
基于食性集团的爱鸟周人工投食建议
2018-06-26 07:57:02李祎斌吴庆明刘新宇
野生动物学报 2018年2期
李 翔 李祎斌 吴庆明 隋 媛 刘新宇
沙力瓦·拍依祖拉木 何富英 夏丹霞
(东北林业大学,哈尔滨,150040)
鸟类是自然界的重要组成部分,是自然界中营养循环、能量流动时空跨度最大的群体之一[1-2],与人类的物质文明、精神文明、生态文明密不可分,对人类的可持续生存具有重要的生态价值、经济价值和社会价值[3-4]。爱鸟周是我国目前唯一的以法规形式开展的鸟类保护行动[5];它源于1981年3月3日和日本正式签订的《中华人民共和国政府和日本国政府保护候鸟及其栖息环境协定》;同年9月,林业部等8个部门联名提出《关于加强鸟类保护,执行中日候鸟保护协定的请示》报告,其内容之一是向国务院建议:在每年的4~5月初确定一个星期为各省、市、自治区等区域的“爱鸟周”。1982年2月,国务院发出了关于保护鸟类的通知,并同意林业部等8单位提出的关于每年各省市都要开展“爱鸟周”宣传活动的建议。1992年国务院批准的《陆生野生动物保护条例》中将“爱鸟周”以法规的形式确定下来。
截至目前,我国的每个省、不同省内的不同市县乃至部分保护区等相关部门均有了自己的爱鸟周,具体时间从2月至5月不一[6-7]。爱鸟周开展的内容也多种多样[8-9],包括学术报告、课堂宣讲、悬挂人工鸟巢、宣讲标本形式的鸟类知识、发放和张贴鸟类手册和宣传画、绘画签名、鸟类观察等,有些区域还有相应的晚会、考察;甚至还有人工投食。尤其是后者,近年来频繁出现在不同区域的爱鸟周中,成为爱鸟周活动中比较引人注目的一个内容。人工投食的食物种类各式各样,投食方式也各异,有的投食鱼类,直接倒入路边水域;有的投食玉米等谷物,直接扬洒入道边的湿地沼泽中;有的投食瓜子松子等壳类食物,直接放置于林下。而且,部分市民和部分高校的学生组织也在积极地制作各种投食器。所有的这些爱鸟周中的人工投食,不论是食物种类还是投放位置,均未考虑觅食对象的针对性、偏好性、栖息特性以及安全性。
众所周知,鸟类的食物具有种间特异性、生活史时期特异性[10-11],不科学的投食和放置,不仅不能起到真正的食物补充作用,还可能带来相应的副作用。以鸟类为例,若真的需要人工投食,应该科学投食,至少应了解该区域内鸟类的种类数量、时空分布、集团结构等。基于此,本文针对爱鸟周中的人工投食活动,以林栖鸟类为投食对象,以东北林业大学校区及其附属的城市林业示范基地为调查地点进行了调查分析。
1 研究地与研究方法
1.1 研究地概况
研究区域为东北林业大学校区及其附属的城市林业示范基地,地理位置为 N45°43'10″,E126°37'15″,总面积136 hm2(含绿化面积超过50 hm2),是省内高校中校园绿化面积最大的区域;该区域位于哈尔滨市中心偏南地段的香坊区与南岗区交界处,马家沟穿区而过[12]。区域内的人工林树种基本涵盖我国东北的主要树种,多样性较高,树种面积大小不一,以樟子松(Pinus sylvestris)、胡桃楸(Juglans mandshurica)、落叶松(Larix gmelinii)、黑皮油松(Pinus tabuliformis)、水曲柳(Fraxinus mandschurica)、蒙古栎(Quercus mongolica)、榆树(Ulmus pumila)、白桦(Betula platyphylla)、黄檗(Phellodendron amurense)等为主[13-14]。该研究区域因与哈尔滨市江南区域绿地面积最大的黑龙江省森林植物园为邻,每年除了有一些固定种类与数量的鸟种在校区繁殖外,春秋迁徙季节还有一些栖息于植物园的鸟类客串于本校区[15-16]。

图1 样线分布图Fig.1 Distribution map of sampling line
1.2 研究方法
1.2.1 鸟类观察
2017年6~9月,通过双筒望远镜采用样点法、样线法对东北林业大学校园内栖息分布的鸟种及其数量进行了调查。共设置了8条样线41个样点,样线布置以校园内的道路为准、长短不一,调查时的步行速度为2.0~3.0 km/h[16]。实际的调查每隔1 d进行1次,选择在当天5:40开始调查(这是学校寝室的开门时间,也是允许我们出楼观鸟的最早时间),此时鸟类活动频繁,遇见率高,便于统计种类和数量。
记录的种类以预定的观测范围内出现的种类为主;对于难以识别的和繁殖季节不易发现的个别物种,标记其出现的和可能出现的区域,采用定点观察法进行补充观测,并通过照相机对鸟种的体态特征和鸣叫特征进行拍照和录像,最后结合《中国鸟类图志》、《东北鸟类图鉴》及东北林业大学校园内鸟类分布的历史名录进行鸟种鉴定[17-18]。
记录的数量以预定的观测范围内单侧飞行的数量为主,实际记录时每个方向的鸟种数量均记录,调查结束后根据实际情况进行单方向统计核实,最大程度降低重复记录所引起的原始数量误差。
1.2.2 数据分析
对调查的数据首先进行了种类组成和优势度分析;然后基于食物类型、取食方式、取食空间3个维度进行了集团划分[19-20],以期对生态习性相似的鸟类进行集团分析。
其中,优势度分析采用频率指数估计法[21-23],具体算法为:RB=(100 d/D)×(S/D)。式中,d为遇见鸟类的天数;D为工作的总天数;S为遇见鸟类的总数量;RB为优势度指数,若RB≥500则定义为优势种,500>RB≥50为常见种,50>RB≥5为少见种,5>RB为偶见种。
鸟类集团划分主要基于食物类型、取食方式、取食空间3个维度的数据进行了聚类分析。其中,食物类型具有生活史时期特点,不同生活史时期不同鸟类的食物类型不同、同一种鸟类的成鸟与幼鸟间食物类型也存在一定的差异,本文以观察期间鸟类所处的生活史阶段为准,相应内容不涉及观察期以外的生活史阶段的食物类型[24-25],分为动物性食物和植物性食物。
取食方式,本文根据食物所处位置和鸟体喙部的深入程度[10]分为:(1)拾食,即食物位于同质实物基质表层或浅层、不需要处理障碍物直接用喙即可夹取的取食方式;(2)啄食,即食物位于同质实物基质浅层或深层、需要用喙啄开障碍物获取食物的取食方式;(3)探食,即食物位于异质实物基质的浅层或深层,需要喙深入第一层基质探取或穿过第一层基质探取第二层基质中食物的取食方式如涉禽类;(4)捕食,即食物位于非实物基质中(如空中),需要鸟类快速运动身体的部分或全部进行捕捉食物的方式。
取食空间,基于本文观察到的鸟类特点按照鸟类取食时所在的基质类型和基质空间分为[26]:(1)地面(0 m);(2)林下层(0~2 m);(3)林中层(2~10 m);(4)林上层(10 m以上至树冠);(5)树冠;(6)空中。
所有的数据通过软件Excel 2010进行统计整理,在软件SPSS 24.0和R 3.4.3(R Development Core Team 2011)中进行了分析。
2 结果与分析
2.1 物种组成及优势度分析
研究期间,研究区域共记录鸟类24种(表1);其中,以雀形目(Passeriformes)鸟类占绝对优势,为19种,占79.17%;鹳形目(Ciconiiformes)、隼形目(Falconiformes)、鸽形目(Columbiformes)、雨燕目(Apodiformes)及鴷形目(Piciformes)各1种,各占4.17%。
优势度分析显示出:优势种较少(8.33%),仅有2种,分别为伴人鸟种麻雀(Passer montanus)和喜鹊(Pica pica);其次是常见种(16.67%),分别为家燕(Hirundo rustica)、金腰燕(Hirundo daurica)、灰喜鹊(Cyanopica cyanus)、普通鳾(Sitta europaea);少见种相对较多(25.00%),分别为大山雀(Parus major)、大斑啄木鸟(Dendrocopos major)、雀鹰(Accipiter nisus)、白眉姬鹟(Ficedula zanthopygia)、黄眉柳莺(Phylloscopus inornatus)、白眉鹀(Emberiza tristrami);其余均为偶见种,所占比例也最多(50.00%)。
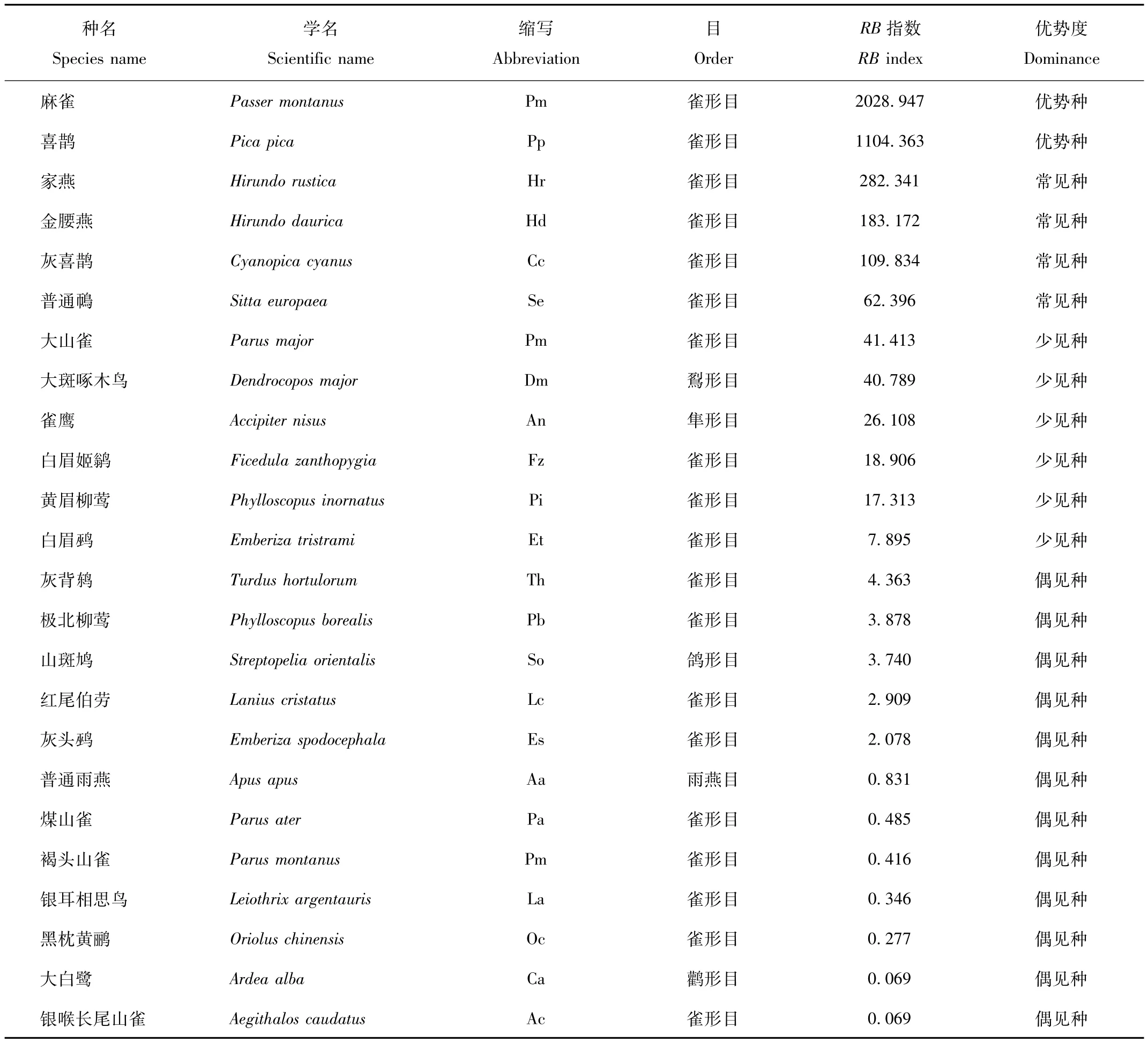
表1 鸟类群落的物种组成及优势度分析Tab.1 Species composition and dominance analysis of avian community
2.2 集团结构分析
根据食物类型、取食方式、取食空间3个维度12类数据通过聚类分析将24种鸟分为5类,分别将其定义为:中下层偏植杂食啄拾集团、中下层偏肉杂食啄食集团、中上层偏肉杂食捕食集团、空中食肉捕食集团、地面食肉探食集团(图2)。
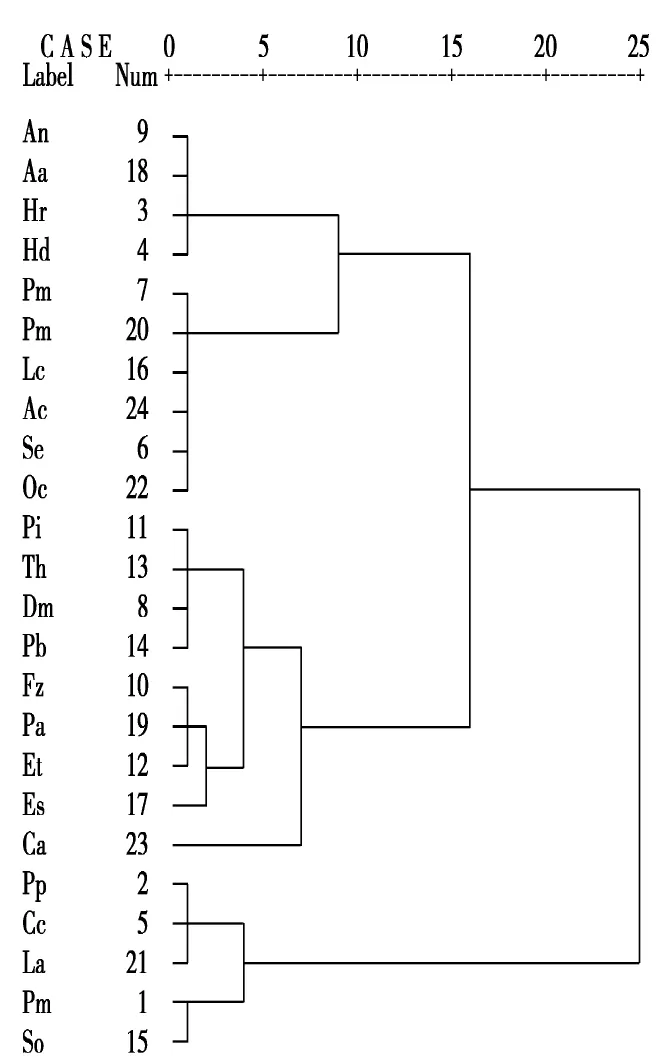
图2 基于取食特点的鸟类集团结构Fig.2 Avian guild structure based on avian diet characteristic
2.2.1 中下层偏植杂食啄拾集团
即栖息于林中中下层位置且偏好植物的杂食性鸟类,该类鸟类取食方式为啄取、拾取,由麻雀、喜鹊、灰喜鹊、山斑鸠、银耳相思鸟等5种鸟类组成。其中,银耳相思鸟不属于该区域应分布的鸟种,推测其为从鸟市或鸟主逃逸出来的个体,观察期间仅1次出现在观察区域内。
2.2.2 中下层偏肉杂食啄食集团
即栖息于林中中下层位置且偏好动物性食物的杂食性鸟类,该类鸟类取食方式为啄食,由大斑啄木鸟、白眉姬鹟、黄眉柳莺、白眉鹀、灰背鸫、极北柳莺、灰头鹀、煤山雀等8种鸟类组成。该类鸟类多为食虫鸟类;其中灰头鹀、白眉鹀本为食谷鸟类,但限于观察期为二者的夏季繁殖期和繁殖后期,该期间为了雏鸟更高效率的增长,食物以昆虫为主,故灰头鹀、白眉鹀被列为该类集团中。
2.2.3 中上层偏肉杂食捕食集团
即栖息于林中中上层位置且偏好动物性食物的杂食集团,该类鸟类的取食方式以捕食为主,由普通鳾、大山雀、红尾伯劳、褐头山雀、黑枕黄鹂、银喉长尾山雀等6种鸟类组成。
2.2.4 空中食肉捕食集团
即常于空中捕食动物性食物的鸟类,由雀鹰、普通雨燕、金腰燕、家燕等4种鸟类组成。
2.2.5 地面食肉探食集团
即多栖息于地面通过探索的方式取食同类基质或异类基质中的动物性食物,该类仅大白鹭1种鸟类。该区域观察到的大白鹭由植物园方向飞来。
2.3 集团鸟种优势度分析
由图3可知,上述分析的5个集团鸟种的总优势度具有明显差异,其中中下层偏植杂食啄拾集团鸟种的总优势度在研究区域占有绝对优势,为3247.23,超过总鸟类群落的80%;其次为空中食肉捕食集团,超过总鸟类群落的10%;其余集团所占比例较小,均低于总鸟类群落的3%。

图3 不同鸟类集团鸟种的总优势度Fig.3 Total dominance of avian species in different avian guild
3 讨论
3.1 物种组成
关于鸟类的物种组成,相比于校园面积而言确实有些少,远低于夏丹霞[16]等的观测结果。综合分析,可能的影响因素主要有两种:其一是调查期间短且仅为繁殖期,该时期鸟类的活动频率低且活动区域比较隐蔽,再加上直接的人车活动影响和间接的城市噪音干扰导致下的数量方面的限制,观测机率较小是客观存在的现象,针对这种现象,我们采用了双筒望远镜和单筒望远镜相结合的方式,同时辅以400 mm的单反照相机和录音设备,针对露面时间较短的个体和只有声音的个体进行动静式的信息收集,然后进行进一步种类的核对,以最大限度地减小观察误差;其二是调查区域内人类活动较多,虽然选择了早晨这种一天中最为清净的时间段,但区域内师生员工家属及车辆晨读晨练等干扰活动仍较多,仍会一定程度地从声音、活动频率、活动强度等方面影响鸟类的活动范围和出现频率。
3.2 集团划分
关于集团划分结果,本文的结果中出现了食物类型不同却划分为一个集团的现象,如灰头鹀、白眉姬鹟等。这方面的现象存在两方面的解释。一方面是调查期间为繁殖期的缘故。繁殖期是鸟类生活史周期中的关键时期,决定着鸟类的种群数量及下一阶段的生存,尤其是需要长距离迁徙的候鸟,其整个繁殖期必须完成卵孵化、雏鸟生长发育、雏鸟飞翔训练等环节。这些环节都需要高能量食物的供给,而且时间存在限制,杂食鸟类食物组成或我们常规中所认知的鸟类食物组成会发生季节性变化,其相应的取食方式等也会发生改变,这是鸟类适者生存的必然选择。另一方面是调查区域为城市环境的缘故。该区域被人为环境和人为活动所包围和伴随,鸟类某一类食物会存在食物量突然不足而迅速觅食另一类食物的快速适应现象。所有这两方面的解释,也许是鸟类针对人为环境产生的各类环境因素挑战而做出的快速适应现象,这方面有待于通过野外环境和城市环境鸟类行为方面的对比研究来进而揭示鸟类城市化适应的行为策略。本研究仅为一个调查季节,相应的数据也存在特殊性的现象,期待能在下一步的研究中进行验证。
3.3 人工投食建议
关于人工投食,科学的投食之前应该考虑的因素很多,需要投食前的科学考察,不可盲目地人工投食。投食之前应根据投食对象进行前期准备,投食对象不同,投食的工作流程也不同。
如果针对某一单一鸟种进行人工投食,前期准备工作应该更细更具体,主要包括:(1)不仅需要对该鸟种栖息区域内食物量及其可获得性进行评估;(2)而且需要对该区域内存在的对食物量有影响的人为干扰做出判断;(3)更需要对该种鸟类取食的行为特点进行资料收集或持续性观察分析;(4)还需要考虑人工投食对该鸟种野生行为的影响评估等,只有这样才可以对单一鸟种进行人工投食。而在我们常见的爱鸟周的人工投食中,食物往往直接投入生境边缘的道路附件区域,不仅未对人为干扰做出判断,对该区域内的食物量、可获得性、投食对象的取食行为特点以及对鸟类野生行为的影响等方面均未进行评估,这是非常不科学的做法,不值得提倡。关于这方面可以针对本文的研究结果进行下一步研究。
如果对某一集团鸟类进行人工投食,在单一鸟种角度评估的基础上,不仅应该从整体角度对投食区域内的鸟类群落的物种组成、居留型、优势种等方面进行系列调查,还应进一步对鸟类群落的集团结构进行深入分析,尤其是调查资源贫瘠区域更应该成为重中之重,根据这些综合性分析结果决定人工投食的器材准备、食物准备、投放事项等内容。否则,人工投食必然会存在盲目性,轻则浪费材料,中则会给自然环境增加不和谐的因素,重则会给一些鸟类的安全带来相应的副作用如安全隐患等。
本文的人工投食分析属于后一种,多鸟种组成的集团结构。基于该分析结果,如果需要对东北林业大学校园及其附属的城市林业示范基地内繁殖期栖息的鸟类进行人工投食,可以根据集团结构分析结果、集团鸟种优势度等方面进行食物类别和投放位置的选择性实施。其中,(1)中下层偏植杂食啄拾集团的鸟种成员优势度最高,超过该区域的80%,但该集团鸟种绝大部分为伴人鸟种,是否需要人工投食,此处不议;若必须投食,可以选择林中10 m以下的中下层位置地面放置或悬挂适于啄食的适合该集团鸟种的粒状或条状食物。(2)空中食肉捕食集团是该区域第二大优势集团,超过10%,该类集团鸟种的投食难度较大,不适合作为普通方式下食物投放的对象;如果真的需要投食,需要长期保持或增加该区域植被群落环境的多样性与自然性,这样自然而然就能增加区域内昆虫类的多样性。(3)中下层偏肉杂食啄食集团的优势度较低,基本属于野生鸟种,因此不是环境中的任何位置都适合其繁殖期利用,再加上区域内的人类活动以及繁殖安全距离影响,区域内能利用的繁殖区域是非常有限的,对该类鸟类进行投食很有必要。关于肉类食物,可以像上述一样尽量保持或增加植被环境的多样性与自然性,通过增加各类昆虫多样性的方式来增加其环境中的肉类食物量,这也适合中上层偏肉杂食捕食集团的鸟种。此外,还可以通过地面放置或林中10 m以下悬挂的方式投放适合该集团鸟种的适于啄食的粒状或条状食物。(4)中上层偏肉杂食捕食集团也是优势度较低的野生鸟类,其投食方式除了维持自然环境外,还可以在林中10 m以上位置悬挂投食器投放适合该集团成员的植物性食物。(5)地面食肉探食集团是该区域的特殊现象,此处不加以讨论。
因此,建议相关管理部门在一些常规性的鸟类管护活动中,一定要慎重人工投食,必须科学投食。
[1] 何嘉乐,刘哲铭,许青.中国东北地区小型鸟类尸体分解利用的分析[J].野生动物学报,2017,38(3):414-421.
[2] 王玄,江红星,张亚楠.稳定同位素分析在鸟类食性及营养级结构中的应用[J].生态学报,2015,35(16):5556-5569.
[3] 王玲,丁志锋,胡君梅,等.广州城市绿地中鸟类对食源树种的偏好 [J].四川动物,2016,35(6):838-844.
[4] 徐世才,沈雪健,王莉,等.陕西子午岭国家级自然保护区湿地鸟类群落特征 [J].生物学杂志,2016,33(4):52-55.
[5] 马逸清.我国的爱鸟周活动[J].自然资源研究,1983(2):80-81,55.
[6] 动物学杂志编辑部.有关我国自然保护区(动物部分)和全国爱鸟周活动情况[J].动物学杂志,1983,18(2):64.
[7] 李春源.爱鸟周在各地[J].野生动物,1982,3(3):56.
[8] 关注候鸟保护守护绿色家园——第34届爱鸟周(2015年4月1日至7日)[J].林业与生态,2015(4):2,50.
[9] 中国野生动物保护协会.让我们和鸟儿共享蓝天——全国“爱鸟周”30周年[J].森林与人类,2011(3):6-9.
[10] 尚玉昌.鸟类的食物多样性及其取食适应[J].生物学通报,2007,42(3):9-13.
[11] 周立志,宋榆钧,马勇.紫蓬山区三种鹭雏鸟的食物多样性比较 [J].生态学杂志,2000,19(3):66-68.
[12] 张鹏宇,孙龙,胡海清.模拟计划火烧对哈尔滨城市林业示范基地典型林型土壤呼吸的影响[J].森林工程,2015,31(3):1-6.
[13] 孙元,兰晓萍,邵红涛,等.东北林业大学城市林业示范基地弹尾虫水平分布研究 [J].黑龙江大学工程学报,2013,4(3):51-54.
[14] 张晶虹,刘丙万.东北林业大学城市林业示范基地蒙古栎种群扩散研究[J].现代农业科技,2013(12):135-137.
[15] 王舒,童玉平,王志洁,等.哈尔滨市城市化建设对鸟类群落的影响[J].野生动物学报,2015,36(3):295-302.
[16] 夏丹霞,何富英,吴庆明,等.东北林业大学校园鸟类群落多样性 [J].野生动物学报,2017,38(2):258-264.
[17] 段文科,张正旺.中国鸟类图志[M].北京:中国林业出版社,2017.
[18] 常家传,桂千惠子,刘伯文,等.东北鸟类图鉴 [M].哈尔滨:黑龙江科学技术出版社,1995.
[19] 陆舟,杨岗,赵东东,等.广西弄岗喀斯特森林繁殖鸟群落的集团结构[J].动物学研究,2013,34(6):601-609.
[20] 陈锦云,周立志.安徽沿江浅水湖泊越冬水鸟群落的集团结构[J].生态学报,2011,31(18):5323-5331.
[21] 晏玉莹,匡中帆,江亚猛,等.贵州桐梓黄莲柏菁自然保护区夏季鸟类资源初步调查 [J].野生动物学报,2017,38(2):265-272.
[22] 程嘉伟,邓昶身,鲁长虎.苏州太湖湖滨人工种植和原生芦苇湿地鸟类群落[J].动物学杂志,2014,49(3):347-356.
[23] 谢光勇,李言阔,李佳,等.鄱阳湖都昌候鸟自然保护区夏季鸟类群落结构[J].四川动物,2014,33(1):139-143.
[24] 孙太福,聂圣鸿,国天曰,等.曲阜孔林鸟类多样性和季节变化 [J].野生动物学报,2017,38(3):462-466.
[25] 李慧,洪永密,邹发生,等.广州市中心城区公园鸟类多样性及季节动态[J].动物学研究,2008,29(2):203-211.
[26] 邹红菲,赵钢.兴凯湖自然保护区湖岗天然带状林中繁殖鸟巢的空间格局[J].野生动物,2008,29(1):21-25.
猜你喜欢
旅游纵览(2023年2期)2023-03-16 09:34:26
学与玩(2022年9期)2022-10-31 02:54:08
湖南大学学报(自然科学版)(2022年8期)2022-09-02 04:22:42
中外文摘(2022年12期)2022-08-02 08:36:08
大众摄影(2021年3期)2021-03-22 02:45:43
文苑(2020年12期)2020-04-13 00:54:14
小太阳画报(2019年1期)2019-06-11 10:29:48
意林(2019年7期)2019-04-28 14:38:50
作文评点报·高中版(2017年39期)2017-10-30 18:41:25
小学生必读(低年级版)(2017年5期)2017-08-12 03:47:07
