
基于Ecopath模型的长江口生态系统营养结构和能量流动研究
2018-06-29 02:54王思凯
海洋渔业 2018年3期
徐 超, 王思凯, 赵 峰, 宋 超, 庄 平
(1. 南京农业大学无锡渔业学院, 江苏无锡 214081;2. 中国水产科学研究院东海水产研究所,农业部东海与长江口渔业资源环境科学观测实验站, 上海 200090)
河口是海洋生态系统的重要组成部分,以其独特的地理位置、水动力条件和物质基础,为众多海洋生物的生存和繁衍提供了有利的生存条件[1]。长江口作为我国最大的河口,位于暖温性的黄海生态系统与暖水性的东海生态系统的交界处,受长江径流及黄海冷水团相互消长的影响,环境因素变化剧烈,生态环境错综复杂。源源不断的长江径流带来了大量营养物质,维持了长江口生态系统的巨大生产力,孕育了丰富的饵料资源,使长江口成为许多重要经济鱼类的产卵场、索饵场、育幼场和洄游通道[2]。近年来对长江口的研究主要集中在对水生生物群落结构[3-4]、水生动物食性[5-6]及水环境质量评价[7-8]等方面。然而,随着捕捞强度的增大、环境污染和渔场海洋环境条件的变化,长江口生物多样性和生态系统环境质量受到前所未有的胁迫和影响,因而对长江口生态系统结构和功能的研究就显得十分必要。
Ecopath with Ecosim (EwE)是一组物质平衡模型软件[9]。它以食物网为主线,根据营养动力学原理,用一组联立方程平衡系统中各功能组生物生产和消费的能量流动,直接构造以物质平衡为基础的生态系统模型。这种方法最早由POLOVINA提出,用来评估稳定状态水域生态系统组成的生物量和食物消耗,经过与ULANOWICZ的能量分析生态学理论结合,逐步发展成为一种生态系统营养成分流动分析方法[10]。该方法可用于研究:生态系统的规模、稳定性和成熟度;物流、能流的分布和循环;系统内部的捕食和层级关系;各层级间能量流动的效率;中间生态位的竞争及彼此互利或危害的程度等,被誉为新一代水域生态系统研究的核心工具[11]。目前,Ecopath模型在水生生态系统中得到了广泛应用,国内外学者利用Ecopath模型描述了渤海[12]、南黄海[13]、东海[14]和南海[15]等不同中国近海海域生态系统的能流过程,而对河口水域的研究相对较少。因此,本文通过利用Ecopath with Ecosim (版本6.4.3)软件构建2016-2017年长江口及邻近水域生态系统的能流网络模式,并对此历史时期生态系统结构与功能进行综合评估,旨在通过对该河口生态系统的结构、能量流动特征的定量分析,为我国长江口生态学研究积累新的资料,并为长江口渔业资源的利用和环境管理提供科学的理论依据。
1 材料与方法
1.1 研究区域
分别于2016年8月(夏季)、11月(秋季)、2017年2月(冬季)、5月(春季)在长江口水域(31°00′00″~31°48′00″N、121°00′00″~122°48′00″E)设置的22个站点进行4次拖网调查(图1)。调查内容包括浮游植物、浮游动物、渔业生物、底栖动物等。渔业生物(主要包括鱼类、虾类、蟹类和头足类)使用的调查船功率为45 kW,单船桁杆拖网为桁杆长6 m的双囊网,网具总长15 m,高2 m,网囊孔径2 cm,每个位点拖网时间0.5 h,平均航速3 kn;分别在涨潮和落潮时使用Ⅰ型浮游动物网和Ⅱ型浮游植物网采集浮游生物;底栖生物样品则是通过各站位用采泥器取两次做一个样品,经0.5 mm筛网冲洗获取。

图1 长江口采样地点示意图Fig.1 Location of sampling sites in the Yangtze Estuary
1.2 数据来源
建立Ecopath模型需要真实可靠的数据,收集的数据应尽可能的完整,覆盖生态系统中的所有生物。模型需要输入的参数包括生物量(B)、生产量与生物量比值(P/B)、消费量与生物量比值(Q/B)和生态营养效率(EE)这4个参数,需要输入其中的3个,另外一个参数(通常为EE)由模型估计得出;同时食物组成矩阵(DC)和捕捞量也必须输入。
本研究的数据来源于2016年8月-2017年5月在长江口及邻近水域调查所得,对水生生物作了详细的生物学测定和胃含物分析。其中,鱼类、虾蟹类和头足类的生物量数据通过采样调查获得,转化公式如下:
M=G/[S(1-E)]
(1)
式(1)中,M为生物量,G为调查生物量,S为网口的扫海面积,E为逃逸率。
隐埋性底栖动物与浮游生物的生物量分别靠底泥采样和浮游生物垂直拖网样品估计,有机碎屑生物量用有机碎屑与初级生产力的经验公式估算[16],食物组成矩阵通过详细的胃含物食性分析法所得,捕捞量通过《中国渔业统计年鉴》中的资料估算,对于难以估计的参数(P/B与Q/B),则主要通过国际水生资源管理中心网站(www.fishbase.org)和历史资料数据所得[16-18],生态营养效率(EE)作为未知参数,通过模型估计得出。
1.3 研究方法
1.3.1 Ecopath模型基本原理
Ecopath模型定义生态系统是由一系列生态关联的功能组组成,并遵循热力学定律研究生态系统能量流动与网络分析。功能群应涵盖生态系统能流的全过程[11]。该模型假定在给定一段时间内(通常为1a)系统能量守恒。它包含两个核心方程,一个方程描述各功能群生产量的流向[下式(2)],一个方程用来描述各功能群的能量守恒[下式(3)]。
Pi=Yi+M2iBi+Ei+BAi+M0iBi
(2)
Qi=Pi+Ri+Ui
(3)
式(2)、式(3)中,Pi表示生产量,Yi为渔获量,M2i为捕食死亡率,Bi为生物量,Ei为净迁出量,BAi为生物量累积,M0i为其它死亡率, 即功能群未被生态系统所利用的那部分能量所占的比重;Qi表示第i功能群对其它功能群的消费量,Pi表示生产量,Ri表示呼吸量,Ui表示未同化量。
1.3.2 功能组的划分
按照研究需要,将生态学或分类地位上相似的物种加以整合,也可以把单个物种或者单个物种的某个年龄阶段(成体或幼体)作为独立的功能组[19],其中功能组中必须包含1个或数个碎屑功能组[20]。本研究主要根据不同物种生态学与生物学特性(食性、栖息地、死亡率等)的相似性以及在渔业方面的重要程度来划分功能组,共14个功能组(表1)。
1.3.3 Ecopath模型的调试
对于模型的建立,首先要考虑的是生态营养效率EE值应介于0~1之间,其次要考虑总效率GE值,它被定义为生产量与消耗量(P/Q)之比。在许多情况下GE值应介于0.1~0.3之间[21]。在食性数据、捕捞数据、基础数据3大类数据输入模型后,模型并未成为平衡状态,仍有生态转移效率大于1的功能组,长江口生态通道模型调试主要从食物组成着手[22],对各功能组的食物构成做出调整,以达到模型平衡状态。
2 结果与分析
2.1 长江口生态系统的营养级
由表2可见,2016-2017年长江口及邻近海域生态系统各功能群的营养级集中在1~3.93。其中,浮游植物与碎屑营养级为1;浮游动物营养级为2.18;隐埋性底栖生物、头足类及虾、蟹类的营养级集中在2.47~3.11;鱼类营养级分布较广,除中上、下层浮游生物碎屑食性鱼类的营养级较低位于2.41~2.47,其它鱼类功能群营养级为3.49~3.93。

表1 长江口2016-2017年生态系统模型的功能组Tab.1 Functional groups of ecosystem model in 2016-2017 in the Yangtze Estuary

表2 长江口生态系统Ecopath模型功能组估算参数Tab.2 In and out parameters of the Ecopath of the Yangtze Estuary ecosystem
注:加粗字体为模型计算的参数
Note: The bold font is the parameter of the model calculation
2.2 长江口生态系统营养级间的物质流动
Lindeman模式可以将复杂的食物网简化成线性的食物链。通过营养级分解[23],将各功能组的能流重组,形成生态系统各营养级的能流分布。2016-2017年长江口生态系统能流分布主要集中在营养级Ⅰ~Ⅲ中(图2),当营养级为Ⅳ甚至更高时能流可以忽略不计。其中来自营养级Ⅰ级的能流在系统总能流中所占比例最高,为53.67%;其次为第Ⅱ营养级所占比例为45.19%。整个营养级Ⅰ流入到营养级Ⅱ的物流通量为1 637 t·km-2·a-1;从营养级Ⅱ到营养级Ⅴ的总流量随其营养级的增加而依次降低,分别为36.52 t·km-2·a-1、6.463 t·km-2·a-1、0.967 t·km-2·a-1。此外,营养级Ⅱ、Ⅲ、Ⅳ的能流转换效率依次分别为2.31%、19.30%和17.80%,平均传输效率为13.14%。
2.3 长江口生态系统营养级间的能量转化效率
营养级间的能流转化效率是指该营养级输出和被摄食的流量与总流量之间的比值,体现了该营养级在生态系统中的被利用的效率[24]。通过比较不同营养级间的能量转换效率(表3)可以看出,长江口初级生产者到第Ⅱ营养级的转换效率为2.2%,而碎屑到第Ⅱ营养级的转换效率为4.3%,生产者和碎屑总能量转换效率为2.3%;第Ⅱ营养级到第Ⅲ营养级转换效率很高,综合转换效率为19.3%;第Ⅲ营养级到第Ⅳ营养级综合转换效率为17.8%;第Ⅳ营养级到第Ⅴ营养级综合转化效率为9.1%。可见,低营养级间的能流转化效率相对较低,而高营养级间的能流转化效率相对较高。来自初级生产者和碎屑的能量转换效率分别为9.1%和10.6%,且长江口生态系统的总体能量转换效率为9.3%。根据生态金字塔能量转换的“十分之一定律”[25],该河口能量的利用效率未达到最适程度。
2.4 长江口生态系统食物网结构
从图3可以看出,长江口生态系统食物网结构较为复杂,营养流动主要有两条途径:一是从浮游植物开始的牧食食物链,如:浮游植物—浮游动物—中上层浮游生物碎屑食性鱼类—中上层混合食性鱼类;二是从碎屑开始的碎屑食物链,如碎屑—隐埋性底栖生物—虾类—底层游泳生物食性鱼类。表4中联结指数(CI)和系统杂食指数(SOI)分别为0.345、0.321,说明长江口食物链普遍较短。因此,一个物种数量的变动或者消失都可能影响到整个长江口生态系统的食物网结构。

图2 长江口生态系统各营养级间的物质流动Fig.2 Trophic flows transmitted through aggregated trophic levels in the Yangtze Estuary注:P表示初级生产者浮游植物;D表示有机碎屑Note: P denotes primary producer phytoplankton; D denotes organic detritus
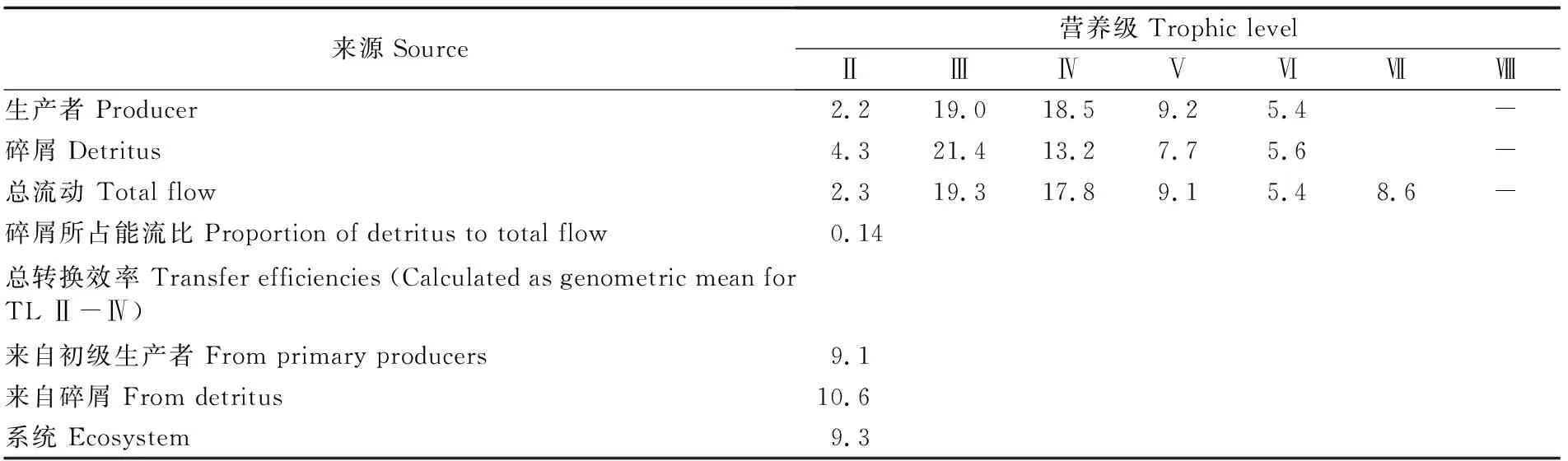
来源Source营养级Trophic levelⅡⅢⅣⅤⅥⅦⅧ生产者 Producer2.219.018.59.25.4-碎屑 Detritus4.321.413.27.75.6-总流动 Total flow2.319.317.89.15.48.6-碎屑所占能流比 Proportion of detritus to total flow0.14总转换效率 Transfer efficiencies (Calculated as genometric mean for TL Ⅱ-Ⅳ)来自初级生产者 From primary producers9.1来自碎屑 From detritus10.6系统 Ecosystem9.3
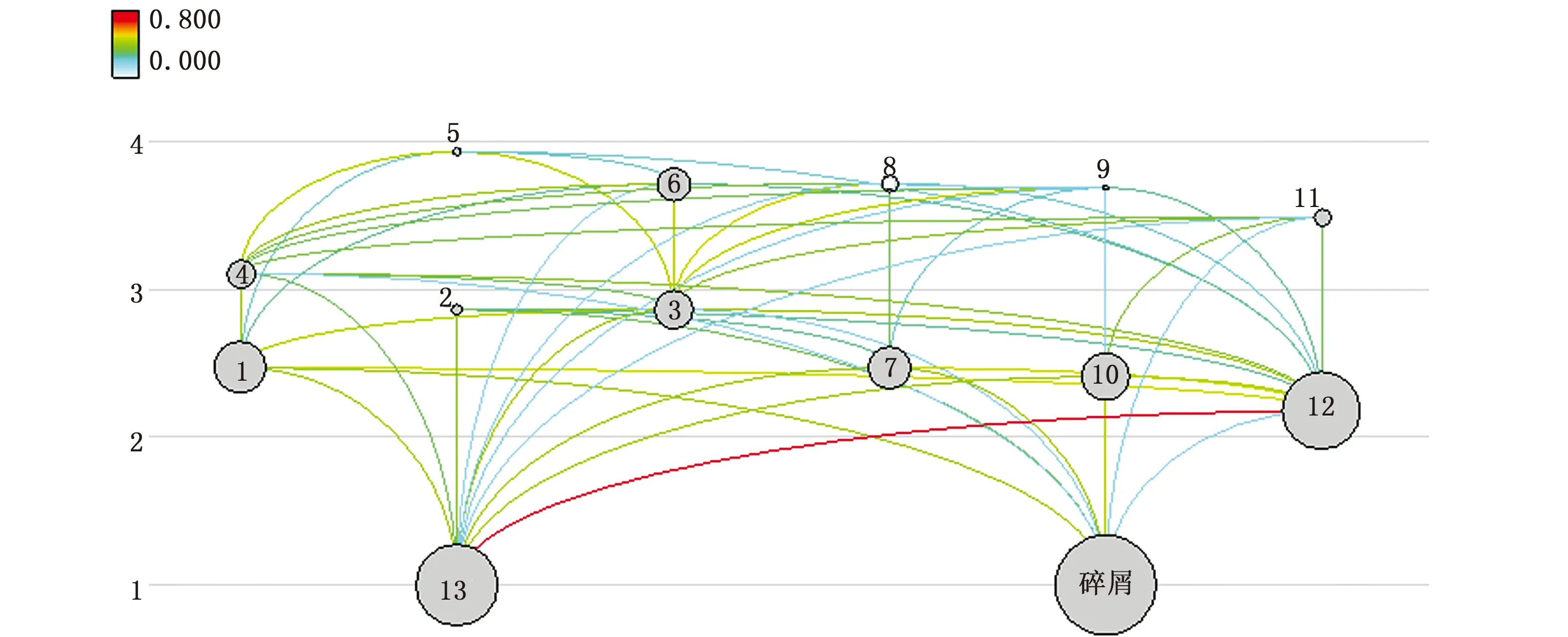
图3 长江口生态系统的食物网Fig.3 Food web of the Yangtze Estuary ecosystem注:1.隐埋性底栖生物; 2.头足类; 3.虾类; 4.蟹类; 5.底层游泳生物食性鱼类; 6.底层混合食性鱼类; 7.中下层浮游生物碎屑食性鱼类;8.中下层游泳生物食性鱼类; 9.中下层混合食性鱼类; 10.中上层浮游生物碎屑食性鱼类; 11.中上层混合食性鱼类; 12.浮游动物; 13.浮游植物Note: 1.Benthic producers; 2.Cephalopods; 3.Shrimps; 4.Crabs; 5.Benthic predators; 6.Benthic mixed predators; 7.Middle-lower plankton detritus predators; 8.Middle-lower predators; 9.Middle-lower mixed predators; 10.Middle-upper plankton detritus predators; 11.Middle-upper mixed predators; 12. Zooplankton; 13.Phytoplankton
2.5 长江口生态系统发育成熟度
生态系统的很多特征均可用于表征生态系统的成熟度[26]。生态系统发育早期,通常系统的净初级生产量Pp较高,而系统的呼吸R消耗较小,因此Pp/R>1;随着生态系统的发育不断趋于成熟,其生物量不断累积,呼吸消耗逐步增大,则Pp/R接近1[27]。通过软件自动计算(表4)可见,2016-2017年长江口的Pp/R值为1.245大于1,表明该河口生态系统趋于不成熟。此外,从生态系统发育的信息学特征来研究生态系统的发育成熟度,联结指数(CI)和系统杂食指数(SOI)是表征系统内部联系复杂程度的指标,越是成熟的生态系统,CI和SOI的数值越接近于1,其各功能组间的联系越强,系统越趋于稳定[28]。长江口生态系统的CI和SOI分别为0.345和0.321,表明该生态系统的初级生产力较高,但各功能组的聚合度较低,联结程度较为松散。

表4 2016-2017长江口生态系统的总体特征Tab.4 General properties of the Yangtze Estuary ecosystem in 2016-2017
3 讨论
本文通过Ecopath模型分析了长江口2016-2017年生态系统营养结构和能量流动,结果表明:长江口尚处于不成熟的发展阶段,其生态系统稳定性较低,生物资源种类组成较为简单,系统的初级生产力/呼吸量比值大于1,说明系统中仍有较多的剩余能量未消耗。
本文计算生态系统功能组的营养级集中在1~3.93,这与张效嘉等[16]构建的长江口1985-1986年Ecopath模型所计算的营养级集中在1~4.52相差较大;此外,罗秉征等[29]运用胃含物分析法研究了长江口20世纪90年代鱼类的营养级大小,集中在3.1~4.5。通过对比发现,长江口水生生物的营养级逐渐降低,而水生生物平均营养级的数值大小反映了渔业资源开发利用程度。随着捕捞强度的增大,长江口水生生物资源量处于退化状态。李建生等[30]通过对长江口2003年渔场渔业资源的调查,并与2000-2002年同期比较,发现渔获物资源量以及种类数都有不同程度的降低。目前以底拖网为主的捕捞渔业使底层的大型经济渔业资源迅速减少,渔获物间的捕食关系发生改变,食物网组成结构也由以往肉食性、长寿的底层鱼类向浮游生物食性、短寿的上层鱼类转变[3];此外,长江口及其邻近海域由于长期受到环境污染和渔场海洋环境条件的变化,如三峡大坝的成功截流、洋山港深水码头以及长江口深水航道治理工程的建设等因素,水生生物群落结构发生显著变化[31]。RYDER等[32]指出,过度捕捞和环境退化使生物群落的生态系统失去恢复力和完整性,生态系统的稳定性变差,使依赖生态系统产出的渔业产量在质和量两个方面具有不可预见的变化。因而长江口水生生物的营养级降低很大程度上应该是环境因素和捕捞因素的共同作用结果[33];此外,本研究选取的参数如P/B、Q/B,以及研究水域与张效嘉等[16]略有差异,因而计算出的营养级不同。
长江口生态系统能量流动和转换通过牧食食物链和碎屑食物链共同实现,且主要发生在5个营养级间。其中,直接来自碎屑仅占总能流的14%,来自初级生产者的占总能流的86%。这与张效嘉等[16]研究的1985-1986年长江口生态系统能量流动存在差异,其研究结果表明,来自碎屑的占总能流的43%,来自初级生产者的占总能流的57%。此外,韩瑞等[22]研究发现,三峡工程蓄水前后碎屑食物链在系统能量流动占比由44%下降到41%。三峡工程蓄水后,长江入海径流量减少,长江冲淡水面积减少,陆源营养物质和悬浮颗粒的输入减少,使得生态系统能量传递更依赖牧食食物链[22]。此外,长江口2016-2017年生态系统中生物量、生产量和流量基本上均呈金字塔型分布,即底层营养级的生物量和流量较大,越到顶级越小,基本上符合能量和生物量金字塔规律。
在生态系统演替过程中,群落结构和功能的同时变化是有规律的,在“幼态”时期,生物多表现为r-对策。随着发育的进程,k-对策生物的比例开始增加[27]。在功能方面,随着演替的不断进行,初级生产量与呼吸量的比值从大于1逐渐发展到接近于1。而群落净生产量从高到低,能流渠道从线性到网状,碎屑食物链的重要性逐渐增加[34]。目前,长江口生态系统在人类活动和自然环境因素的影响下发生了逆行演替的现象,系统特征从较为“成熟”的状态向“幼态”转变。2016-2017年长江口生态系统的CI、SOI值分别为0.345、0.321;表明长江口生态系统的成熟度和稳定性以及系统抵抗外力干扰的能力较低。在自然进化过程中,生态系统的演替是一个较为缓慢的过程,但是自然环境变动和人类活动可以加速演替的进程[27]。因此,如何可持续开发与利用长江口渔业资源,维护其生态系统的健康和稳定,合理控制其生态演替的速度,是目前急需解决的问题。从长江口生态系统的结构、物流和能流的特征来看,应依据生态学原理,包括生态演替、食物链网结构、物质循环和转化、生物间的交互营养关系等原则,从不断完善生态系统的结构组成、促进功能的发展成熟角度出发,建立可持续发展的生态系统,从而保护长江口生态环境、实现人与自然的和谐发展。
参考文献:
[1] KIMMERER W J. Physical, biological, and management responses to variable Freshwater flow into the San Francisco estuary [J]. Estuary, 2002, 25(6B): 1275-1290.
[2] 罗秉征, 沈焕庭. 三峡工程与河口生态环境[M]. 北京: 科学出版社, 1994.
LUO B Z, SHEN H T. Impacts of the Three Gorges Project on estuarine ecology and environment [M]. Beijing: Science Press, 1994.
[3] 金显仕, 单秀娟, 郭学武, 等. 长江口及其邻近海域渔业生物的群落结构特征[J]. 生态学报, 2009, 29(9): 4761-4772.
JIN X S, SHAN X J, GUO X W,etal. Community structure of fishery biology in the Yangtze River estuary and its adjacent waters [J]. Acta Ecological Sinica, 2009, 29(9): 4761-4772.
[4] 吴 强, 李显森, 王 俊, 等. 长江口及邻近海域无脊椎动物群落及其生物多样性研究[J]. 水生态学杂志, 2009, 2(2): 73-79.
WU Q, LI X S, WANG J,etal. Community structure and biodiversity of invertebrate resources in the Yangtze River Estuary and its adjacent waters [J]. Journal of Hydroecology, 2009, 2(2): 73-79.
[5] 罗 刚, 庄 平, 章龙珍, 等. 长江口中华鲟幼鱼的食性组成及摄食习性[J]. 应用生态学报, 2008, 19(1): 144-150.
LUO G, ZHUANG P, ZHANG L Z,etal. Diet composition and feeding habits of juvenile Acipenser sinensis in Yangtze River estuary [J]. Chinese Journal of Applied Ecology, 2008, 19(1): 144-150.
[6] 洪巧巧, 庄 平, 杨 刚, 等. 长江口中国花鲈食性分析[J]. 生态学报, 2012, 32(13): 4181-4190.
HONG Q Q, ZHUANG P, YANG G,etal. Feeding habits of Lateolabrax maculates in Yangtze River estuary [J]. Acta Ecological Sinica, 2012, 32(13): 4181-4190.
[7] 刘启贞, 李九发, 戴志军, 等. 长江口颗粒态金属污染五时空分布规律分析[J]. 海洋环境科学, 2009, 28(3): 251-255.
LIU Q Z, LI J F, DAI Z J,etal. Spatial and temporal distribution of particulate metal pollutants in Changjiang Estuary [J]. Marine Environmental Scinece, 2009, 28(3): 251-255.
[8] 毕春娟, 陈振楼, 许世远, 等. 长江口潮滩大型底栖动物对重金属的累积特征[J]. 应用生态学报, 2006, 17(2): 309-314.
BI C J, CHEN Z L, XU S Y,etal. Heavy metals accumulation in macrobenthos in intertidal flat of Yangtze Estuary [J]. Chinese Journal of Applied Ecology, 2006, 17(2): 309-314.
[9] 陈 亮, 刘 一, 禹 娜, 等. 分水江水库浮游动物群落结构的初步研究及水质评价[J]. 华东师范大学学报(自然科学版), 2010(6): 72-82.
CHEN L, LIU Y, YU N,etal. Preliminary study on the metazoan zooplankton community structure of Fenshuijiang Reservoir and water quality assessment [J]. Journal of East China Normal University (Natural Science), 2010(6): 72-82.
[10] PAYLY D, CHRISTENSEN V, WALTER C J. Ecopath, ecosim and ecospace as tools for evaluating ecosystem impact of fisheries [J]. ICES Journal of Marine Science, 2000(57): 697-706.
[11] CHRISTENSEN V, PAULY D. ECOPATHⅡ-a software for balancing steady-state ecosystem models and calculating network characteristics [J]. Ecological Modelling, 1992, 61(3): 169-185.
[12] 仝 龄, 唐启升. 渤海生态通道模型初探[J]. 应用生态学报, 2000, 11(3): 435-440.
TONG L, TANG Q S. A Preliminary approach on mass-balance ecopath model of the Bohai Sea [J]. Chinese Journal of Applied Ecology, 2000, 11(3): 435-440.
[13] LIN Q, JIN X, ZHANG B. Trophic interactions, ecosystem structure and function in the southern Yellow Sea [J]. Chinese Journal of Oceanology and Limnology, 2013, 31(1): 46-58.
[14] CHEN J H, CHENG W W, PITCHER T J. Mass-balance ecosystem model of the East China Sea [J]. Progress in Natural Science, 2009(10): 1271-1280.
[15] 陈作志, 邱永松. 南海北部生态系统食物网结构,能量流动及系统特征[J]. 生态学报, 2010, 30(18): 4855-4865.
CHEN Z Z, QIU Y S. Assessment of the food-web structure, energy flows, and system attribute of northern South China Sea ecosystem [J]. Acta Ecologica Sinica, 2010, 30(18): 4855-4865.
[16] 张效嘉, 线薇薇. 1985-1986年长江口生态系统能流网络分析[J]. 海洋科学, 2016, 40(7): 60-72.
ZHANG X J, XIAN W W. Energy flow and network analysis of the Yangtze Estuary ecosystem during 1985-1986 [J]. Marine Sciences, 2016, 40(7): 60-72.
[17] 林 群, 金显仕, 郭学武, 等. 基于Ecopath模型的长江口及毗邻水域生态系统结构和能量流动研究[J]. 水生态学杂志, 2009, 2(2): 28-36.
LIN Q, JIN X S, GUO X W,etal. Study on the structure and energy flow of the Yangtze River Estuary and adjacent waters ecosystem based on ecopath model [J]. Journal of Hydroecology, 2009, 2(2): 28-36.
[18] 李 睿, 韩 震, 程和琴, 等. 基于ECOPATH模型的东海区生物资源能量流动规律的初步研究[J]. 资源科学, 2010, 32(4): 600-605.
LI R, HAN Z, CHENG H Q,etal. A preliminary study on biological resources energy flows based on the ecopath model in the East China Sea [J]. Resources Science, 2010, 32(4): 600-605.
[19] CHRISTENSEN V, WALTERS C, AHRENS R,etal. Database-driven models of the world's large marine ecosystems [J]. Ecological Modelling, 2009, 9(10): 1984-1996.
[20] LI Y K, YONG C, BING S,etal. Ecosystem structure and functioning of Lake Taihu (China) and the impacts of fishing [J]. Fisheries Research, 2009(95): 309-324.
[21] VILLY C, CARL J W, DANIEL P. Ecopath with ecosim: A user’s guide [D]. Vancouer Canada: Fisheries Centre University of British Columbia, 2004: 36-42.
[22] 韩 瑞, 陈求稳, 王 丽, 等. 基于生态通道模型的长江口水域生态系统结构与能量流动分析[J]. 生态学报, 2016, 36(15): 4907-4918.
HAN R, CHEN Q W, WANG L,etal. Analysis of the ecosystem structure and energy flow of the Yangtze River estuary and adjacent seas, based on the Ecopath model [J]. Acta Ecologica Sinica, 2016, 36(15): 4907-4918.
[23] ULANPWICZ R. Ecosystem trophic foundations: Lindeman exonerata. Complex ecology: The part-whole relation in ecosystems prentice-hall [J]. Englewood Cliffs, 1995: 549-560.
[24] JACKSON B C, KIRBY M X, BERGER W H,etal. Historial overfishing and the recent collapse of coastal ecosystems [J]. Science, 2001(293): 629-637.
[25] LINDEMAN R L. The trophic-dynamic aspect of ecology [J]. Ecology, 1942(23): 339-417.
[26] ODUM E P. The strategy of ecosystem development [J]. Science, 1969(164): 262-270.
[27] 武 震, 贾佩峤, 胡忠军, 等. 基于Ecopath模型分析分水江水库生态系统结构和功能[J]. 应用生态学报, 2012, 23(3): 812-818.
WU Z, JIA P Q, HU Z J,etal. Structure and function of Fenshuijiang Reservoir ecosystem based on the analysis with Ecopath model [J]. Chinese Journal of Applied Ecology, 2012, 23(3): 812-818.
[28] CHRISTENSEN V. Ecosystem maturity-towards quantification[J]. Ecological Modelling, 1995, 77(1): 3-32.
[29] 罗秉征, 韦 晟, 窦硕增. 长江口鱼类食物网与营养结构的研究[J]. 海洋科学集刊, 1997(3): 143-153.
LUO B Z, WEI S, DOU S Z. Study on food web and trophic structure of fish in the Changjiang river estuary [J]. Studia Marina Sinica, 1997(3): 143-153.
[30] 李建生, 程家骅. 长江口渔场渔业生物资源动态分析[J]. 海洋渔业, 2005, 27(1): 33-37.
LI J S, CHENG J H. Analysis on the dynamic of fishery biological resources on the fishing grounds of the Changjiang estuary [J]. Marine Fisheries, 2005, 27(1): 33-37.
[31] 陈吉余, 陈沈良. 近20年来长江口生态系统变化[J]. 净水技术, 2002, 1(4): 1-3.
CHEN J Y, CHEN S L. Ecological environment changes in the Changjiang Estuary over recent 20 years [J]. Water Purification Technology, 2002, 1(4): 1-3.
[32] RYDER R A, KERR S R, TAYLOR W W,etal. Community consequences of fish stock diversity [J].Canadian Journal of Fisheries and Aquatic Sciences, 1981(38): 1856-1866.
[33] FINNEY B P, GREGORY-EAVES I, DOUGLAS M S V,etal. Fisheries productivity in the northeastern Pacific Ocean over the past 2 200 years [J]. Nature, 2002(416): 729-733.
[34] 陈作志, 邱永松, 贾晓平, 等. 基于Ecopath模型的北部湾生态系统结构和功能[J]. 中国水产科学, 2008, 15(3): 460-468.
CHEN Z Z, QIU Y S, JIA X P,etal. Structure and function of Beibu Gulf ecosystem based on Ecopath model [J]. Journal of Fishery Sciences of China, 2008, 15(3): 460-468.
猜你喜欢
法人(2022年3期)2022-03-30
阅读(快乐英语高年级)(2022年3期)2022-03-30
当代水产(2021年10期)2022-01-12
海洋通报(2021年1期)2021-07-23
海洋通报(2020年2期)2020-09-04
中国油脂(2019年4期)2019-04-29
伴侣(2018年2期)2018-02-28
教育教学论坛(2016年49期)2017-02-27
河北渔业(2014年11期)2014-11-17
岩土力学(2014年1期)2014-01-20
