛俊峰,等.华南地区常见树种导管特征与树干液流的关系[J].热带亚热带植物学报,2014,(6):537~548.
[12] 赵春彦,司建华,冯起,等.胡杨夜间液流通量及其影响因子研究[J].干旱区研究,2015,32(6):1173~1180.
[13] 王华田,赵文飞,马履一.侧柏树干边材液流的空间变化规律及其相关因子[J].林业科学,2006,42(7):21~27.
[14] 蒋文伟,杨广远,赵明水,等.天目山柳杉树干液流的昼夜及季节变化[J].南京林业大学学报自然科学版,2012,36(5):77~80.
[15] 徐利岗,苗正伟,杜历,等.干旱区枸杞树干液流变化特征及其影响因素[J].生态学报,2016,36 (17):5519~5527.
[16] 周翠鸣,赵平,倪广艳,等.广州地区木荷夜间树干液流补水的影响因子及其对蒸腾的贡献[J].应用生态学报,2012,23(7):1751~1757.
[17] 凡超,邱燕萍,李志强,等.荔枝树干液流速率与气象因子的关系[J].生态学报,2014,34(9):2401~2410.
[18] 万艳芳,于澎涛,刘贤德,等.祁连山青海云杉树干液流密度的优势度差异[J].生态学报,2013,37(9):1771~1778.
[19] 常学向,赵文智.荒漠绿洲农田防护树种二白杨生长季节树干液流的变化[J].生态学报,2004,24(7):1436~1441.
猜你喜欢
东坡赤壁诗词(2022年4期)2022-10-30
天津农林科技(2020年3期)2020-08-13
意林(2020年9期)2020-06-01
领导文萃(2020年7期)2020-05-15
电子制作(2019年15期)2019-08-27
高原山地气象研究(2016年4期)2016-02-28
高原山地气象研究(2016年4期)2016-02-28
电源技术(2016年9期)2016-02-27
中国质量与标准导报(2014年5期)2014-02-28
储能科学与技术(2014年5期)2014-02-27
