
小麦-长穗偃麦草T7BS·7EL易位系鉴定及7EL小片段易位诱导
2018-06-13 02:12:24李海凤罗贤磊段亚梅张璐璐陈建民
麦类作物学报 2018年5期
李海凤,罗贤磊,段亚梅,戴 毅,高 勇,张 军,张璐璐,陈建民
(1.扬州大学生物科学与技术学院,江苏扬州 225009; 2.扬州市职业大学,江苏扬州 225012)
小麦赤霉病(Fusarium head blight,FHB)是由禾谷镰刀菌(Fusariumgraminearum)等真菌侵染所造成的生育后期的气候性病害,在温暖湿润和半湿润地区(如中国、日本、南美等国)危害尤其严重。该病不仅导致小麦产量损失,而且籽粒中由赤霉菌产生的毒素脱氧雪腐镰刀菌烯醇(DON)会危害人畜健康。由于普通小麦内源的抗赤霉病基因资源较少,其抗性还不能满足抗赤霉病育种的需要。因此,在小麦近缘种属中挖掘新的赤霉病抗源并将其导入小麦尤显重要。
长穗偃麦草(Thinopyrumelongatum,syn.Agropyronelongatum,syn.Elytrigiaelongata)是小麦的近缘野生种,属于小麦的三级基因源,具有抗锈病、白粉病、条纹花叶病、赤霉病等多种抗性,亦具有大穗多花、抗寒耐旱、耐盐碱和籽粒蛋白质含量高等诸多优良性状[1-3]。在自然界中,长穗偃麦草有二倍体、四倍体和十倍体3种不同的倍性类型。一般认为二倍体长穗偃麦草[Thinopyrumelongatum(Host)A. Löve,2n=2x=14,EE或EeEe]的E染色体是长穗偃麦草多倍体物种的基本染色体组之一[4-5]。研究表明,二倍体长穗偃麦草1E和7E染色体上携带有赤霉病抗性基因[6-10]。Dvorak[11]利用二倍体长穗偃麦草育成了整套中国春-长穗偃麦草附加系和代换系。由于带有完整外源染色体的附加系和代换系的遗传稳定性和农艺性状差,难以在生产上直接利用,因此培育遗传稳定及农艺性状优良的小麦-长穗偃麦草易位系对于小麦抗赤霉病育种具有现实意义。Knott等[12-14]通过诱导小麦7D染色体与十倍体长穗偃麦草的7el2染色体间部分同源重组获得易位系KS10-2(7el2S·7el2L-7DL)和KS24-1(7DS·7el2L),将长穗偃麦草携带的抗秆锈病及抗赤霉病基因转入小麦;随后,Shen和Zhang等[15-16]报道十倍体长穗偃麦草7el2染色体长臂携有抗赤霉病基因Fhblop。为了将二倍体长穗偃麦草7E染色体抗赤霉病基因转入栽培小麦,张璐璐等[17]用60Co-γ射线(剂量30 000 rad)照射[中国春-二倍体长穗偃麦草7E代换系DS7E(7B)×扬麦16]的F2代种子,自M1至M4通过抗赤霉病接种和染色体特异分子标记鉴定,从M4代中获得小麦-长穗偃麦草7EL抗赤霉病易位系TW-7EL2。本研究以TW-7EL2为材料,利用荧光原位杂交、分子标记技术和赤霉病抗性评价,继续对该易位系进行染色体身份及抗性鉴定。携带目标基因的小片段易位染色体遗传稳定性更好,可有效减少外源染色体带来的冗余基因,是转移外源有利基因的理想材料,因此,本研究同时利用60Co-γ射线辐射处理该易位系的花粉,诱导产生长穗偃麦草染色体7EL的不同片段大小的结构变异材料,以期为赤霉病抗性基因定位和向小麦导入长穗偃麦草有益基因奠定基础。
1 材料与方法
1.1 试验材料
小麦-长穗偃麦草7EL易位系TW-7EL2、小麦-长穗偃麦草7E短臂附加系W-DA7ES,由扬州大学提供;二倍体长穗偃麦草,中国春-长穗偃麦草二体附加系DA7E,7E端二体附加系DA7EL、DA7ES,中国春-长穗偃麦草二体代换系DS7E(7A)、DS7E(7B)和中国春,由加拿大农业部Ottawa研究中心惠赠;安农8455、苏麦3号和扬麦16,由江苏里下河地区农业科学研究所惠赠;中国春缺-四体材料N7AT7B、N7BT7A和N7DT7A,由山东农业大学惠赠。
1.2 试验方法
1.2.1 花粉辐射及辐射后代处理
将小麦-长穗偃麦草7EL纯合易位系TW-7EL2即将开花的穗子(中部小穗基部小花的花药呈微黄色的穗子)从茎基部剪下,置于盛有清水的三角瓶中,用60Co-γ 射线照射处理,辐射剂量1 200 rad,剂量率为100 rad·min-1。取辐射处理当天收集的新鲜花粉授予已去雄的小麦品种扬麦158,收获杂交种M1。利用基因组原位杂交(GISH)技术检测M1植株中7EL染色体的结构变异体,统计外源染色体小片段顶端易位、小片段中间插入易位以及7EL顶端缺失的M1植株数。从每株M1变异材料中选取2~3个穗子去雄后与扬麦158回交,其余穗子套袋自交获得后代种子。
1.2.2 细胞学鉴定
易位系根尖体细胞有丝分裂中期染色体制片参照Gill等[18]的方法,在相差显微镜下进行染色体计数、观察,放入-70℃超低温冰箱中冷冻后揭去盖玻片,在70%、95%、100% 乙醇中梯度脱水后进行荧光原位杂交(fluorescent in situ hybridization,FISH)。双色荧光原位杂交参照Zhang等[19]的方法,以Fluorescein-12-dUTP标记的长穗偃麦草基因组DNA和以 Digoxinin-11-dUTP 标记的质粒 pSc119.2 为探针进行荧光原位杂交,以中国春的基因组DNA为封阻[23-24]。杂交洗涤后用DAPI(4,6-diamidino-2-phenylindole,DAPI)染色,每张载玻片加10 μL DAPI(含抗褪变剂)复染,盖上盖玻片。在Nikon(Ni-U)荧光显微镜下进行观察,尼康DS-U3数码相机摄取图像。
1.2.3 分子标记鉴定
参照Sharp等[20]的方法提取供试材料的DNA,利用长穗偃麦草染色体7E短臂特异标记P7E_No.39[21]、7E长臂特异标记Xgwm333[7,22]对易位系及诱导后代进行分析。扩增反应体系与程序参见Taq酶(TaKaRa,Japan)说明书,其中退火温度50~60 ℃,延伸时间1 min 。扩增产物用 1%琼脂糖凝胶电泳进行检测,在凝胶成像系统中观察、照相。
1.2.4 赤霉病抗性鉴定
于2015-2017年在扬州大学试验田采用单花滴注接种禾谷镰刀菌(菌种为本地强毒株的分生孢子混合液,由江苏省里下河地区农业科学研究所惠赠)对供试材料进行抗病性鉴定,每份材料接种 10~20穗,接种 21 d后,调查接种穗的发病小穗数和总小穗数。病小穗率=病小穗数/总小穗数×100%。
2 结果与分析
2.1 易位系的细胞学鉴定结果
双色荧光原位杂交结果显示,一对易位染色体的长穗偃麦草部分有绿色弥散信号,并且在其端部显示出pSc119.2的较强红色信号,与7EL长臂相似,进一步证实易位染色体中长穗偃麦草片段为7E长臂(图1A);在易位染色体中小麦片段端部显示出较强的pSc119.2杂交信号,与染色体7B短臂的杂交信号相似(图1B)。由此推测该易位染色体是小麦7B短臂和长穗偃麦草7E染色体长臂的整臂易位。
花粉母细胞(PMC)减数分裂中期I染色体构型分析表明,一对易位染色体在绝大多数PMC中均能配对成环状二价体(图1C)。在所观察的35个花粉母细胞中,平均每个花粉母细胞中含0.23个单价体,1.74个棒状二价体,19.06个环状二价体,0.11个三价体,三价体并不涉及小麦与长穗偃麦草的易位。上述结果表明该易位系具有较好的细胞学稳定性。
2.2 易位系的分子标记鉴定结果
利用标记P7E_No.39和Xgwm333对中国春、扬麦16、二倍体长穗偃麦草等材料进行PCR扩增,结果表明,标记P7E_No.39在易位系TW-7EL2 中未能扩增出约330 bp的7ES特异条带(图2),而标记Xgwm333能扩增出约130 bp的7EL特异条带(图3),说明该易位染色体长穗偃麦草部分来自7EL。利用标记Xgwm333对中国春、扬麦16、中国春的第七同源群缺-四体、长穗偃麦草的7E附加系、代换系和易位系进行分析,结果表明,中国春缺体7B-四体7A(N7BT7A)、小麦-长穗偃麦草代换系DS7E(7B)和易位系TW-7EL2均缺失约170 bp的特异条带,而代换系DS7E(7B)与易位系TW-7EL2的带型一致,说明该易位系中缺失小麦的7BL染色体。综合细胞学研究结果,该易位系的易位染色体涉及小麦7BS和长穗偃麦草7EL染色体,命名为T7BS·7EL易位。
2.3 易位系赤霉病抗性鉴定
连续3年的赤霉病接种鉴定结果表明,易位系T7BS·7EL的赤霉病抗性显著高于感病亲本中国春及感病对照安农8455,也显著优于中抗赤霉病亲本扬麦16,与抗病对照苏麦3号差异不显著(表1)。穗部发病程度见图4。
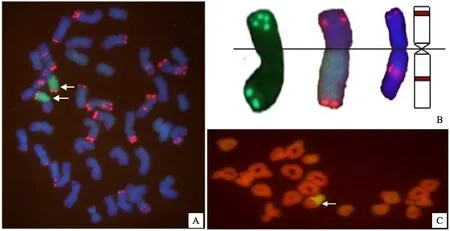
A:绿色为长穗偃麦草基因组DNA杂交信号,红色为pSc119.2杂交信号,箭头示易位染色体; B:染色体7E,T7BS·7EL和7B的荧光原位杂交及模式图,点状绿色和红色信号为pSc119.2杂交信号; C:易位系TW-7EL2 减数分裂中期I染色体的荧光原位杂交,小麦染色体呈红色,长穗偃麦草染色体呈黄绿色; 箭头示配成环状二价体的易位染色体T7BS·7EL。
In panel A,Th.elongatumgenomic DNA was visualized with green signals,and pSc119.2 was visualized with red signals; the arrows show translocation chromosomes. In panel B,from the left to right:FISH patterns of chromosome 7E,the translocation chromosome T7BS·7EL and chromosome 7B; pSc119.2 was visualized with green and red spot signals. In panel C,pollen mother cells at MI of the translocation line TW-7EL2; wheat chromosomes display red color,andTh.elongatumchromosome segments show yellow-green color; arrow shows the ring bivalent formed by a pair of T7BS·7EL.
图1易位系TW-7EL2的染色体荧光原位杂交结果(2n=42)
Fig.1ImagesofchromosomesinthetranslocationlineTW-7EL2(2n=42)detectedbyFISH
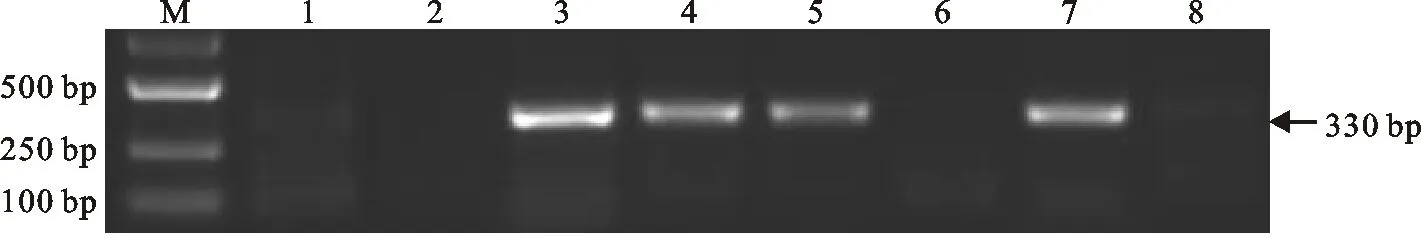
M:Marker 2501; 1:中国春; 2:扬麦16; 3:二倍体长穗偃麦草; 4:中国春-长穗偃麦草附加系DA7E; 5:7E短臂附加系; 6:7E长臂附加系; 7:中国春-长穗偃麦草代换系DS7E(7B); 8:易位系T7BS·7EL。
M:Marker 2501; 1:Chinese Spring; 2:Yangmai 16; 3:Th.elongatum; 4:DA7E; 5:DA7ES; 6:DA7EL; 7:DS7E(7B); 8:T7BS·7EL.
图2标记P7E_No.39(7ES)的扩增结果
Fig.2ProductsamplifiedwithmarkerofP7E_No.39(7ES)
2.4 长穗偃麦草7EL小片段结构变异体的创制
本研究共得到M1植株96株,对这些材料进行根尖细胞有丝分裂中期染色体GISH鉴定,共检测到含有7EL染色体小片段结构变异的单株15株,占M1观察植株数的15.62%。除6株只含有单个小片段中间插入或者顶端易位外(图5A、图5B),其余9株细胞内同时具有两种7EL染色体小片段结构变异,包括不同长度的小片段顶端易位、小片段中间插入易位染色体和7EL顶端缺失染色体(图5C、图5D)。在获得的15株材料中,有2株在抽穗前死亡,其余13株自交结实率差,均通过与扬麦158回交获得回交种子,使易位染色体得以传递下来。分离各种类型易位系的纯合体正在进行中。
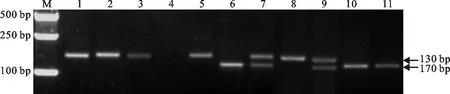
M:Marker 2501; 1:中国春; 2:扬麦16; 3:缺体7A四体7B; 4:缺体7B四体7A; 5:缺体7D四体7A; 6:二倍体长穗偃麦草; 7:中国春-长穗偃麦草附加系DA7E; 8:7E短臂附加系; 9:7E长臂附加系; 10:中国春-长穗偃麦草代换系DS7E(7B); 11:易位系T7BS·7EL。
M:Marker 2501;1:Chinese Spring; 2:Yangmai 16; 3:N7AT7B; 4:N7BT7A; 5:N7DT7A; 6:Th.elongatum; 7:DA7E; 8:DA7ES; 9:DA7EL; 10:DS7E(7B); 11:T7BS·7EL.
图3标记Xgwm333(7BL,7EL)的扩增结果
Fig.3ProductsamplifiedwithmarkerofXgwm333(7BL,7EL)
A:易位系7BS·7EL; B:7ES端体附加系; C:中国春; D:扬麦16; E:苏麦3号; F:安农8455。
A:T7BS·7EL; B:W-DA7ES; C:Chinese Spring; D:Yangmai 16; E:Sumai 3; F:Annong 8455.
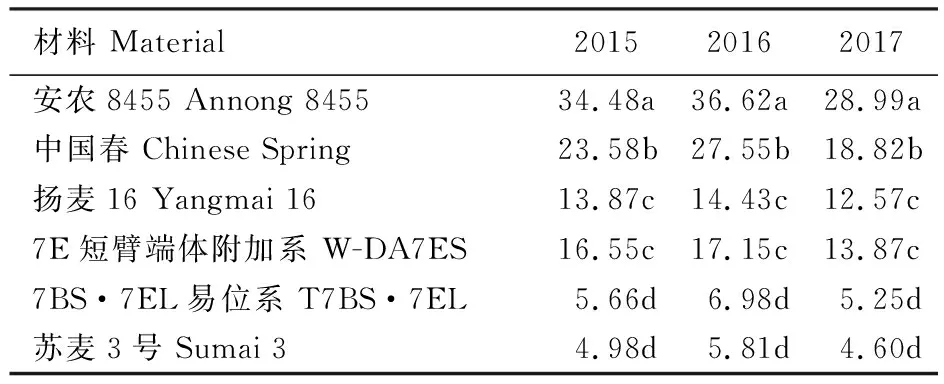
图4 供试材料的赤霉病抗性鉴定结果
表中数据为3年赤霉病病小穗率;同一列数据后的不同小写字母表示在0.05 水平上有显著差异。
The data in the table are the spikelet rate of scab for 3 years; Different letters following values indicate significant difference among lines at 0.05 level.
3 讨 论
Kibridge-Sebuny等[13]报道,利用中国春ph1b突变体诱导小麦7D染色体与十倍体长穗偃麦草的7el2染色体间部分同源重组,选育到易位系7el2S·7el2L-7DL(KS10-2)和7DS·7el2L(KS24-1)[14]。目前,这两份易位系已被作为中间材料将7el2的抗赤霉病基因和抗秆锈病基因 Sr43转入硬粒小麦和普通小麦中[27-28]。Guo等[25]利用中国春ph1b突变体与7DS·7el2L整臂易位杂交,从后代中筛选出一个约含有1/3长穗偃麦草7el2长臂的小片段易位系,将抗赤霉病基因 Fhb7定位于7el2L上,并获得聚合 Fhb1与 Fhb7抗病基因的小麦抗赤霉病种质。在已报道的小麦-长穗偃麦草代换系和易位系中,来源于十倍体长穗偃麦草的一对部分同源染色体7el1和7el2,与二倍体长穗偃麦草7E染色体具有部分同源性,7E与7el1间配对频率为13.6%,而7el1和7el2染色体间配对频率为71.64%[26]。赤霉病抗性研究表明,7el1的代换系和易位系均没有抗性,而整臂易位7DS·7el2L具有抗性[7]。二倍体长穗偃麦草的E组染色体是长穗偃麦草的基本染色体组之一,深入开展二倍体长穗偃麦草的研究可以推动和加速偃麦草属基因的开发和利用。张璐璐等[17]用60Co-γ射线照射DS7E(7B)×扬麦16的F2代种子,从M4代中成功获得小麦-长穗偃麦草7EL抗赤霉病易位系。在此工作基础上,本研究利用双色GISH-FISH结合分子标记对易位系TW-7EL2进行鉴定,表明该易位系为涉及小麦7BS和长穗偃麦草7EL的整臂易位,遗传稳定性好。赤霉病抗性鉴定表明,该易位系具有较高的赤霉病抗性,是小麦抗赤霉病育种又一新的种质材料。
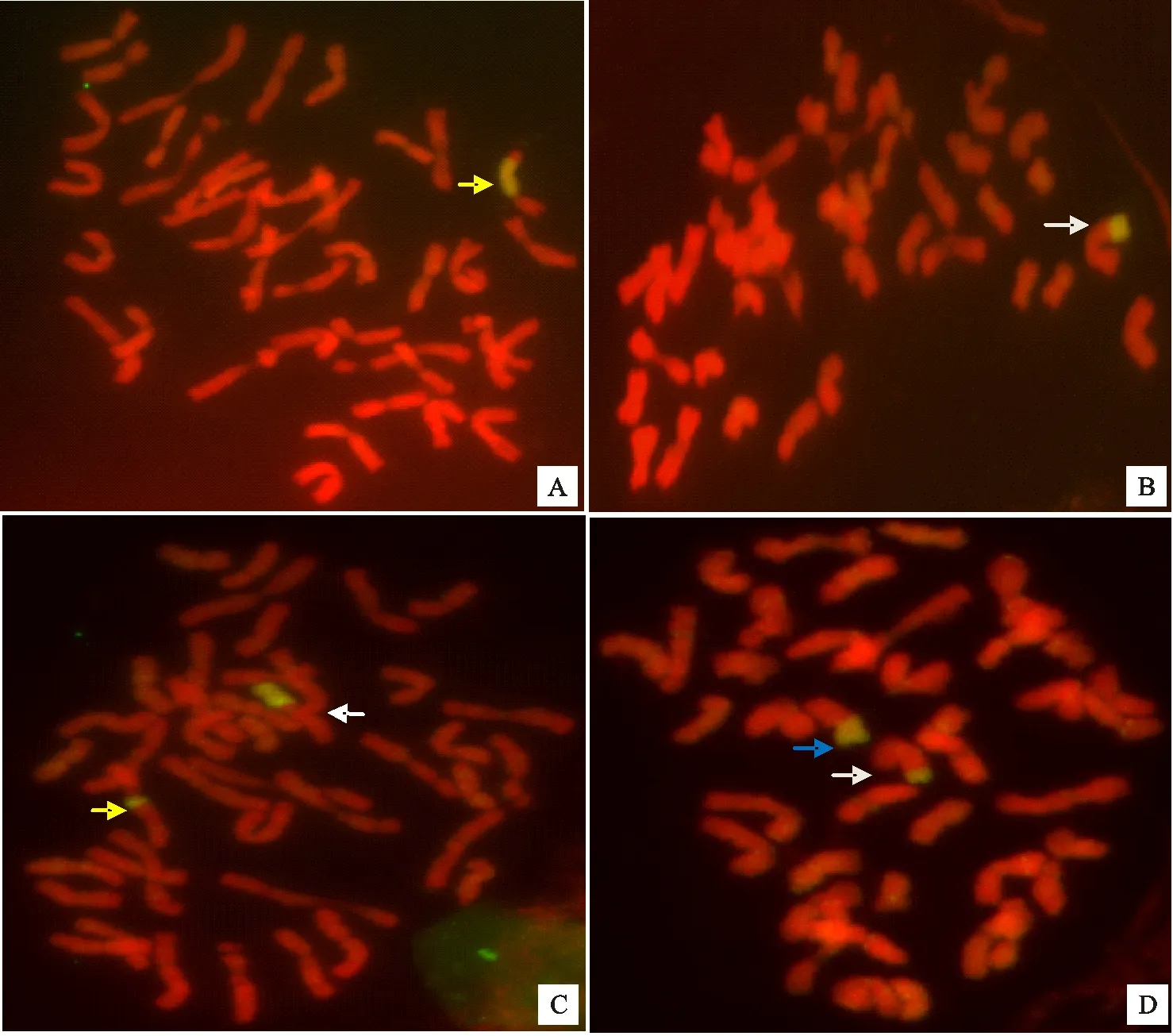
A:箭头所指为一条中间插入易位染色体; B:箭头所指为一条顶端易位染色体; C:白色箭头所指为一条顶端易位染色体,黄色箭头所指为一条中间插入易位染色体; D:白色箭头所指为小片段顶端易位染色体,蓝色箭头为顶端缺失染色体。小麦染色体呈红色,长穗偃麦草染色体呈黄绿色。以二倍体长穗偃麦草基因组DNA作探针,中国春基因组DNA为封阻进行GISH。
A:The arrow shows an interstitial chromosome with small segment; B:The arrow shows a small segment terminal translocation chromosome; C:The white arrow shows a small segment terminal translocation chromosome; the yellow arrow shows an interstitial chromosome with different segment of chromosome arm 7EL; D:The white arrow shows a small segment terminal translocation chromosome; the blue arrow shows a deletion chromosome of chromosome arm 7EL. Wheat chromosomes display red color,andTh.elongatumchromosome segments show yellow-green color. Total genomic DNA ofTh.elongatum(2X) was labeled as probe and Chinese spring genomic DNA as block for GISH.
图5含有7EL结构变异染色体的M1植株根尖细胞有丝分裂中期染色体GISH
Fig.5GISHpatternsofchromosomeaberrationsinvolvingthelongarmofchromosome7Eof
Th.elongatumatthemitoticmetaphaseintheroot-tipcellsofM1plants
小麦异源易位系在小麦遗传改良中具有重要的意义,培育携有外源目的基因的片段尽可能小的易位系在小麦育种中利用价值较高。电离辐射是一种有效诱导染色体结构变异的方法[29]。陈升位等[30]利用60Co-γ 射线处理T6AL·6VS整臂易位系的成熟雌配子,从534株M1材料中检测到97株涉及6VS染色体的小片段结构变异植株,诱变频率为18.16%。Zhao等[31]利用60Co-γ 射线处理T4DL·4VS纯合易位系成熟花粉,从266株M1材料中检测到含有4VS染色体小片段结构变异的单株40株,诱变频率为15.04%。本研究利用60Co-γ射线处理T7BS·7EL纯合易位系即将开花的穗子,从96株M1材料中检测到含有7EL染色体的小片段结构变异的单株15株,占M1观察植株数的15.62%,表明通过电离辐射处理小麦-长穗偃麦草易位系的成熟花粉,可以快速获得较多类型的变异体,达到较高的诱变效应。以整臂易位系作为辐射材料,由于仅涉及外源染色体的单条臂,针对性强,外源染色体臂发生一次断裂即可产生小片段中间易位或顶端易位,或者形成缺失。但是,由于电离辐射在诱发易位的同时也会引起小麦染色体本身存在较多变异,导致变异植株难以结实或后代种子不发芽、夭亡或不育,因此,为了解决辐射杂种M1代中的易位染色体通过自交传递率低的问题,本研究中利用含有结构变异染色体的植株做母本与小麦品种继续回交,使得M1代中具有结构变异的染色体通过雌配子得以传递,降低了后代中的结构变异染色体数量,在后代选择中再利用分子标记选择,保留具有长穗偃麦草染色体的植株,随着自交世代的增加,可获得结实正常的长穗偃麦草染色体小片段易位系。目前,选育涉及染色体7EL的小片段纯合易位系的工作及赤霉病抗性鉴定正在进行中,有望获得不同类型的变异体用于抗赤霉病基因定位和分子标记物理作图。
参考文献:
[1] DVORAK J,EDGE M,ROSS K.On the evolution of the adaptation ofLophopyrumelongatumto growth in saline environments [J].ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica,1988,85(11):3805.
[2] ZHONG G Y,DVORAK J.Chromosomal control of the tolerance of gradually and suddenly imposed salt stress in theLophopyrumelongatumand wheat,TriticumaestivumL.genomes [J].TheoreticalandAppliedGenetics,1995,90(2):229.
[3] LI Z S,HAO S.Chromosome engineering of wheat in China [J].CriticalReviewsinPlantSciences,1992,10(5):471.
[4] 董玉琛,郑殿生.中国小麦遗传资源 [M].北京:中国农业出版社,2000:183.
DONG Y C,ZHENG D S.Genetic resource ofTriticeaein China [M].Beijing:China Agricultural Press,2000:183.
[5] 唐朝晖,刘少翔,张兰萍,等.二倍体长穗偃麦草E组染色体研究进展 [J].山西农业科学,2007,35(5):3.
TANG Z H,LIU S X,ZHANG L P,etal.Advances in research on E genome ofThinopyrumelongatum[J].JournalofShanxiAgriculturalSciences,2007,35(5):3.
[6] 刘登才,郑有良,王志容,等.影响小麦赤霉病抗性的Lophopyrumelongatum染色体定位 [J].四川农业大学学报,2001,19(3):200.
LIU D C,ZHENG Y L,WANG Z R,etal.Distribution of chromosomes in diploidLophopyrumelongatum(Host) A.Löve that influences resistance to head scab of common wheat [J].JournalofSichuanAgriculturalUniversity,2001,19(3):200.
[7] SHEN X R,Kong L R,OHM H.Fusarium head blight resistance in hexaploid wheat(Triticumaestivum)-Lophopyrumgenetic lines and tagging of the alien chromatin by PCR markers [J].TheoreticalandAppliedGenetics,2004,108:808.
[8] WANG J R,WANG L,GULDEN S,etal.RNA profiling of Fusarium head blight-resistant wheat addition lines containing theThinopyrumelongatumchromosome 7E [J].CanadianJournalofPlantPatholog,2010,32(2):188.
[9] JAUHAR P P,PETERSON T S,XU S S.Cytogenetic and molecular characterization of a durum alien disomic addition line with enhanced tolerance to Fusarium head blight [J].Genome,2009,52(5):467.
[10] 陈士强,黄泽峰,张 勇,等.中国春背景下长穗偃麦草抗赤霉病相关基因的染色体定位 [J].麦类作物学报,2012,32(5):839.
CHEN S Q,HUANG Z F,ZHANG Y,etal.Chromosomal location of the genes associated with FHB resistance ofLophopyrumelongatumin Chinese Spring background [J].JournalofTriticeaeCrops,2012,32(5):839.
[11] DVORAK J.Disomic and ditelosomic additions of diploidAgropyronelongatumchromosomes toTriticumaestivum[J].Genome,1974,16(2):399.
[12] KNOTT D R,DVORAK J,NANDA J S.The transfer to wheat and homoeology of anAgropyronelongatumchromosome carrying resistance to stem rust [J].Genome,1977,19(1):75.
[13] KIBIRIDGE-SEBUNYA I,KNOTT D R.Transfer of stem rust resistance to wheat from anAgropyronchromosome having a gametocidal effect [J].Genome,1983,25(3):215.
[14] KIM N S,ARMSTRONG K,KNOTT D R.Molecular detection ofLophopyrumchromatin in wheat-Lophopyrumrecombinants and their use in the physical mapping of chromosome 7D [J].TheoreticalandAppliedGenetics,1993,85(5):561.
[15] SHEN X R,OHM H.Fusarium head blight resistance derived fromLophopyrumelongatumchromosome 7E and its augmentation with Fhb1 in wheat [J].PlantBreeding,2006,125(5):424.
[16] ZHANG X L,SHEN X R,HAO Y F,etal.A genetic map ofLophopyrumponticumchromosome 7E,harboring resistance genes to Fusarium head blight and leaf rust [J].TheoreticalandAppliedGenetics,2011,122(2):263.
[17] 张璐璐,陈士强,李海凤,等.小麦-长穗偃麦草7E抗赤霉病易位系培育 [J].中国农业科学,2016,49(18):3477.
ZHANG L L,CHEN S Q,LI H F,etal.Development of wheat-Thinopyrumelongatumtranslocation lines resistant to Fusarium head blight [J].ScientiaAgriculturaSinica,2016,49(18):3477.
[18] GILL B S,FRIEBE B,ENDO T R.Standard karyotype and nomenclature system for description of chromosome bands and structural aberration in wheat [J].Genome,1991,34(5):830.
[19] ZHANG P,LI W,FRIEBE B,etal.Simultaneous painting of three genomes in hexaploid wheat by BAC-FISH [J].Genome,2004,47(5):979.
[20] SHARP P J,CAO S,DESAI S,etal.The isolation,characterization and application in theTriticaleof a set of wheat RFLP probes identifying each homoeologous chromosome arm [J].TheoreticalandAppliedGenetics,1989,78(3):342.
[21] CHEN S Q,HUANG Z F,DAI Y,etal.The development of 7E chromosome-specific molecular markers forThinopyrumelongatumbased on SLAF-seq technology [J].PLoSOne,2013,8(6):e65122.
[22] SOMERS D J,ISAAC P,EDWARD K.A high-density microsatellite consensus map for bread wheat(TriticumaestivumL.) [J].TheoreticalandAppliedGenetics,2004,109(6):1105.
[23] MUKAI Y,NAKAHARA Y,YAMAMOTO M.Simultaneous discrimination of the three genomes in hexaploid wheat by multicolor fluorescence in situ hybridization using total genomic and highly repeated DNA probes [J].Genome,1993,36(3):489.
[24] LINC G,SEPSI A,MOLNAR-LANG M.A FISH karyotype to study chromosome polymorphisms for theElytrigiaelongateE genome [J].CytogeneticGenomeResearch,2012,136(2):138.
[25] GUO J,ZHANG X L,HOU Y,etal.High-density mapping of the major FHB resistance gene Fhb7 derived fromThinopyrumponticumand its pyramiding with Fhb1 by marker-assisted selection [J].TheoreticalandAppliedGenetics,2015,128(11):2301.
[26] DVORAK J.Meiotic pairing between single chromosomes of diploidAgropyrumelongatumand decaploidA.elongatuminTriticumaestivum[J].CanadianJournalofGeneticsandCytology,1975,17(3):329.
[27] FORTE P,VIRILI M E,KUZMANOVIC L,etal.A novel assembly ofThinopyrumponticumgenes into the durum wheat genome:Pyramiding lines previously engineered for other beneficial traits from the same alien species [J].MolecularBreeding,2014,34(4):1701.
[28] NIU Z,KLINDWORTH D L,YU G,etal.Development and characterization of wheat lines carrying stem rust resistance gene Sr43 derived fromThinopyrumponticum[J].TheoreticalandAppliedGenetics,2014,127:969.
[29] BIE T D,CAO Y P,CHEN P D.Mass production of intergeneric chromosomal translocations through pollen irradiation ofTriticumdurum-Haynaldiavillosaamphiploid [J].JournalofIntegrativePlantBiology,2007,49(11):1619.
[30] 陈升位,陈佩度,王秀娥.利用电离辐射处理整臂易位系成熟雌配子诱导外源染色体小片段易位[J].中国科学C辑(生命科学),2008,38(3):346.
CHEN S W,CHEN P D,WANG X E.Inducement of chromosome translocation with small alien segments by irradiating mature female gametes of the whole arm translocation line [J].ScienceChinaLifeScienece,2008,51(4):346.
[31] ZHAO R H,WANG H Y,JIA Q,etal.Development of EST-PCR Markers for the chromosome 4V ofHaynaldiavillosaand their application in identification of 4V chromosome structural aberrant [J].JournalofIntegrativeAgriculture,2014,13(2):282.
猜你喜欢
中国卫生标准管理(2022年21期)2023-01-03 02:36:34
作物学报(2022年2期)2022-11-06 12:08:56
快乐语文(2020年14期)2020-07-04 00:02:00
江苏农业科学(2017年16期)2017-10-27 12:48:11
现代农业科技(2017年8期)2017-06-10 09:32:18
西部(2016年6期)2016-05-14 19:09:59
医学研究杂志(2015年2期)2015-06-10 06:45:00
中国果业信息(2015年6期)2015-01-23 17:38:07
河南科技(2014年15期)2014-02-27 14:12:50
俄罗斯问题研究(2012年1期)2012-03-25 09:54:41
