
扎龙湿地不同生境芦苇分株构件生物量动态分析
2018-06-27 13:03徐嘉政于欣宇陈雨竹
防护林科技 2018年6期
徐嘉政,于欣宇,陈雨竹
(1.东北农业大学 资源与环境学院,黑龙江 哈尔滨 150300;2.齐齐哈尔大学 生命科学与农林学院,黑龙江 齐齐哈尔 161006;3.东北农业大学 动物科学技术学院,黑龙江 哈尔滨 150300)
1977年Harper提出植物种群构件理论[1],把植物种群划分群落水平和个体水平相关联又截然不同的层次,构件理论的建立,为植物种群生态学的深入研究开拓了新途径[2]。
植物对环境的适应性变化首先表现在构件上,通过构件间的相互作用形成了基础特定的形态特征,因此,植物构件的大小不仅能够反映出植物个体及种群的生长发育节律,也反映了其对环境的适应能力,进而体现了环境条件对植物个体及种群的影响和饰变程度[3]。在异质生境中,分株构件大小虽具有较大的可塑性,均可在种群和分株等水平上进行检测[4],朝鲜碱茅在孕穗期和果后营养期,叶构件的生物量及其占总生物量的比率均达到最大值,这是叶构件重要功能的体现,也是种群对营养物质的分配策略之一[5]。野生葎草茎可反映植株的空间拓展能力,其数量体现无性系的再生能力,匍匐株与攀缘株以不同的角度和时间接受光照,植株的生长采取不同的拓展策略,确保足够的光合面积,野生葎草在种群拓展上表现出多样性[6]。因此,开展构件水平种群与环境因子关系的研究,不仅可以获得种群对环境因子反应以及对某些环境因子适应性等重要信息,并且在一定程度上,对揭示无性系植物群落结构的形成及群落相对稳定均具有重要意义[7]。
芦苇为典型长根茎型无性系植物,天然种群主要依靠营养繁殖补充和更新种群[8]。分株为芦苇种群的现实种群,分株主要由叶片、叶鞘、茎构件组成,构件生物量是种群物质生产的重要参数,在一定程度上可以反映生产力及生长环境条件的优劣。因此,开展扎龙湿地芦苇分株构件生物量及其分配规律的动态研究,可以了解芦苇分株构件水平的物质生产和分配规律,为揭示芦苇的表型可塑性和趋异适应机理提供基础,也可为扎龙湿地的科学管理提供理论支持。
1 材料与方法
1.1 研究区概况
本项研究在扎龙湿地进行。46°52′—47°32′ N,123°47′—124°37′ E,该地区属于中温带大陆性季风气候,年平均气温为3.2 ℃,最冷月平均气温-19.4 ℃,最热月平均气温22.9 ℃,年日照时数2 700~3 000 h,年辐射总量是2 052~2 140 Jcm-2,积温2 600~3 000 ℃ ,平均地温4.9 ℃,无霜期128 d[9,10]。
1.2 样地设置
在扎龙湿地,以土壤水分条件,从旱生、湿生、水生及盐碱化土壤,共设置4个生境样地。
1.2.1 旱生生境(H1) 土壤为草甸土,全年无积水或7—8月雨季短时积水,经常形成羊草+芦苇群落,微地形中心也可形成面积不等的单优种芦苇群落斑块[11,12]。
1.2.2 湿生生境(H2) 土壤为沼泽土,平时无积水或少量积水,7—8月雨季来临以后逐渐积水,积水深度一般情况低于30 cm[11,12]。
1.2.3 水生生境(H3) 土壤为腐殖沼泽土,形成湿地内面积最大的芦苇群落,常年积水,水深变化为10~100 cm[11,12]。
1.2.4 盐碱生境(H4) 土壤为盐碱土,全年无积水,土壤表层已完全丧失,具有明显盐碱斑,有的碱斑可形成单优势种芦苇居群[11,12]。
1.3 研究方法
按照芦苇的生育期特性确定取样时间,于2016年6月8日(营养生长初期),7月10日(营养生长旺盛期),8月10日(生殖生长初期),9月9日(生殖生长旺盛期),10月8日(休眠期),分别在上述4个样地内随机抽取30个分株,齐地剪取带回实验室,按叶片、叶鞘和茎构件分别装入信封,置于80 ℃烘箱烘至恒质量并称量。
1.4 数据处理
用 Excel2007绘制芦苇分株构件生物量动态和生物量分配图。对不同月份和不同生境间构件生物量比率差异进行单因素方差分析(One-way ANOVA,ɑ=0.05)
2 结果与分析
2.1 芦苇分株构件生物量的时空动态
芦苇分株不同构件生物量动态如图1-图3所示。从图中可以看出,整个生育期内,4个生境芦苇分株叶片、叶鞘、茎的生物量均呈现出比较一致的变化规律,从营养生长初期的6月开始,各构件生物量均呈逐渐升高的趋势,至生殖生长旺盛期的9月达到最大值,然后到休眠期的10月又均有不同程度的降低。由此表明,芦苇分株不同构件的生物量存在比较的稳定季节性变化规律,这种变化规律的稳定性是遗传因素所决定的生育期节律。

图1 不同生境芦苇分株叶片生物量动态

图2 不同生境芦苇分株叶鞘生物量动态

图3 不同生境芦苇分株茎生物量动态
从图1-图3还可知,芦苇分株叶片、叶鞘和茎构件生物量在不同生境中具有相对稳定的差异和差异序位。6—10月,各构件生物量均表现为以水生生境(H3)为最高,依次是旱生生境(H1)和湿生生境(H2),盐碱生境(H4)为最低。由此表明,不同生境的芦苇分株构件在物质生产上具有较大的生态可塑性,并表现出生境条件对生物量的影响和饰变程度。
2.2 芦苇分株构件生物量的分配
芦苇分株不同构件生物量的分配比率如图4-图7所示,从图中可知,整个生育期内,4个生境芦苇分株叶片和叶鞘的生物量比率均呈降低的趋势,而茎的生物量比率均呈升高的趋势。其中叶片的生物量比率在18%~39%,叶鞘的生物量比率在9%~33%,茎的生物量比率在30%~74%,总体趋势是茎的生物量比率最高,其次是叶片的生物量比率,均以叶鞘的生物量比率最低。各构件执行不同的功能,在物质分配上的差异正是芦苇固有的生物学特性的具体体现。
从图4~图7还可知,整个生育期内,4个生境芦苇分株的叶片、叶鞘、茎的生物量比率分别为旱生生境:22.8%,17%,60.2%; 湿生生境:24.2%,18.4%,57.4%;水生生境:23%,16%,51%;盐碱生境:28%,21.4%,50.6%。经统计分析,不同生境条件叶片、叶鞘和茎构件的生物量分配比率差异均未达显著水平(P>0.05)。分株叶片、叶鞘和茎构件间生物量分配比率的相对稳定体现着光合产物的转移规律,分株的物质分配随生育期而发生着优势换位现象,不同生境分株构件的生物量分配比率却相对稳定。

图4 旱生生境芦苇分株不同构件生物量分配比率
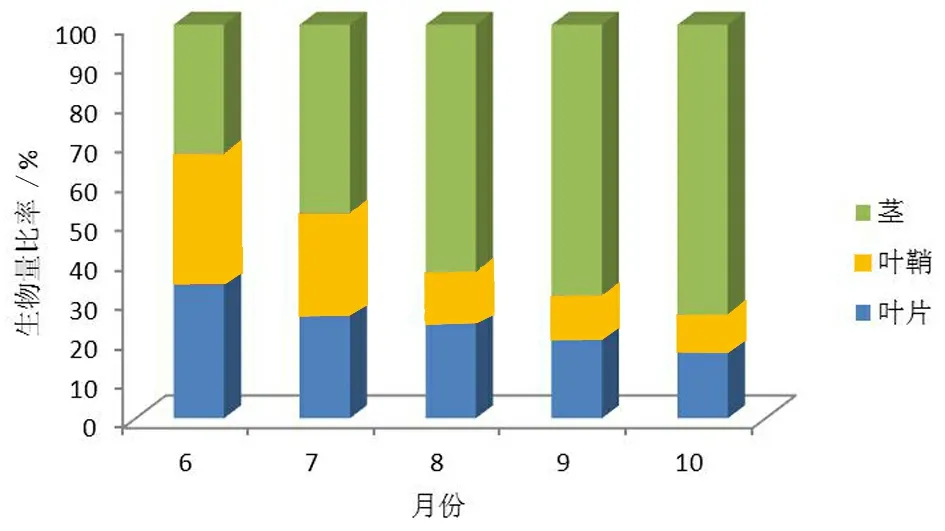
图5 湿生生境芦苇分株不同构件生物量分配比率

图6 水生生境芦苇分株不同构件生物量分配比率

图7 盐碱生境芦苇分株不同构件生物量分配比率
3 结论与讨论
构件植物是由构件单位组成的微种群,构件植物包括由基株形成的个体种群和基株上的构件单位形成的构件种群,这两个水平的种群动态既互相影响,又互相依赖,种群大小和密度的调节也由两个结构水平在环境作用下共同反应的结果[13]。
芦苇分株包括支持构件(茎)、光合构件(叶片)、保护构件(叶鞘)。分株构件生物量的动态是芦苇生物学特性及生长发育过程的具体表现, 扎龙湿地芦苇具有特定的生育期,6-10月依次为营养生长初期,营养生长旺盛期,生殖生长初期,生殖生长旺盛期,休眠期。各构件生物量从6月持续增长至9月,然后又略有降低。构件的生物量与分株生物量具有一致的变化规律。已有研究表明芦苇分株生物量从返青后持续增加到生殖生长旺盛期的9月,至休眠期的10月又有所降低,生物量呈“单峰型”变化[7,14,15],在生长季后期,芦苇种群将生产的物质主要分配转移到地下储藏与营养繁殖器官的形成与生长上[7]。因此,4个生境芦苇分株种群和构件中间在生长及物质生产上表现出共同的生物学特性和一致的时间节律性,这种稳定性的过程是遗传因素所控制的,一般较难改变。各构件生物量在生境间均表现为以水生生境为最高,依次是旱生生境、湿生生境,盐碱生境为最低。均以常年积水或季节性积水的水生环境最高,以既无临时积水,土壤条件又恶劣的碱斑生境最低,这一点与分株生物量生境间的差异和差异序位也完全一致,均表现出生境差异对分株和构件生长的饰变影响。
尽管分株和构件的生物量存在着季节动态和生境间差异,但整个生育期内,生境间各个构件生物量分配的比率的差异均未达到显著水平(P>0.05)。其中,叶片的分配比率在22.8%~28%,叶鞘的分配比率在16%~21.4%,茎的分配比率在50.6%~61%, 分株将50%以上的物质投入给茎,其余的物质分配给叶片和叶鞘,这种物质分配也充分体现芦苇的营养生长策略,分株为了获取更多的O2、CO2及光资源,将较多的物质分配给茎的伸长生长,以提高个体对空间和资源的利用[16]。不同生境芦苇分株构件生物量间的差异与生境条件密切相关,各构件生物量分配比率反映了芦苇在物质分配方面的生物学特性是相对稳定的。
参考文献:
[1] Harper J L. Population Biology of plants [M]. London and New York:Academic Press,1977
[2] 钟章成,曾波.植物种群生态研究进展[J].西南师范大学学报:自然科学版,2001,26(2):230-236
[3] Arenas F,Viejo RM,Fernández C. Density-dependent regulation in aninvasive seaweed: responses at plant and modular levels[J]. Journal of Ecology,2002,90: 820-829
[4] Harper J L. The concept of population in modular organisms//May R M,ed. Theoretical Ecology: Principles and Applications. London: Blackwell,1981: 53-77
[5] 刘佩勇,张庆灵,杨允菲.松嫩平原朝鲜碱茅无性系种群构件生物量结构及相关模型分析[J].应用生态学报,2004,15(4):543-548
[6] 刘金平,王艳.南充地区野生蓰草构件性状多样性分析[J].草业科学,2009,26(8):68-71
[7] 焦德志,荣子,曹瑞,等.扎龙湿地保护区异质生境芦苇种群分株构件的数量特征[J].生态学报,2017,37(23):7843-7853
[8] 焦德志,么璐,黄曌月,等.东北草地异质生境芦苇种群根茎芽年龄结构及输出规律[J].生态学报,2015,35(2):370-377
[9] 中华人民共和国林业部.扎龙国家级自然保护区管理计划北京[M]:北京:中国林业出版社,1997:1-5
[10] 赵魁义.中国沼泽志[M].北京:科学出版社,1999:231-239
[11] 焦德志,么璐,黄曌月,等.东北草地异质生境芦苇芽种群动态[J].应用生态学报,2015,26(2):404-410
[12] 焦德志,黄曌月,周婵,等.扎龙湿地异质生境芦苇种群根茎动态及年龄结构[J].生态学杂志,2016,35(4):888-895
[13] Mccue K F,Hanson A D. Salt-inducible betaine aldehyde dehydrogenase from sugar beet: cDNA cloning and expression[J]. Plant Molecular Biology,1992,18(1):1-11
[14] 许秀丽,张奇,李云良,等.鄱阳湖洲滩芦苇种群特征及其与淹水深度和地下水埋深的关系[J].湿地科学,2014,12(6):714-722
[15] 夏贵菊,何彤慧,赵永全,等.银川平原芦苇生态特征与土壤因子的关系[J].土壤通报,2015,46(1):99-104
[16] 罗文泊,谢永宏,宋凤斌.洪水条件下湿地植物的生存策略[J].生态学杂志,2007,26(10):1478-1485
猜你喜欢
落叶果树(2021年6期)2021-02-12
伙伴(2020年4期)2020-05-11
读者(2018年14期)2018-06-28
农机化研究(2018年2期)2018-06-05
青年文学家(2017年28期)2017-11-28
青年文学家(2016年34期)2017-03-31
植物保护(2016年1期)2016-09-14
体育科技(2016年2期)2016-02-28
西华师范大学学报(自然科学版)(2015年3期)2015-02-27
应用海洋学学报(2014年2期)2014-11-26
