
灌浆期低温对离体培养玉米强弱势粒发育的影响
2018-06-29 08:24:46张巽郝建平王璞张萍陈璐洁
中国农业科学 2018年12期
张巽,郝建平,王璞,张萍,陈璐洁
灌浆期低温对离体培养玉米强弱势粒发育的影响
张巽1,2,郝建平1,王璞3,张萍3,陈璐洁3
(1山西农业大学农学院,山西太谷 030801;2大同大学生命科学学院,山西大同 037009;3中国农业大学农学院,北京 100193)
【目的】通过对玉米灌浆过程中籽粒干物质、淀粉的积累,籽粒内源激素含量及淀粉积累相关酶活性的研究,揭示低温对灌浆过程中玉米强、弱势籽粒灌浆生理过程的影响规律,为生产上抗御低温冷害提供理论参考。【方法】选用郑单958为试验品种,采用玉米籽粒离体培养的方式,将大田人工授粉后3 d的玉米果穗按照弱势粒和强势粒进行取样,无菌环境接种到人工培养基培养,低温处理和对照分别设置培养平均温度为16℃及25℃。自授粉后每10 d取样一次,分别测定灌浆过程中玉米强、弱势粒干物质积累量、内源激素、籽粒淀粉含量及淀粉积累相关酶活性。【结果】低温胁迫下强、弱势粒灌浆后期粒重分别比对照低47.58%、50.95%,强、弱势粒灌浆高峰期的平均灌浆速率较对照分别显著降低55.39%、54.72%。低温胁迫下玉米籽粒灌浆速率前期提升和后期减小的速度明显减缓,活跃灌浆时间延长5—7 d。授粉后10 d低温处理显著降低玉米强、弱势粒生长素(IAA)、玉米素(ZR)和脱落酸(ABA)含量,显著提高玉米籽粒赤霉素(GA3)的含量。授粉后30 d低温处理显著降低了弱势粒的IAA、ZR含量,增加了强势粒的ABA含量。低温胁迫显著减弱了灌浆前期和灌浆中期的可溶性酸性转化酶(SAI)、蔗糖合酶(SS)、淀粉合酶(SSS)及ADPG焦磷酸化酶(APGase)的活性,低温下弱势粒SAI活性降幅大于强势粒,对SS、SSS及APGase活性的降低幅度表现为强势粒大于弱势粒,导致玉米籽粒淀粉含量降低。【结论】受低温胁迫影响,灌浆前期玉米籽粒的IAA、ZR、ABA含量减少,GA3含量增加,SAI活性降低,导致籽粒库容量减少,库活性不足。在灌浆中期,低温降低SS活性,造成淀粉合成底物供应不足,影响淀粉的合成,降低籽粒淀粉含量。低温处理降低玉米强、弱势粒的灌浆速率,导致籽粒干物质积累减少。低温对玉米强弱、势粒的灌浆过程都造成较大的影响,且对弱势粒的影响大于强势粒。
玉米;低温;离体培养;灌浆期;激素
0 引言
【研究意义】在气候变暖成为全球性气象问题的背景下,极端天气事件的发生频率仍呈现增加趋势[1-2],局部地区的农业气象灾害事件多发[3],低温冷害成为全国性农业灾害,在玉米北方产区发生得尤为严重和频繁[4]。玉米属于喜温作物,低温对其生长发育影响较大,我国北方玉米产区每3—4年就会发生一次低温冷害[5],导致减产20%—30%[6],成为限制玉米产量提高的一个重要因素[7]。生育后期的低温威胁导致玉米灌浆过程不能顺利完成,进而造成减产[8],因此研究灌浆期低温对玉米籽粒生长发育过程的影响,能够为生产中抗御低温冷害提供理论依据。【前人研究进展】研究发现,低温胁迫下玉米的光合性能受到限制,群体产量明显降低[9-12],这一过程是玉米在逆境胁迫下,各组织器官生长发育变化的综合结果。籽粒作为玉米最终形成产量的重要库器官,对温度胁迫较为敏感[13-14],低温对玉米籽粒发育及内部的生理机制影响显著[15]。前人研究发现低温胁迫扰乱了作物内源激素平衡[16-18],降低了植株体内淀粉和蛋白质等大分子物质的含量,增加了可溶性糖和游离氨基酸等小分子物质含量[19]。淀粉是玉米籽粒主要成分,而籽粒发育过程中,与淀粉积累合成代谢密切相关的可溶性酸性转化酶(soluble acid invertase,SAI,EC 3.2.1.26),蔗糖合酶(sucrose synthase,SS,EC 2.4.1.13),可溶性淀粉合成酶(soluble starch synthase,SSS,EC 2.4.1.21)和腺苷二磷酸葡萄糖焦磷酸化酶(ADP- glucose pyrophosphorylase,APGase,EC 2.7.7.21)的活性对外界环境条件的变化十分敏感[20],籽粒灌浆期间环境条件的改变将直接影响籽粒中贮存物质的合成[21-22]。玉米籽粒发育过程中,受内源激素及淀粉积累相关酶影响,不同粒位的籽粒发育差异较大,逆境胁迫下弱势粒易产生败育[23-24]。【本研究切入点】在逆境胁迫下,玉米果穗不同粒位籽粒因库活性的差异,强、弱势粒生长发育存在明显差异,直接关系到玉米粒重和产量的高低。目前低温胁迫对灌浆期玉米不同粒位籽粒生长发育影响的研究较少,进一步揭示低温胁迫下玉米强、弱势粒的物质积累和库活性的变化有重要意义。【拟解决的关键问题】本文通过玉米籽粒离体培养的方式,利用人工气候箱创建低温环境,研究低温对玉米灌浆过程中强、弱势粒干物质和淀粉的积累,以及内源激素含量和淀粉积累相关酶活性变化的影响。揭示低温胁迫下玉米籽粒灌浆过程中籽粒物质积累及内源激素、淀粉积累相关酶的变化规律,为生产上采取有效措施防控低温冷害提供依据。
1 材料与方法
1.1 试验材料
本试验选用郑单958为试验材料,2015年6月15日播种于中国农业大学北京上庄实验站(40°08′ 25″N,116°10′ 39″E,海拔49 m)。试验地为粉砂质潮土,土壤地力中等,基础养分为有机质1.47%,全氮含量0.86 g·kg-1,有效磷22.76 mg·kg-1,速效钾63.06 mg·kg-1。试验田无前茬作物,玉米季外未种植其他作物,常规田间管理。于玉米开花吐丝前,选择生长均匀一致健壮的植株进行雌穗套袋,开花后统一人工授粉。籽粒离体培养试验参考陈素省等[25]的方法,在中国农业大学农业部农作制度重点实验室进行,于3 DAP(day after pollination,DAP)取雌穗回实验室进行籽粒离体培养,离体籽粒取样参考张凤路等[14]的方法。离体籽粒在无菌条件下切穗取样,弱势粒取样部位为雌穗顶部向下1—5 cm,强势粒取样部位为雌穗基部向上第18至20粒位区间,取样后籽粒在无菌操作条件下接种到灭菌培养基。
接种后的籽粒在25℃恒温下暗培养1 d,5 DAP开始温度处理。设置4个处理,分别为LTS(低温处理强势粒)、LTI(低温处理弱势粒),NTS(常温处理强势粒)、NTI(常温处理弱势粒),每个处理4次重复,每个重复至少包含30颗籽粒;培养条件由人工气候箱(BIC-300型,上海博讯)实现,低温处理设置14℃至19℃的梯度温度,平均温度为16℃,对照处理设置22℃至27℃的梯度温度,平均温度为25℃;人工气候箱不设光照,培养过程在黑暗条件下进行,培养环境湿度为85%。
1.2 籽粒离体培养方法
参考Gengenbach[26]的试验方法,果穗接种块按照籽粒与穗轴比例1﹕9取样[27],在无菌操作环境下接种到培养基。培养基制备参考Cully[28]和赵丽晓等[24]的方法,籽粒接种到培养基后,置于人工气候箱进行培养。
1.3 测定项目与方法
1.3.1 籽粒干重分别在10、20、30、40、50 DAP取样,籽粒于105℃杀青30 min,80℃烘干后测定样品干重。
1.3.2 灌浆速率玉米籽粒灌浆过程应用Logistic曲线拟合,曲线方程为W=a/(1+be-ct)。式中,籽粒干重(W)为因变量,授粉后天数(t)为自变量,b、c为常数,a为理论籽粒最大干重。对拟合曲线方程进行求导和积分后,得到籽粒灌浆速率V的方程: V=dWdt=abce-ct/(1+be-ct)2 [29]。
1.3.3 籽粒淀粉含量测定干重后的籽粒研磨成粉,采用酸水解-DNS法[30]测定玉米籽粒淀粉含量。
1.3.4 激素含量的测定取10、30 DAP籽粒鲜样,利用酶联免疫法[31-32]测定籽粒IAA、ZR、GA3和ABA含量,酶联免疫试剂盒由中国农业大学提供,每个样品分别重复测定3次,取平均值。IAA、ZR、GA3和ABA的回收率分别为94.3%、89.2%、90.5%和91.8%。
1.3.5 淀粉积累相关酶活性在10、30 DAP取籽粒鲜样,按照Nakamura[33]和Zinselmeier等[34]的试验方法进行可溶性酸性转化酶(SAI)、蔗糖合酶(SS)、可溶性淀粉合成酶(SSS)和腺苷二磷酸葡萄糖焦磷酸化酶(APGase)4种酶的提取及活性的测定。
1.4 数据分析
采用IBM SPSS Statistics 21软件统计分析数据,采用Duncan’ s法进行显著性检验,文中图表采用Microsoft Excel 2013制作。
2 结果
2.1 灌浆期低温对籽粒干物质积累的影响
2.1.1 籽粒干重变化规律 不同温度处理下,玉米强、弱势粒的干重积累变化规律相同,呈现“S”形曲线变化(图1-A)。灌浆初期,低温胁迫处理与对照处理差异不显著,从20 DAP开始,低温胁迫处理的强、弱势粒干重都显著低于对照处理,且籽粒干重增长速度也慢于对照,50 DAP低温处理强势粒干重较对照低47.58%,弱势粒干重较对照低50.95%,弱势粒干重受低温影响幅度大于强势粒。
2.1.2 利用Logistic方程模拟籽粒灌浆 利用Logistics方程W=a/(1+be-ct)对籽粒物质积累的动态规律进行模拟,方程拟合后参数见表1。各处理拟合方程经统计检验,方程决定系数都在0.98以上,均达到极显著水平,方程拟合度较好。受低温影响,玉米籽粒活跃灌浆期时间延长,其中强势粒延长7.24 d,弱势粒延长5.77 d。低温胁迫下,玉米籽粒最大灌浆速率和平均灌浆速率均减小,强势粒籽粒最大灌浆速率减少58.30%,弱势粒减少58.85%,强势粒籽粒平均灌浆速率减少55.89%,弱势粒减少57.57%。
2.1.3 籽粒灌浆速率的变化规律 由图1-B可知,各处理强、弱势粒的灌浆速率都随着灌浆时间的增加而加快,峰值出现在30 DAP左右,随即呈现下降趋势,在50 DAP达最低值,强、弱势粒有相似的变化规律。在10—40 DAP,低温处理降低了籽粒灌浆速率,强、弱粒均表现为随灌浆时间延续,灌浆速率呈先增加后减小的变化规律,但峰值出现时间要晚于对照处理。在30 DAP,低温处理强势粒和弱势粒灌浆速率均低于对照处理。在50 DAP,低温处理强、弱势粒灌浆速率均高于对照处理。可见,籽粒经低温处理后,灌浆前期的灌浆速率增加缓慢,而且后期灌浆速率降低也缓慢,表现为籽粒灌浆持续时间的延长。
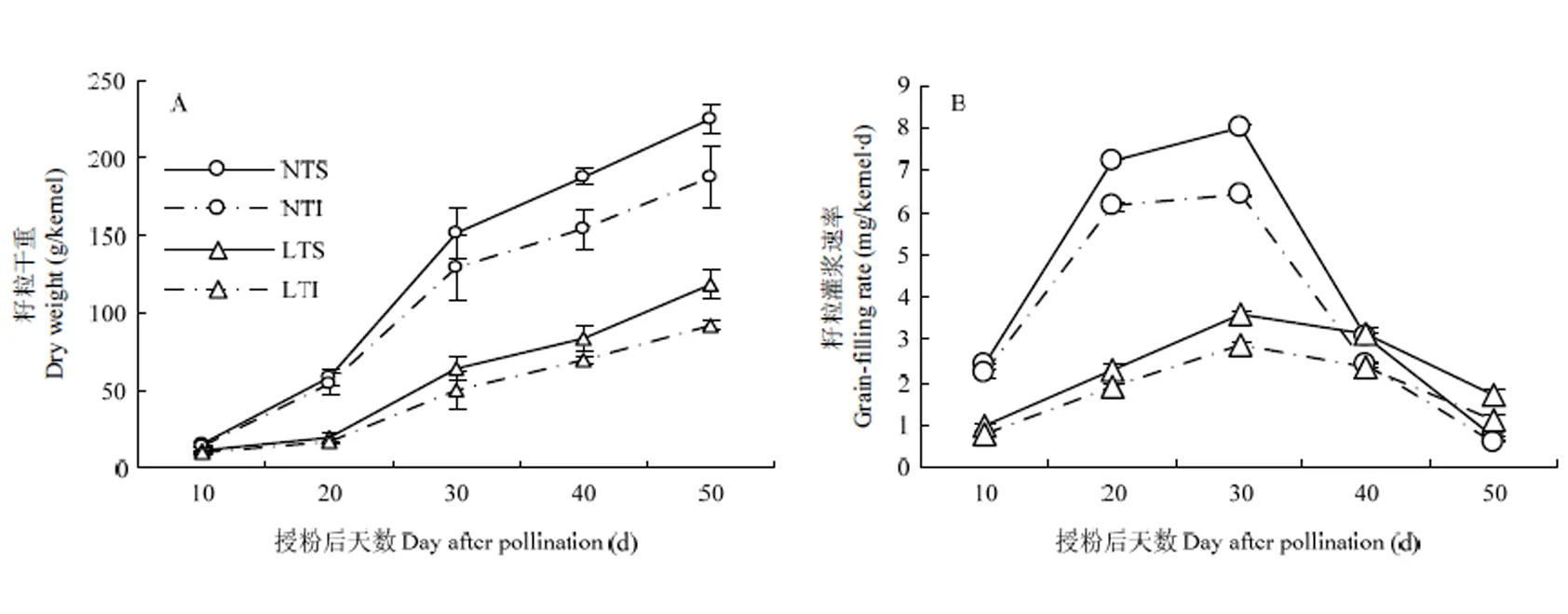
NTS:强势粒常温处理;NTI:弱势粒常温处理;LTS:强势粒低温处理;LTI:弱势粒低温处理。下同
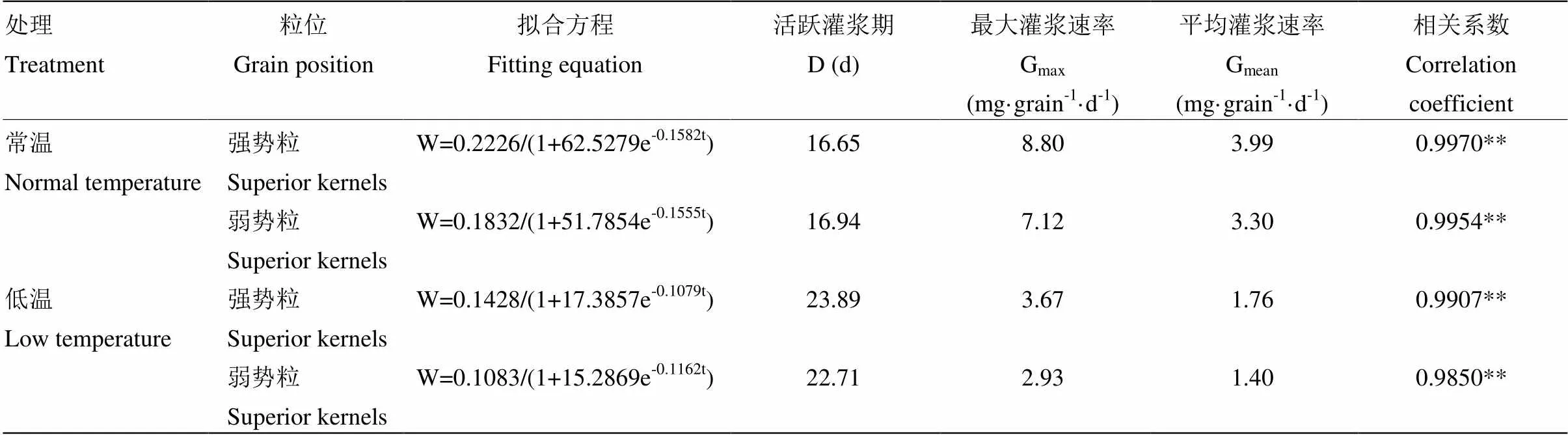
表1 玉米灌浆期低温下籽粒灌浆模型与灌浆参数
D:籽粒活跃灌浆天数;Gmax:籽粒最大灌浆速率;Gmean:平均灌浆速率;*,**为相关系数分别达到0.05和0.01显著性水平
D: Duration of active grain-filling; Gmax: Maximum of grain-filling rate; Gmean: mean of grain-filling rate; *, ** mean significant difference at 0.05 and 0.01 levels
2.2 灌浆期低温对籽粒内源激素含量的影响
玉米籽粒内源激素的含量受低温胁迫后发生明显改变(图2)。在10DAP,低温处理后,籽粒IAA含量显著低于对照处理,且弱势粒降低幅度大于强势粒;GA3的含量均高于对照,其中强势粒差异显著,弱势粒差异不显著;ZR及ABA的含量均表现为强、弱势粒显著低于对照,且弱势粒减低幅度大于强势粒。在30 DAP,低温处理下强势粒IAA含量与对照差异不明显,而弱势粒的IAA含量显著低于对照;GA3含量受低温影响较小,强、弱势粒均与对照差异不显著;ZR含量表现为强势、弱势粒均低于对照,但强势粒差异不显著,而弱势粒差异显著;ABA含量受低温影响,强、弱势粒均高于对照处理,且处理间差异显著,其中强势粒增加幅度大于弱势粒,随着低温胁迫时间的累加,低温对玉米籽粒内源激素含量的影响依然存在,且对弱势粒的影响更加明显。可见,低温引起了籽粒内源激素含量的紊乱,严重影响了受多种激素调控的籽粒灌浆过程。
2.3 灌浆期低温对籽粒淀粉含量的影响
低温胁迫下,玉米强、弱势粒淀粉含量变化趋势基本一致(图3)。低温处理强、弱势粒淀粉含量均低于常温处理,在整个灌浆过程中其变化趋势与籽粒干重及灌浆速率表现保持一致。在10 DAP低温处理下,强、弱势粒淀粉含量与常温处理差异不显著,从20 DAP开始,低温处理下强弱势粒淀粉含量均显著低于常温处理,且20 DAP强、弱势粒淀粉含量较常温处理降低幅度最大,其中强势粒降低65.97%,弱势粒降低66.35%。随灌浆时间推进,从30 DAP开始,各处理间淀粉含量差距呈逐渐增大趋势,在50 DAP达到最大,低温胁迫下,强势粒比常温低53.27%,弱势粒比常温低56.32%,说明低温处理对籽粒最终淀粉含量的影响,弱势粒大于强势粒。

玉米标注不同小写字母柱值在0.05水平上差异显著。下同

图3 玉米灌浆期低温胁迫下籽粒淀粉含量的变化
2.4 灌浆期低温对籽粒积累相关酶活性的影响
从图4可知,受低温胁迫影响,玉米籽粒灌浆前期及灌浆中期的酸性蔗糖转化酶(SAI)和蔗糖合酶(SS)的活性均低于对照,灌浆期玉米籽粒淀粉合成底物转化效率因低温胁迫而受到限制。在10 DAP,低温处理强、弱势粒的SAI、SS活性均显著低于对照,强势粒的两种酶活性降低程度均大于弱势粒。在30 DAP,低温处理强势粒的SAI及SS活性显著低于对照,其中弱势粒SAI活性的降低幅度大于强势粒,而SS活性降低幅度小于的强势粒。低温胁迫下,玉米籽粒可溶性淀粉合酶(SSS)和ADPG焦磷酸化酶(APGase)活性显著低于对照处理(图4),导致玉米籽粒淀粉的合成过程受限。在10 DAP,低温处理强、弱势粒的SSS、APGase活性与对照相比较均有大幅下降,表明在灌浆前期,低温对强、弱势粒的两种酶的影响较大。在30 DAP,低温处理强势粒的SSS、APGase活性显著低于对照处理,弱势粒SSS活性显著低于对照,而AGPase活性与对照差异不显著。在灌浆中期,低温对SSS、AGPase两种酶活性的影响表现为强势粒大于弱势粒,且对SSS活性的影响大于对APGase活性的影响。
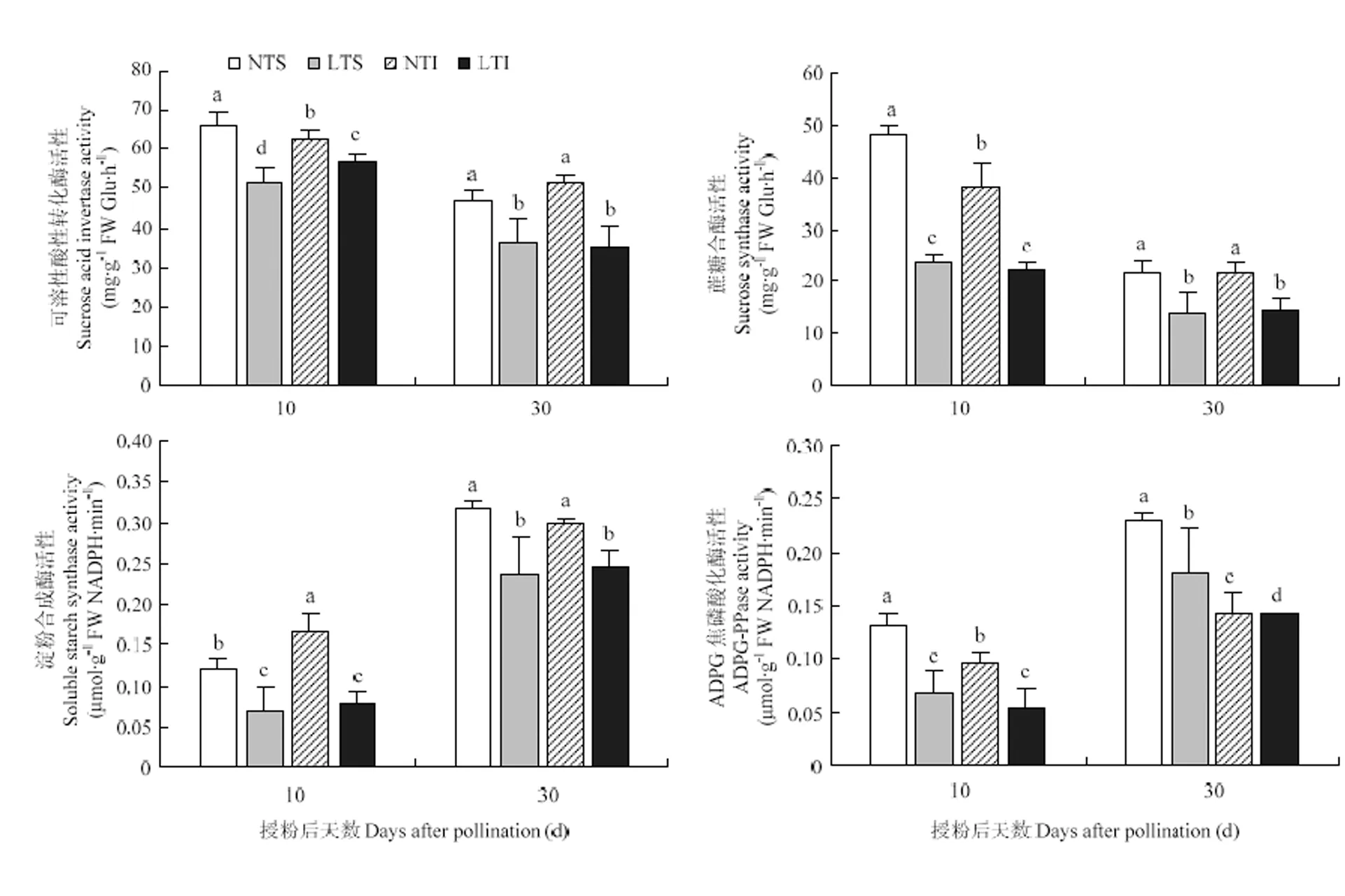
图4 玉米灌浆期低温胁迫下籽粒淀粉合成相关酶活性的变化
2.5 灌浆期低温处理下玉米籽粒灌浆速率、籽粒内源激素含量及淀粉积累相关酶活性间的相关分析
通过灌浆速率(V)模拟方程计算10、30 DAP的强、弱势粒灌浆速率,并与10 DAP和30 DAP强、弱势粒内源激素含量及淀粉积累相关酶活性进行相关分析,结果见表2。由表可知,低温处理后,玉米籽粒灌浆速率与GA3含量呈显著负相关,与SSS、AGPase活性呈极著正相关。各激素含量之间,GA3与IAA及ZR呈显著负相关,IAA与ZR呈显著正相关。各淀粉积累相关酶活性之间,SSS与AGPase呈极显著正相关。各激素含量与淀粉合成酶活性之间,SSS、AGPase与GA3呈极显著负相关,与IAA及ZR呈显著正相关。可见,低温降低玉米的灌浆速率,可能与GA3含量的增加及SSS、AGPase活性的降低有关。
3 讨论
灌浆过程是玉米产量形成的重要时期,也是籽粒发育和物质积累的关键时期,该阶段对环境温度要求较高,最适宜的灌浆温度为日均22℃—24℃[35]。灌浆过程对低温比较敏感[36],温度低于18℃,就会受到限制[37]。低温对灌浆前期影响最大[38],该时期是玉米产量受温度胁迫影响的敏感期[39]。研究表明,灌浆期低温降低籽粒灌浆速率,进而延缓了整个灌浆进程[40],最终导致籽粒物质积累不足,温度每降低1℃,玉米减产300 kg·hm-2[41]。本研究发现,低温能够延长玉米籽粒的活跃灌浆时间6—7 d,但同时也降低了灌浆速率,且灌浆时间的延长无法补偿灌浆速率的降低。遭遇低温后,玉米灌浆前期的籽粒灌浆速率受影响较大,授粉后0—20 d,低温处理强、弱势粒均表现出灌浆速率增长缓慢,峰值低于对照,且峰值出现时间晚于对照的现象,从而降低了籽粒干物质的积累效率,最终导致籽粒干重的显著降低,这与笔者前期研究结果一致[42]。
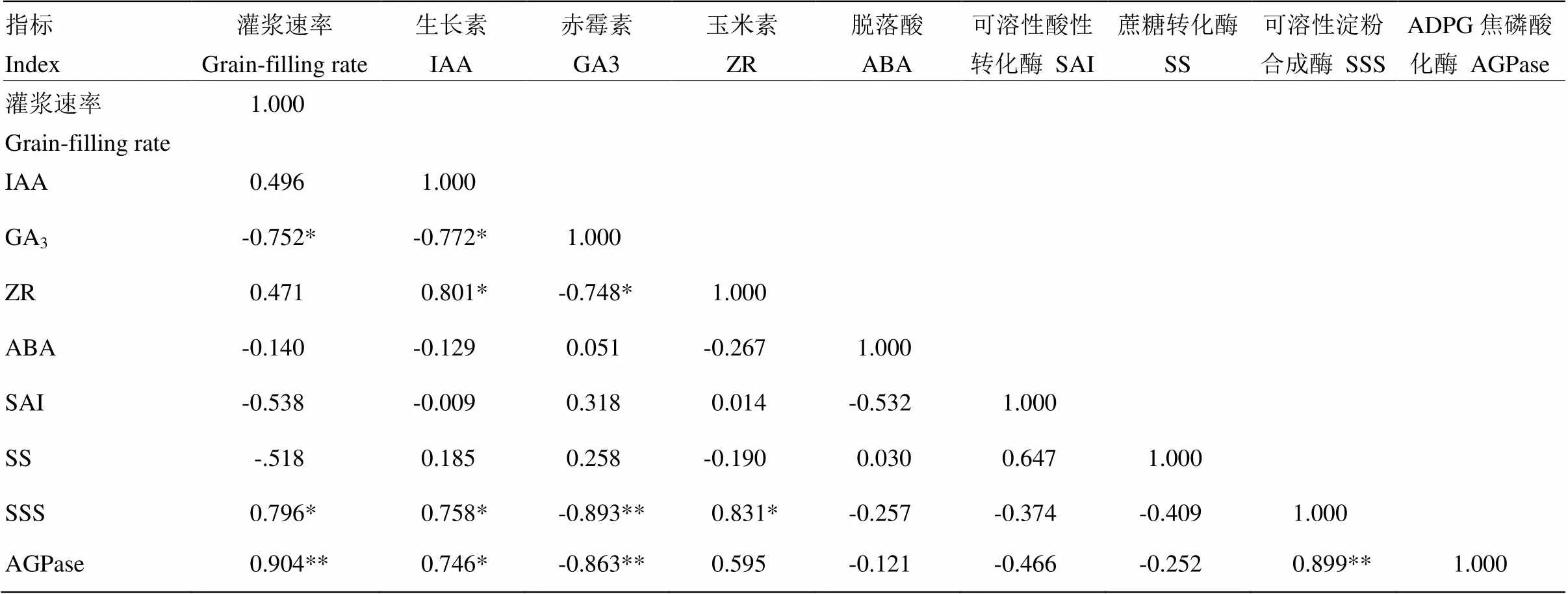
表2 灌浆期低温下玉米籽粒灌浆速率与籽粒内源激素含量及淀粉积累相关酶活性变化的相关分析
*代表0.05差异显著水平,**代表0.01差异显著水平
* indicated significant at 0.05 level; ** indicated significant at 0.01 level
植物内源激素对玉米籽粒的发育起着非常重要的调节作用[18],其对逆境胁迫异常敏感,低温扰乱了籽粒中内源激素平衡,干扰了籽粒正常的生长发育过程[16, 43]。前人在对玉米籽粒的灌浆速率及胚乳细胞增殖速度的研究中发现,在籽粒灌浆前期,这两项指标与籽粒中ZR、IAA及ABA的含量呈显著正相关,在活跃灌浆期与籽粒中GA3的含量呈显著负相关[44]。在本研究中,同样发现在灌浆前期,籽粒灌浆速率与GA3含量表现为显著负相关,籽粒GA3含量因低温胁迫而显著提高,其中强势粒显著提高24.7%,而弱势粒提高不显著。籽粒的IAA、ZR及ABA的含量因低温胁迫而显著降低,其中,强势粒3种激素分别降低了44.1%、39.3%、5.9%,弱势粒分别降低了56.6%、34.6%、12.1%,低温胁迫引起内源激素的变化,是导致灌浆初期籽粒灌浆速率低于对照的主要原因之一。研究表明,胚乳细胞的平均增殖速率和最大胚乳细胞数与ZR高含量呈显著正相关,玉米籽粒胚乳细胞的分裂受ZR的调控[45]。在本研究中,灌浆初期受低温影响,玉米籽粒GA3含量升高而ZR含量降低,且GA3含量与ZR含量表现为负相关,这对玉米籽粒胚乳细胞的分裂不利,进而影响玉米籽粒库容的形成,降低了籽粒同化物积累的能力。在灌浆中期,籽粒GA3含量受低温影响变化不大,ABA含量受低温的影响要高于对照处理,但此时玉米灌浆速率低于对照,说明在长时间低温胁迫下,籽粒ABA含量虽然升高,但不能增加籽粒灌浆速率,反而会抑制籽粒灌浆。在灌浆中期,低温处理下玉米强、弱势粒的IAA及ZR含量低于对照,其中强势粒IAA含量比对照低6.7%,弱势粒IAA含量比对照低38.4%,表明在灌浆中期,低温胁迫对弱势粒的影响尤为显著,导致弱势粒粒重及灌浆速率受低温影响较强势粒更大。
灌浆期低温导致籽粒干物质充实不良,这与籽粒保持最大灌浆速率时间缩短,最大灌浆速率、平均灌浆速率降低[46],淀粉含量降低关系密切[47]。本研究中,低温胁迫影响了玉米籽粒的淀粉积累,随着低温胁迫时间的延长和危害的累积,在灌浆过程中,低温处理强、弱势粒淀粉含量显著低于常温处理,导致籽粒干物质积累量明显减少。在50 DAP,强势粒重减少了47.6%,弱势粒重减少了50.9%,表明在离体培养条件下,低温对弱势粒干物质积累的影响程度略大于强势粒。
研究表明,灌浆前期是籽粒库容形成的关键时期,也是籽粒库活力最活跃的时期[48],此时玉米灌浆过程受籽粒库活性影响较大[49]。在灌浆初期,酸性蔗糖转化酶(SAI)是参与蔗糖降解的主要酶,其活性是衡量籽粒库活力的主要指标[50]。本研究中灌浆初期低温处理显著降低了玉米强、弱势粒的SAI活性,减弱了籽粒蔗糖转化能力,影响了籽粒蔗糖代谢,导致灌浆前期灌浆速率及籽粒干重增长缓慢,表现为这两项指标都显著低于对照。在灌浆期协助籽粒将同化物质有效转化为淀粉的是蔗糖合酶(SS)、淀粉合酶(SSS)和ADPG焦磷酸化酶(AGPase)等[22],灌浆期低温降低了以上淀粉积累合成关键酶的活性[51]。本研究发现,籽粒灌浆速率同SSS及AGPase的活性关系密切,呈显著正相关。灌浆前期低温显著降低了SAI及SS的活性,降低了淀粉合成底物的生成及供应能力,同时也显著降低了SS、AGPase的活性,限制了同化产物的转化积累。灌浆后期低温胁迫对SAI、SS、SSS及AGPase活性的负面影响依然存在,降低了淀粉的合成底物的转化和淀粉积累效率,制约了淀粉合成的直接供体ADPG的产生,限制了淀粉链的延长,导致淀粉积累不足,在灌浆后期强势粒SSS和AGPase活性降低程度要高于弱势粒,这可能是导致最终强势粒淀粉含量降低程度大于弱势粒的主要原因之一。
4 结论
低温处理导致玉米强、弱势粒的灌浆速率的降低,是造成籽粒干物质积累减少的重要原因。玉米籽粒受低温影响,在灌浆前期,籽粒生长素(IAA)、玉米素(ZR)、脱落酸(ABA)含量减少,可溶性酸性转化酶(SAI)活性降低,导致库活性不足,库容量减少。在灌浆中期,籽粒淀粉合成中底物的供应及淀粉的合成过程受低温限制,导致籽粒淀粉合成不足,含量降低。低温对玉米强、弱势粒的灌浆过程都造成较大的影响,且对弱势粒的影响大于强势粒。
[1] 马玉平, 王石立, 李维京. 基于作物生长模型的东北玉米冷害监测预测. 作物学报, 2011, 37(10): 1868- 1878.
MA Y P, WANG S L, LI W J. Monitoring and predicting of maize chilling damage based on crop growth model in northeast china., 2011, 37(10): 1868-1878. (in Chinese)
[2] Mariani L, Parisi S G, Cola G. Space and time behavior of climatic hazard of low temperature for single rice crop in the mid latitude., 2009, 29(12): 1862-1871.
[3] 张建平, 王春乙, 赵艳霞, 杨晓光, 王靖. 基于作物模型的低温冷害对我国东北三省玉米产量影响评估. 生态学报, 2012, 32(13): 4132-4138.
ZHANG J P, WANG C Y, ZHAO Y X, YANG X G, WANG J. Impact evaluation of low temperature to yields of maize in northeast China based on crop growth model., 2012, 32(13): 4132-4138. (in Chinese)
[4] 杨若子, 周广胜. 1961-2013年东北三省玉米低温冷害频率的时空动态研究. 气象科学, 2016, 36(3): 311-318.
YANG R Z, ZHOU G S. Temporal-spatial dynamics of maize chilling damage frequency in northeast China during 1961-2013., 2016, 36(3): 311-318. (in Chinese)
[5] 马树庆, 袭祝香, 王琪. 中国东北地区玉米低温冷害风险评估研究. 自然灾害学报, 2003, 12(3): 137-141.
MA S Q, XI Z X, WANG Q. Risk evaluation of cold damage to corn in northeast China., 2003, 12(3): 137-141. (in Chinese)
[6] 王琪, 马树庆, 郭建平, 张铁林, 于海, 徐丽萍. 温度对玉米生长和产量的影响. 生态学杂志, 2009, 28(2): 255-260.
WANG Q, MA S Q, GUO J P, ZHANG T L, YU H, XU L P. Effects of air temperature on maize growth and its yield., 2009, 28(2): 255-260. (in Chinese)
[7] 李祎君, 王春乙. 东北地区玉米低温冷害综合指标研究. 自然灾害学报, 2007, 16(6): 15-20.
LI Y J, WANG C Y. Research on comprehensive index of chilling damage to corn in northeast China., 2007, 16(6): 15-20. (in Chinese)
[8] 李绍长, 白萍, 吕新, 刘淑云, 董树亭. 不同生态区及播期对玉米籽粒灌浆的影响. 作物学报, 2003, 29(5): 775-778.
LI S C, BAI P, LÜ X, LIU S Y, DONG S T. Ecological and sowing date effects on maize grain filling., 2003, 29(5): 775-778. (in Chinese)
[9] 郭志强, 侯立白, 赵明, 张保明, 董志强. 高寒地区低温胁迫条件下化学调控对玉米生理变化的影响. 作物杂志, 2008(2): 42-45.
GUO Z Q, HOU L B, ZHAO M, ZHANG B M, DONG Z Q. Effect of chemical regulation on corn growth and physiological changes under low temperature stress in frost environments., 2008(2): 42-45. (in Chinese)
[10] 王若男, 任伟, 李叶蓓, 张萍, 陈璐洁, 陶洪斌, 王璞. 灌浆期低温对夏玉米光合性能及产量的影响. 中国农业大学学报, 2016, 21(2): 1-8.
WANG R N, REN W, LI Y B, ZHANG P, CHEN L J, TAO H B, WANG P. Effects of low temperature during grain filling stage on photosynthetic characteristics and yield of summer maize., 2016, 21(2): 1-8. (in Chinese)
[11] 祁利潘, 陶洪斌, 周祥利, 吴景玉, 赵丽晓, 王璞. 化学药剂处理对灌浆期低温条件下玉米光合特性及产量的影响. 玉米科学, 2013, 21(3): 52-56.
QI L P, TAO H B, ZHOU X L, WU J Y, ZHAO L X, WANG P. Effects of chemical regulation on maize photosynthesis and yield under low temperature stress during grain filling stage., 2013, 21(3): 52-56. (in Chinese)
[12] 张毅, 顾慰连, 戴俊英. 低温对玉米光合作用超氧物歧化酶活性和籽粒产量的影响. 作物学报, 1992, 18(5): 397-400.
ZHANG Y, GU W L, DAI J Y. Effect of low temperature on photosynthesis activity of superoxide dismutase and kernel yield of maize., 1992, 18(5): 397-400. (in Chinese)
[13] 张建平, 赵艳霞, 王春乙, 杨晓光, 王靖. 不同时段低温冷害对玉米灌浆和产量的影响模拟. 西北农林科技大学学报(自然科学版), 2012, 40(9): 115-121.
ZHANG J P, ZHAO Y X, WANG C Y, YANG X G, WANG J. Modeling the impact of low temperature disaster during different development stages on grain filling processes and yields of maize., 2012, 40(9): 115-121. (in Chinese)
[14] 张凤路, 孙国伟, 赵国顺, 江亚丽. 不同温度处理对离体培养玉米子粒发育的影响研究. 玉米科学, 2006, 14(6): 83-85.
ZHANG F L, SUN G W, ZHAO G S, JIANG Y L. Studies on the effects of temperature on maize kernel development in vitro., 2006, 14(6): 83-85. (in Chinese)
[15] 于涛, 李耕, 刘鹏, 董树亭, 张吉旺, 赵斌, 柏晗. 玉米早期发育阶段粒位效应的蛋白质组学分析. 中国农业科学, 2016, 49(1): 54-68.
YU T, LI G, LIU P, DONG S T, ZHANG J W, ZHAO B, BAI H. Proteomics analysis of grain position effects during early developmental stages of maize., 2016, 49(1): 54-68. (in Chinese)
[16] 曾研华, 张玉屏, 潘晓华, 朱德峰, 向镜, 陈惠哲, 张义凯. 花后低温对水稻籽粒灌浆与内源激素含量的影响. 作物学报, 2016, 42(13) : 4132-4138.
ZENG Y H, ZHANG Y P, PAN X H, ZHU D F, XIANG J, CHEN H Z, ZHANG K Y. Effect of low temperature after flowering on grain filling and plant hormones contents in rice., 2016, 42(13): 4132-4138. (in Chinese)
[17] Nayyar H, Kaur G, Kumar S, Upadhyaya H D. Low temperature effects during seed filling on chickpea genotypes (L.): probing mechanisms affecting seed reserves and yield.2007, 193(5): 336-344.
[18] ZHANG W Y, CAO Z Q, ZHOU Q, CHEN G, GU J F, LIU L J, WANG Z Q, YANG J C, ZHANG H. Grain filling characteristics and their relations with endogenous hormones in large- and small-grain mutants of rice.2016, 11(10): e0165321.
[19] 戴俊英, 苏正淑, 张毅. 灌浆期低温对玉米籽粒的伤害作用. 作物学报, 1995, 21(1): 71-75.
DAI J Y, SU Z S, ZHANG Y. Chilling injury of kernel of maize plant stressed at filling stage., 1995, 21(1): 71-75. (in Chinese)
[20] Farooq M, Aziz T, Wahid A, Lee D J, Siddique K H M. Chilling tolerance in maize: agronomic and physiological approaches.2009, 60(6): 501-516.
[21] 张海艳, 董树亭, 高荣岐, 李玉全. 玉米籽粒淀粉积累及相关酶活性分析. 中国农业科学, 2008, 41(7): 2174-2181.
ZHANG H Y, DONG S T, GAO R Q, LI Y Q. Starch accumulation and enzyme activities associated with starch synthesis in maize kernels., 2008, 41(7): 2174-2181. (in Chinese)
[22] ZHANG H Y, DONG S T, GAO R Q, Li Y Q. Comparison of starch synthesis and related enzyme activities in developing grains among different types of maize.y, 2007, 33(1): 25-32.
[23] 王志刚, 梁红伟, 余少波, 苏治军, 孙继颖, 于晓芳, 高聚林. 玉米弱势粒库特征及其调控机理研究进展. 作物杂志, 2015(2): 7-11.
WANG Z G, LIANG H W, YU S B, SU Z J, SUN J Y, YU X F, GAO J L. Review on sink traits and regulation mechanism of inferior kernels of maize., 2015(2): 7-11. (in Chinese)
[24] 赵丽晓, 张萍, 王若男, 王璞, 陶洪斌. 花后前期高温对玉米强弱势籽粒生长发育的影响. 作物学报, 2014, 40(10): 1839-1845.
ZHAO L X, ZHANG P, WANG R N, WANG P, TAO H B. Effect of high temperature after flowering on growth and development of superior and inferior maize kernels., 2014, 40(10): 1839-1845. (in Chinese)
[25] 陈素省, 张凤路, 郝艳敏, 赵国顺, 江亚丽, 王兆晓. 离体条件下玉米籽粒发育动态研究. 华北农学报, 2006, 21(S1): 28-31.
CHEN S X, ZHANG F L, HAO Y M, ZHAO G S, JIANG Y L, WANG Z X. Studies on the development of maize kernel culture., 2006, 21(S1): 28-31. (in Chinese)
[26] Gengenbach B G. Development of maize caryopses resulting from in-vitro pollination., 1977, 134(1): 91-93.
[27] 宋庆芳, 赵丽晓, 吴景玉, 王璞, 陶洪斌. 玉米子粒离体培养条件下适宜子粒穗轴比的筛选. 玉米科学, 2012, 20(3): 48-50.
SONG Q F, ZHAO L X, WU J Y, WANG P, TAO H B. Improvement of the technology of maize kernels cultured.s, 2012, 20(3): 48-50. (in Chinese)
[28] Cully D E, Gengenbach B G, Smith J A,Rubenstein I, Connelly J A, Park W D. Endosperm protein synthesis and l-[S] methionine incorporation in maize kernels cultured., 1984, 74(2): 389-394.
[29] 乔嘉, 朱金城, 赵姣, 郑志芳, 王璞, 廖树华. 基于Logistic模型的玉米干物质积累过程对产量影响研究. 中国农业大学学报, 2011, 16(5): 32-38.
QIAO J, ZHU J C, ZHAO J, ZHENG Z F, WANG P, LIAO S H. Study on the effect of dry matter accumulation process on maize yield based on logistic model., 2011, 16(5): 32-38. (in Chinese)
[30] 何照范. 粮油籽粒品质及其分析技术. 北京: 中国农业出版社, 1985.
HE Z F.. Beijing: Agriculture press, 1985. (in Chinese)
[31] 宋松泉, 程红焱, 龙春林, 姜孝成. 种子生物学研究指南. 北京: 科学出版社, 2005.
SONG S Q, CHENG H Y, LONG C L, JIANG X C.. Beijing: Science Press, 2005. (in Chinese)
[32] Yang J, Zhang J, Wang W. Hormonal changes in the grains of rice subjected to water stress during grain filling., 2001, 127(1): 315-323.
[33] Nakamura Y, Yuki K, Park S Y,Ohya T. Carbohydrate metabolism in the developing endosperm of rice grains., 1989, 30(6): 833-839.
[34] Zinselmeier C, Westgate M E, Schussler J R, Jones R J. Low water potential disrupts carbohydrate metabolism in maize (L.) ovaries., 1995, 107(2): 385-391.
[35] 黄振喜, 王永军, 王空军, 李登海, 赵明, 柳京国, 董树亭, 王洪军, 王军海, 杨今胜. 产量15 000 kg·ha-1以上夏玉米灌浆期间的光合特性. 中国农业科学, 2007, 40(9): 1898-1906.
HUANG Z X, WANG Y J, WANG K J, LI D H, ZHAO M, LIU J G, DONG S T, WANG H J, WANG J H, YANG J S. Photosynthetic characteristics during grain filling stage of summer maize hybrids with high yield potential of 15, 000 kg/ha., 2007, 40(9): 1898-1906. (in Chinese)
[36] 郭庆法, 王庆成, 汪黎明. 中国玉米栽培学. 上海: 上海科学技术出版社, 2004.
GUO Q F, WANG Q C, WANG L M.. Shanghai: Shanghai Scientific and Technical Publishers, 2004. (in Chinese)
[37] 马玉平, 王石立, 李维京. 基于作物生长模型的玉米生殖期冷害致灾因子研究. 作物学报, 2011, 37(9): 1642-1649.
MA Y P, WANG S L, LI W J. Chilling disaster factors in maize reproductive stage based on crop growth model., 2011, 37(9): 1642-1649. (in Chinese)
[38] 张建平, 赵艳霞, 王春乙, 杨晓光, 王靖. 不同发育期低温冷害对玉米灌浆和产量影响模拟. 中国农学通报, 2012, 28(36): 176-182.
ZHANG J P, ZHAO Y X, WANG C Y, YANG X G, WANG J. The simulation of the effects of low temperature disasters on maize growth and yield during different development stages., 2012, 28(36): 176-182. (in Chinese)
[39] 王柳, 熊伟, 温小乐, 冯灵芝. 温度降水等气候因子变化对中国玉米产量的影响. 农业工程学报, 2014, 30(21): 138-146.
WANG L, XIONG W, WEN X L, FENG L Z. Effect of climatic factors such as temperature, precipitation on maize production in China., 2014, 30(21): 138-146. (in Chinese)
[40] Ahmed N, Maekawa M, Tetlow I J. Effects of low temperature on grain filling, amylose content, and activity of starch biosynthesis enzymes in endosperm of basmati rice., 2008, 59(7): 599-604.
[41] 陈振林, 张建平, 王春乙, 郑江平. 应用WOFOST模型模拟低温与干旱对玉米产量的综合影响. 中国农业气象, 2007, 28(4): 440-442.
CHEN Z L, ZHANG J P, WANG C Y, ZHENG J P. Application of WOFOST model in simulation of integrated impacts of low temperature and drought on maize yield., 2007, 28(4): 440-442. (in Chinese)
[42] 张巽, 张萍, 陈璐洁, 陶洪斌, 王璞. 灌浆期低温对离体玉米子粒灌浆过程及内源激素的影响. 玉米科学, 2017, 25(2): 49-54.
ZHANG X, ZHANG P, CHEN L J, TAO H B, WANG P. Effect of grain filling and endogenous hormones changes on maize grains in vitro culture under low temperature stress during grain filling stage.2017, 25(2): 49-54. (in Chinese)
[43] BRENNER M L, CHEIKH N. The role of hormones in photosynthate partitioning and seed filling//Dabies P J.. Dordrecht: Springer, 1995: 649-670.
[44] 徐云姬, 顾道健, 张博博, 张耗, 王志琴, 杨建昌. 玉米果穗不同部位籽粒激素含量及其与胚乳发育和籽粒灌浆的关系. 作物学报, 2013, 39(8): 1452-1461.
XU Y J, GU D J, ZHANG B B, ZHANG H, WANG Z Q, YANG J C. Hormone contents in kernels at different positions on an ear and their relationship with endosperm development and kernel filling in maize., 2013, 39(8): 1452-1461. (in Chinese)
[45] 杨建昌, 仇明, 王志琴, 刘立军, 朱庆森. 水稻发育胚乳中细胞增殖与细胞分裂素含量的关系. 作物学报, 2004, 30(1): 11-17.
YANG J C, QIU M, WANG Z Q, LIU L J, ZU Q S. Relationship between the cell proliferation and cytokinin contents in rice endosperm during its development., 2004, 30(1): 11-17. (in Chinese)
[46] 徐田军, 吕天放, 赵久然, 王荣焕, 陈传永, 刘月娥, 刘秀芝, 王元东. 玉米籽粒灌浆特性对播期的响应. 应用生态学报, 2016, 27(8): 2513-2519.
XU T J, LÜ T F, ZHAO J R, WANG R H, CHEN C Y, LIU Y E, LIU X Z, WANG Y D. Response of grain filling characteristics of maize to sowing date.2016, 27(8): 2513-2519. (in Chinese)
[47] 夏楠, 赵宏伟, 吕艳超, 赵振东, 邹德堂, 刘化龙, 王敬国, 贾琰. 灌浆结实期冷水胁迫对寒地粳稻籽粒淀粉积累及相关酶活性的影响. 中国水稻科学, 2016, 30(1): 62-74.
XIA N, ZHAO H W, LÜ Y C, ZHAO Z D, ZOU D T, LIU H L, WANG J G, JIA Y. Effect of cold-water stress at grain-filling stage on starch accumulation and related enzyme activities in grains ofrice in cold-region., 2016, 30(1): 62-74. (in Chinese)
[48] LIANG J, ZHANG J, CAO X. Grain sink strength may be related to the poor grain filling of indica-japonica rice () hybrids., 2010, 112(4): 470-477.
[49] 梁红伟, 高聚林, 王志刚, 于晓芳, 孙继颖, 胡树平, 王振, 余少波, 李亚剑. 基于控制授粉技术的玉米弱势粒发育与库特征的关系. 作物学报, 2016, 42(6): 917-923.
LIANG H W, GAO J L, WANG Z G, YU X F, SUN J Y, HU S P, WANG Z, YU S B, LI Y J. Relationship of inferior kernel setting with sink strength under controlling pollination., 2016, 42(6): 917-923. (in Chinese)
[50] 张凤路, 赵久然, 王纪华. 离体条件下玉米籽粒发育调控研究. 杂粮作物, 2001, 21(4): 34-35.
ZHANG F L, ZHAO J R, WANG J H. Regulating study on maize kernel development in vitro., 2001, 21(4): 34-35. (in Chinese)
[51] 戴忠民, 王振林, 高凤菊, 李文阳, 闫素辉, 蔡瑞国, 张敏, 尹燕枰. 两种供水条件下两穗型小麦品种籽粒淀粉积累及相关酶活性的变化特征. 作物学报, 2007, 33(4): 682-685.
DAI Z M, WANG Z L, GAO F J, LI W Y, YAN S H, CAI R G, ZHANG M, YIN Y P. Characterization of starch accumulation and activities of enzymes involved in starch synthesis in grains of wheat cultivars differing in spike types field-grown in irrigation and rainfed conditions., 2007, 33(4): 682-685. (in Chinese)
(责任编辑 杨鑫浩)
Effects of Low Temperature on Maize Superior and Inferior Kernels Development during Grain Filling in vitro
ZHANG Xun1,2, HAO JianPing1, WANG Pu3, ZHANG Ping3, CHEN LuJie3
(1College of Agriculture, Shanxi Agricultural University, Taigu 030801, Shanxi;2School of Life Sciences, Datong University, Datong 037009, Shanxi;3College of Agronomy and Biotechnology, China Agricultural University, Beijing 100193)
【Objective】In this study, the physiological mechanism of kernel weight reduction under low temperature had been uncovered through grain filling dynamic, carbon metabolism, and phytohormone balance, and reveal the effect of low temperature on maize superior and inferior kernels during grain filling, which could provide a theoretical guidance for crop production. 【Method】 Hybrid maize Zhengdan 958 was chosen for the experimental material, which was grown in the filed condition. Kernel was collected at 3 days after pollination, and was divided into superior and inferior kernel according to its position at the ear. Afterward, kernels were separately incubated with solid medium under normal (25°C) and low (16°C) temperature in artificial climate incubators. Superior and inferior kernels were sampled with 10-day interval since pollination for analyzing dry weight dynamic, hormone contend, starch accumulation and its related enzymes. 【Result】Compared with CK, Final kernel weight decreased by 47.58% for superior kernel and 50.95% for inferior kernels under low temperature, respectively. A longer active grain-filling period had been observed, caused by slower grain filling rate increase during lag period and slower decrease during maturation drying period. But, the grain-fill rate during active grain-filling period decreased 55.39% for superior kernel and 54.72% for inferior kernel, respectively. The increase of grain-filling rate in lag periods and the decrease of grain-filling rate in maturation drying periods of maize kernels were reduced significantly, and the time of active grain-filling periods was prolonged by 5-7 days under low temperature stress. The content of IAA, ZR and ABA decreased significantly in both superior and inferior kernels after 10 days low temperature incubation, while it was opposite for GA3. Moreover, decreased content of IAA and ZR in inferior kernels and increased ABA in superior kernels had been found after 30 days low temperature treatment. Low temperature decreased activity of soluble acid invertase (SAI), sucrose synthase (SS), soluble starch synthase (SSS) and ADP-glucose pyrophosphorylase (APGase) in both superior and inferior kernel during early grain filling periods, resulting in decreased starch accumulation in kernel. Furthermore, the decrease of SAI in inferior kernels was greater than that of superior kernels, while it was opposite for other enzymes.【Conclusion】The disturbed hormone balance during earlier grain filling period greatly weakened the sink capacity under low temperature stress, while the decreased activity of enzyme during mid-grain filling period limited the starch accumulation. The grain filling rate decreased in both superior and inferior kernels, resulting in kernel weight reduction. The grain filling process had been affected in both superior and inferior kernels under low temperature stress. Furthermore, the inferior kernels was more sensitive to low temperature than superior kernels.
maize; low temperature; vitro culture; grain filling stage; phytohormone
2017-09-15;
2018-05-02
国家玉米产业技术体系建设专项(CARS-02-26)、山西省大同市基础科技项目(2015105)
张巽,E-mail:sxdtuzx@163.com。
郝建平,E-mail:tghjp88@sina.com。通信作者王璞,E-mail:wangpu@cau.edu.cn
10.3864/j.issn.0578-1752.2018.12.004猜你喜欢
现代畜牧科技(2021年4期)2021-12-05 15:34:45
现代畜牧科技(2021年10期)2021-11-19 08:42:26
今日农业(2021年12期)2021-10-14 07:31:14
疯狂英语·初中天地(2020年8期)2020-09-11 07:43:38
中国化肥信息(2020年7期)2020-03-19 01:54:06
中国化肥信息(2019年7期)2019-08-26 09:46:54
河北农业科学(2018年2期)2018-07-26 11:28:14
江西建材(2018年4期)2018-04-10 12:37:06
水利规划与设计(2017年6期)2017-07-18 10:56:27
商洛学院学报(2017年2期)2017-05-17 05:19:53
