
夏玉米对土壤水分持续减少的响应及其转折点阈值分析
2018-06-07 02:58石耀辉周广胜王秋玲麻雪艳冯晓钰
生态学报 2018年8期
石耀辉,周广胜,王秋玲,麻雪艳,冯晓钰
中国气象科学研究院,北京 100081
玉米是世界上种植最广泛的粮食作物之一,其总产量居三大粮食作物(水稻、小麦和玉米)之首。中国是世界上第二大玉米生产大国,占世界总产量的20%,玉米在保障中国粮食安全和国民经济发展方面发挥着重要作用[1-4]。中国有 2/3 的玉米分布在依靠自然降水的旱地上,降水是影响玉米生产的重要环境因素之一,严重干旱年份可造成部分地区绝收[2,5-9]。全球变化导致的温度升高会诱发降水格局改变,半个世纪以来,中国年降水量变化总趋势是逐渐减少,其中夏季减少最多[2,10-11]。1981—2010年的 30年间,中国夏玉米主产区的旱灾受灾面积占4种气象灾害(旱灾、涝灾、风暴和冷害)总受灾面积的64%,干旱是影响夏玉米的主要气象灾害[9]。未来气候变化情景下,干旱和高温等极端天气气候事件呈频发并有加剧的趋势,将威胁到作物产量和粮食安全,中国北方地区温度的升高会使得水资源短缺加剧,夏玉米受干旱影响可能会更严重,玉米旱灾的应对越来越受到关注[11-15]。
干旱指标是确定干旱是否发生及评估其严重程度的标准,是干旱监测预警的基础。植物生理生态指标是植物耐旱能力的具体体现,直接反映植物受旱程度,主要包括植物形态指标和植物生理指标两类[1,3,16-17]。干旱胁迫下叶片形态变化比较明显,单叶叶面积和叶数降低,植株总叶面积减小,并且叶片常常发生卷曲萎蔫,导致光合有效叶面积降低,进而影响作物产量的形成[18-22]。干旱胁迫下玉米节间缩短,株高降低,茎杆变细[16,18,22-26]。随着水分减少,多数植物比叶面积减小,叶片增厚[20,27-28]。在受到干旱胁迫时叶片含水量显著下降[28-29]。叶片缺水后,会影响叶绿素的生物合成,并促进已有叶绿素的加速分解,从而导致叶片发黄,影响叶片的光合作用,进而影响作物产量[8,29-30]。Siddique等[27]和刘京宝等[16]报道,叶水势是反映植物对水分胁迫响应较好的指标;Lobel等[7]提出,植物饱和水汽压差(VPD)可以较敏感地反映玉米对干旱胁迫的响应。这些研究为认识植物对干旱的响应提供了支持。
为预防和减缓干旱对玉米生产带来的不良影响,需要及时准确地获取作物干旱的信息[3]。干旱是一个逐步发展的动态过程,同时又是一个长时间的累积过程,受持续时间、土壤水分传输等一系列因素影响,静态的土壤含水量所包含的干旱的累积信息少[3,31]。玉米在整个生育期对水分都十分敏感,动态评估干旱发生时间,准确反映干旱累积效应,提高对干旱的预测精度和及时性,对于玉米干旱研究有着重要意义[2-3]。本研究以夏玉米为研究材料,基于土壤水分持续减少模拟试验,尝试定量评估夏玉米生理特征、生物量积累与形态特征对土壤干旱过程的响应,为客观辨识夏玉米干旱的发生发展及监测预警提供参考。
1 材料与方法
1.1 试验材料和环境条件
以夏玉米郑单958品种为供试材料,试验于2015年6—10月在中国气象局固城生态与农业气象试验站(39°08′N,115°40′E)开展。该区域属暖温带大陆性季风气候,年均气温12.2℃,年均降水量528mm,年平均日照时数2264h[32]。试验为小区控水模拟试验,每小区面积为8m2(2m×4m),各小区之间用3m深混凝土墙隔离以防止小区间水分相互渗透。试验场配有大型电动遮雨棚,降雨时启用,排除自然降水干扰。小区土壤类型为沙壤土,0—50cm平均土壤容重和田间持水量分别为1.37g/cm3和22.1%。
1.2 试验设计
试验设2个水分处理,(1) 充分供水(A):夏玉米全生育期维持土壤相对湿度为75%±5%;(2)土壤水分持续减少处理(D):夏玉米拔节期前充分供水维持土壤相对湿度为75%±5%,拔节期开始不再补水,模拟土壤水分持续减少对夏玉米的影响。每个处理3个重复,共6个试验小区。2015年6月25日播种,小区玉米行距40cm,株距30cm,每个小区约67株,播种后追施磷酸二铵300kg/hm2,每个小区240g。
1.3 测定内容与方法
1.3.1 土壤相对湿度
土壤相对湿度利用Diviner 2000便携式土壤水分仪观测,全生育期约每3天测定1次,测定0—50cm每10cm土壤分层含水量,对充分供水(土壤相对湿度75%±5%)小区进行灌溉补水。灌溉量(m3)依据实测各小区各层土壤水分含量(体积含水量θv)与参考土壤水分含量(体积含水量θref)的差值,采用式(1)计算,利用水表精确控制灌溉量。
ΔDw=A·∑(θref-θv)·h·ρ
(1)
式中,ΔDw是灌溉量;A是灌溉面积(m2);h是土层厚度(本试验每0.1m为1层);ρ为土壤容重。依据每次充分供水处理灌溉量ΔDw,采用公式(2)可以计算土壤水分持续减少处理组(D)相对于充分供水组(A)随时间的水分亏缺累积量(Dw)
(2)
1.3.2 指标观测
2015年7月18日夏玉米进入拔节期,土壤水分持续减少处理小区开始停止供水,记为干旱试验指标观测时间原点0天(d),之后每7—10d对各生理生态指标及生物量进行1次观测,每次每小区观测1株标准株。指标观测及方法如下。
(1) 植物器官生物量和含水量
生物量(g)采用烘干法进行测定,参照《农业气象观测规范》(1993),在9:00—11:00对样本植株破坏性取样。将叶,茎,根和穗(如果有)分离并标记,称取各部分鲜重(根除外),放入烘箱105℃杀青1h,之后在80℃下烘干至恒重,称取各部分干重(生物量),并计算样本各部分的含水量,含水量=(鲜重-干重)/鲜重。
(2) 形态特征
株高(cm)在样本植株取样前进行测定,抽雄期及以前,从土壤表面量至植株最高叶尖;抽雄期后,从土壤表面量至雄穗穗尖。叶数为出叶数,在取样后进行各器官分离时对所有可见叶计数。叶面积(cm2)在取样后采用面积系数法测定,用直尺量取样本植株每片完整绿色叶片长度(Li)和叶片最宽处宽度(Di),单株叶面积S用式(3)计算。单叶面积(cm2)=叶面积/叶数。比叶面积(cm2/g)=叶面积/叶生物量。
(3)
式中,k为校正系数,取值0.75。
(3) 叶绿素含量和叶水势
叶绿素含量采用SPAD 502叶绿素仪在样本植株取样前对其顶部第1片完全展开叶(Top- 1)和第3片叶(Top- 3)进行测定。该仪器测定值为相对值,无单位,每个叶片测定3次取平均值。叶水势(MPa)采用1505D-EXP型便携式植物水势气穴压力室进行测定,样本植株取样后,将顶部第1片完全展开叶片距基部1/3处剪下,沿主叶脉剪取略小于压力室叶室直径的样品,插入叶室内,通气并进行观测,记录当叶片样品边缘出现第1液滴时的压力值。
1.4 数据分析
数据分析采用SPSS 18.0分析软件进行, 在0.05水平上确定持续干旱对各指标影响的显著性。
2 结果与分析
2.1 土壤相对湿度
土壤相对湿度测定结果表明,夏玉米拔节期开始到试验结束,充分供水组(A)土壤相对湿度维持在75%±5%,土壤水分持续减少处理组(D)的土壤相对湿度随干旱持续时间呈持续下降状态,最终下降到约30%,两组处理呈现显著差异(P<0.001),达到预设控水效果。土壤水分持续减少处理组(D)的土壤相对湿度与干旱持续时间可用方程y=0.0045x2-0.896x+73.275较好的拟合(R2=0.94,P<0.001) (图1a)。依据每次充分供水组(A)灌溉量(mm),可以计算土壤水分持续减少处理组(D)相对于充分供水组(A)的水分亏缺累积量,水分亏缺累积量与处理时间呈较好的线性关系:y=3.2308x-1.7048 (R2=0.99,P<0.001) (图1)。
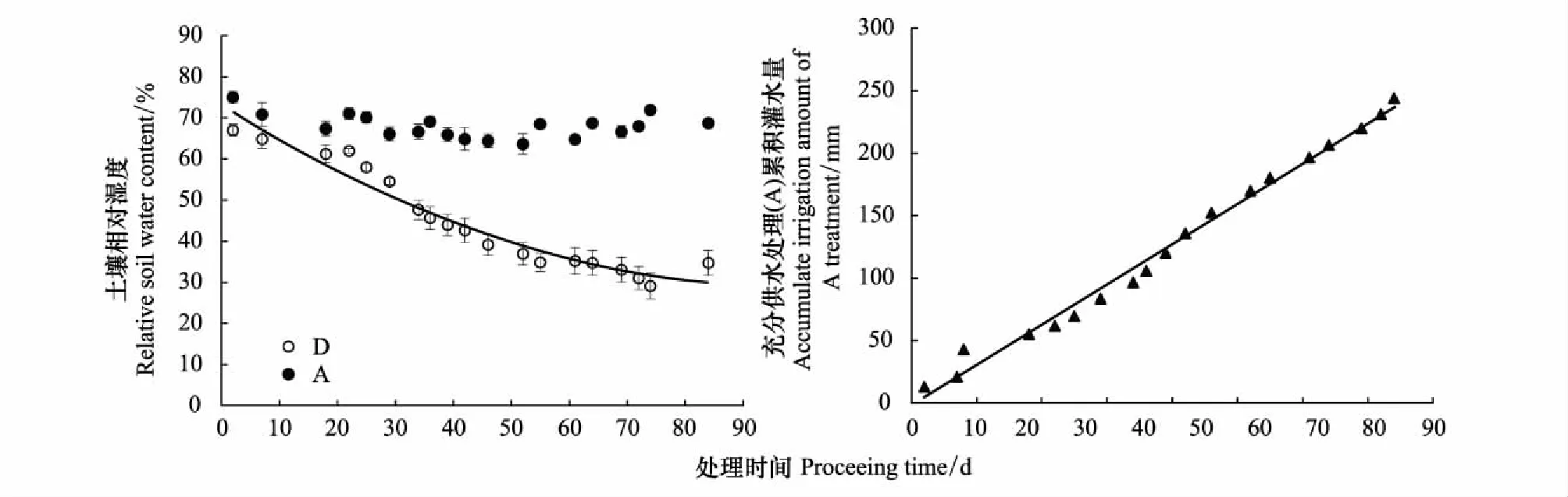
图1 两个处理组土壤相对湿度及水分差(灌水量累积)随时间的变化Fig.1 Variation of soil relative humidity and water deficit (irrigation accumulation) with time in two treatment groups
2.2 形态特征
土壤水分持续减少处理组(D)的株高、叶数、总叶面积和单叶面积在处理初期大于充分供水组(A),随着土壤水分持续减少,株高、叶数和总叶面积逐渐开始低于充分供水组(A) (图2),但单叶面积与充分供水组(A)差异逐渐减小(图2)。从整个试验期看,两个处理组的比叶面积差异没有达到显著水平,但土壤水分持续减少处理组(D)的比叶面积与充分供水组(A)相比在干旱胁迫后期呈现降低趋势(图2, 表1)。株高、叶数和总叶面积在土壤水分持续减少的前期和后期呈显著差异。两个处理组的株高、叶数和总叶面积与土壤水分减少的持续时间拟合的曲线交点是持续干旱造成不利影响开始点(图2, 表2),即形态指标随土壤水分持续减少受到胁迫时间阈值(d),分别为35、27d和38d。此时相对应的水分亏缺累积量即为指标开始受到胁迫时的水分亏缺阈值(mm),相对应的土壤相对湿度为指标开始受到胁迫时的土壤相对湿度阈值(%)。依据时间阈值,采用图1得到的干旱持续时间与水分亏缺累积量关系以及与土壤相对湿度的关系可以得出指标开始受到胁迫时的水分亏缺阈值和土壤相对湿度阈值,分别为111、86、121mm和47%、52%、46% (表2)。依据时间阈值的先后,水分亏缺阈值和土壤相对湿度阈值的大小,形态特征对土壤水分减少的敏感性顺序为叶数>株高>总叶面积。
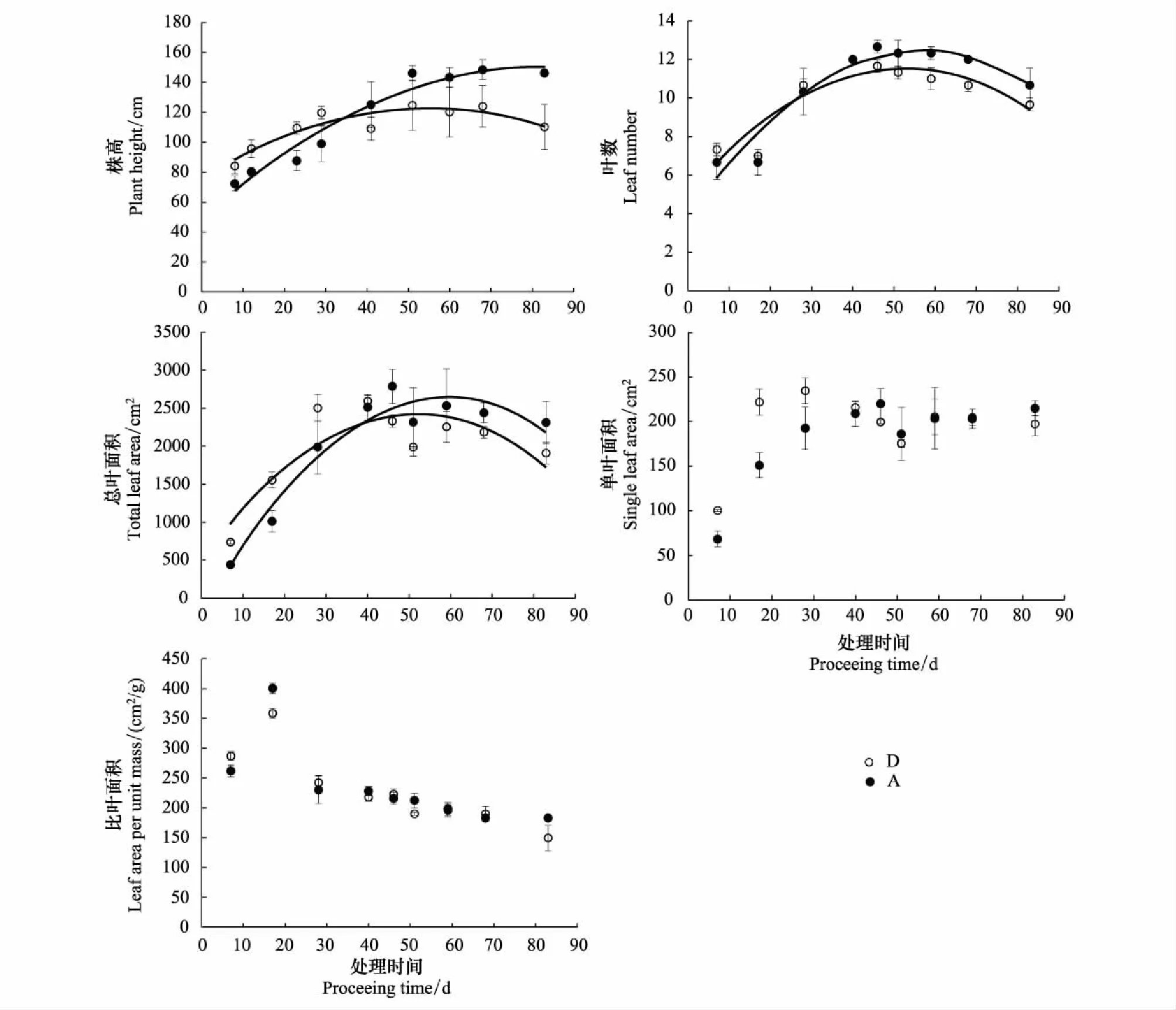
图2 土壤水分持续减少对形态特征的影响 (数据为平均值±标准误, n=3)Fig.2 The effects of consecutive soil water decrease on morphological characteristics (mean±SE, n=3)
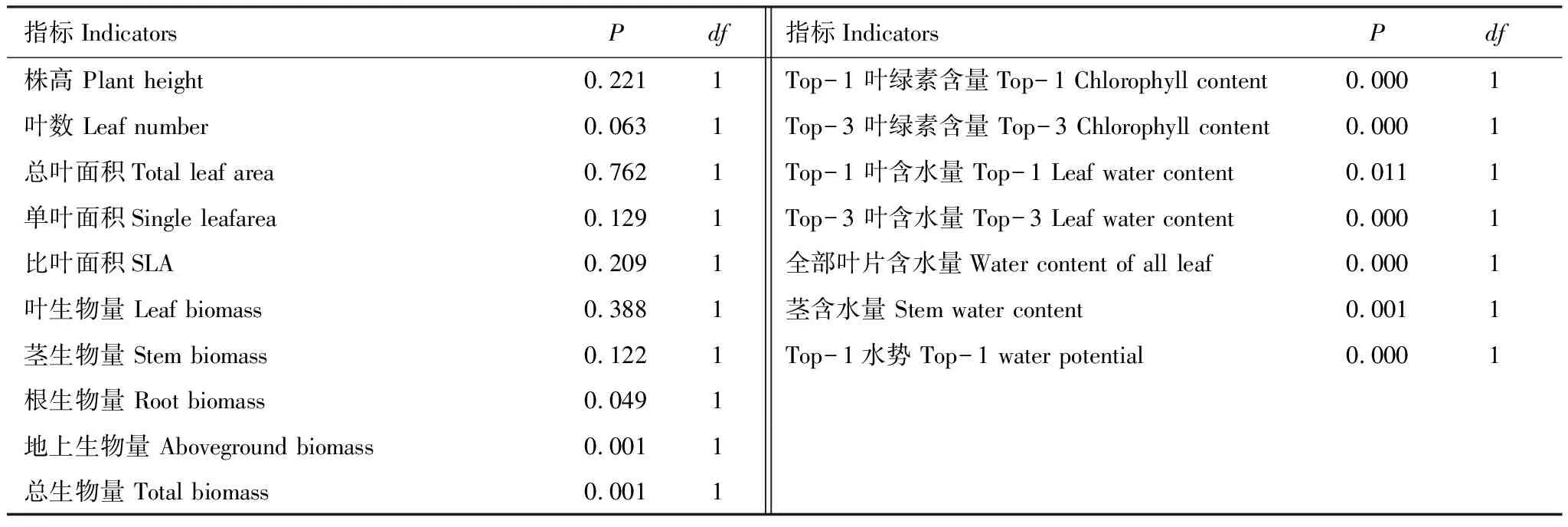
指标IndicatorsPdf指标IndicatorsPdf株高 Plant height0.2211Top-1 叶绿素含量Top-1 Chlorophyll content0.0001叶数 Leaf number0.0631Top-3 叶绿素含量 Top-3 Chlorophyll content0.0001总叶面积Total leaf area0.7621Top-1 叶含水量 Top-1 Leaf water content0.0111单叶面积Single leafarea0.1291Top-3 叶含水量 Top-3 Leaf water content0.0001比叶面积SLA0.2091全部叶片含水量Water content of all leaf0.0001叶生物量 Leaf biomass0.3881茎含水量 Stem water content0.0011茎生物量 Stem biomass0.1221Top-1水势 Top-1 water potential0.0001根生物量 Root biomass0.0491地上生物量 Aboveground biomass0.0011总生物量 Total biomass0.0011
表2夏玉米生理生态指标受土壤水分胁迫开始的时间(T),水分亏缺(WD)和土壤相对湿度(RSWC)的阈值
Table2Thresholdsofduration(T),Waterdeficit(WD)andRelativesoilwatercontent(RSWC)whensummermaizeeco-physiologicalindicatorswereaffectedbysoilwaterstress

指标Indicators水分处理Water treatment方程EquationsR2P阈值ThresholdT/dWD/mmRSWC/%形态特征Morphological characteristics株高 Plant heightAy=-0.015x2+2.469x+48.7740.96<0.013511147Dy=-0.0156x2+1.7197x + 75.6010.85<0.01叶数 Leaf numberAy=-0.0027x2+0.304x+3.7870.92<0.01278652Dy=-0.0023x2+0.2451x + 5.07220.83<0.01总叶面积 Total leaf areaAy=-0.8186x2 + 96.812x-214.650.94<0.013812146Dy = -0.7227x2+75.005x+481.280.460.01生理特征Physiological characteristicsTop-1 叶绿素含量 Ay=-0.0014x2+0.0351x + 37.8450.570.07175359Top-1 Chlorophyll contentDy=0.0023x2-0.5888x+47.3740.810.07Top-3 叶绿素含量Ay=-0.0026x2+0.2513x+34.7130.5610.08165060Top-3 Chlorophyll contentDy =-0.3557x + 44.0160.73<0.01Top-1 叶含水量 Ay=-0.002x+0.83750.85<0.01113464Top-1 Leaf water contentDy=4E-05x2 - 0.0054x+0.86960.90<0.01Top-3 叶含水量 Ay=-0.0022x+0.82630.89<0.01134062Top-3 Leaf water contentDy=-0.0015x+0.8170.87<0.01全部叶片含水量Ay=8E-06x2 - 0.0023x+0.84420.86<0.01154761Water content of all leafDy=2E-05x2-0.004x+0.86660.96<0.01茎含水量 Ay=-0.0015x+0.91340.79<0.01165060Stem water contentDy=1E-05x2-0.0033x+0.93930.93<0.01Top-1 水势 Ay=-0.0081x-0. 870.620.02113464Top-1 water potentialDy = 0.0004x2-0.0547x-0.39270.87<0.01生物量Biomass茎生物量 Stem biomassAy=-0.0058x2+0.8507x - 8.50270.92<0.01216656Dy=-0.0028x2+0.4952x-2.4150.91<0.01根生物量 Root biomassAy = -0.0008x2 + 0.1297x-0.71280.91<0.01226956Dy=-0.0006x2 + 0.0814x+0.250.750.02地上生物量Aboveground biomassAy = 1.4464x-19.4410.95<0.01268253Dy=0.576x+2.85850.92<0.01植株总生物量Total biomassAy=1.5095x-19.0840.96<0.01257954Dy=0.6023x+3.99380.91<0.01
A: 充分灌水Adequate water supply; D: 土壤水分持续减少 Consecutive decrease in soil water
2.3 生理特征
土壤水分持续减少处理组(D)的植株顶部第1片完全展开叶(Top- 1)和第3片叶(Top- 3)的叶绿素含量和叶含水量显著低于充分供水组(A) (图3,表1)、全部叶片的含水量和茎含水量也显著低于充分供水组(A) (图3, 表1)。土壤水分持续减少处理组(D)的Top- 1叶水势显著高于充分供水组(A) (图3, 表1)。同样,两个处理组各生理指标与土壤水分减少持续时间拟合的曲线交点是土壤水分持续减少造成不利影响开始点(图3, 表2),即夏玉米生理指标随土壤水分持续减少受到胁迫的时间阈值,对应的水分亏缺累积量和土壤相对湿度是指标开始受到胁迫的水分亏缺阈值和土壤相对湿度阈值。各生理指标开始受到土壤水分持续减少胁迫的时间阈值如下,Top- 1叶绿素含量、叶含水量和叶水势分别为17、11和11d,Top- 3叶绿素含量和叶含水量分别为16和13d,全部叶含水量为15d,茎含水量为16d;开始受到胁迫时的水分亏缺阈值如下,Top- 1叶绿素含量、叶含水量和叶水势分别为53、34、34mm,Top- 3叶绿素含量和叶含水量分别为50mm和40mm;全部叶含水量为47mm;茎含水量为50mm;开始受到胁迫时的土壤相对湿度阈值如下,Top- 1叶绿素含量、叶含水量和叶水势分别为59%、64%和64%,Top- 3叶绿素含量和叶含水量分别为60%和62%;全部叶含水量为61%;茎含水量为60% (表2)。依据时间阈值的先后,水分亏缺阈值和土壤相对湿度阈值的大小,生理特征对土壤水分减少的敏感性顺序为:顶端第1片完全展开叶的含水量和水势>第3片叶的含水量>全部叶片的含水量>茎含水量=第3片叶的叶绿素含量>顶端第1片完全展开叶的叶绿素含量。

图3 土壤水分持续减少对生理特征的影响 (数据为平均值±标准误)Fig.3 The effects of consecutive soil ware decrease on physiological characteristics (mean±SE)
2.4 生物量
土壤水分持续减少处理组(D)的叶生物量、茎生物量、根生物量、地上生物量和总生物量在处理初期大于充分供水组(A),随着土壤水分持续减少逐渐开始低于充分供水组(A) (图4)。虽然整个生长期两个处理组的叶生物量和茎生物量差异未达到显著水平(表1),但是茎生物量在后期有显著差异的时间段出现。同理于形态和生理指标的时间阈值,水分亏缺阈值和土壤相对湿度阈值,茎生物量、根生物量、地上生物量和总生物量开始受到干旱胁迫的时间阈值分别为21、22、26、25d,水分亏缺阈值分别为66、69、82、79mm,土壤相对湿度阈值分别为56%、56%、53%和54% (表2)。依据时间阈值的先后,水分亏缺阈值和土壤相对湿度阈值的大小,生物量积累对土壤水分减少的敏感性顺序为:茎生物量>根生物量>地上生物量。
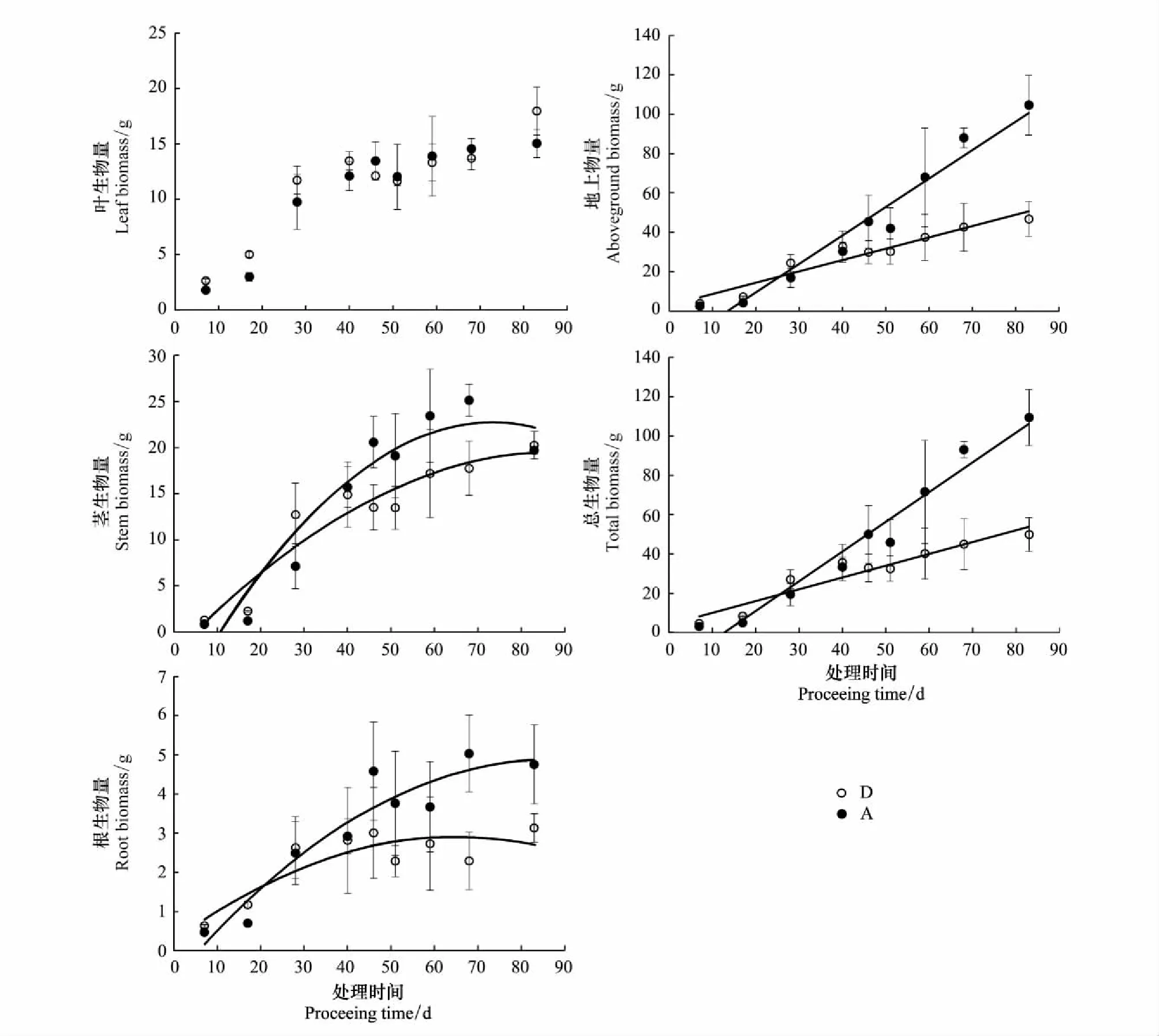
图4 土壤水分持续减少对生物量的影响 (数据为平均值±标准误,n=3)Fig.4 The effects of consecutive soil water decrease on biomass (mean±SE, n=3)
3 讨论
作物对干旱存在阈值响应,即当水分低于一定临界点以后才会对植物生理生态等生长指标造成显著影响,不同生长指标对干旱的敏感性差异导致各自阈值不同,进而出现干旱胁迫症状的时间不同[33-34]。干旱胁迫使玉米发生一系列生理变化,首先叶片等器官的含水量发生响应,叶片光合作用在叶片形态未发生变化时受到影响,随着胁迫程度的加剧,叶绿素合成受到影响,叶片形态颜色变化,进一步限制了光合速率,最终影响到生物量积累和其他形态特征[35]。土壤水分是最常见的作物干旱指标,然而干旱是一个逐步发展的动态过程,静态的土壤含水量只能反映当时的土壤水分状况,包含的干旱累积信息少[3,31,34]。陈家宙等[36]提出了基于累积相对失水量的表达方法。本研究从指标受到干旱胁迫的时间,水分相对亏缺累积量和土壤相对湿度三个方面分析了夏玉米从拔节期开始各生长指标随土壤水分持续减少出现干旱胁迫的阈值。在干旱胁迫持续10d之后,土壤水分相对亏缺累积到34mm,土壤相对湿度下降到64%后各项生理指标陆续开始受到胁迫,20d之后土壤水分相对亏缺累积达到66mm,土壤相对湿度下降到56%后开始影响到生物量的积累,最后影响到株高和叶面积等形态特征。同样体现了干旱发生时先对生理特征产生胁迫后生物量和形态特征的过程。最早开始出现胁迫的是顶端第1片完全展开叶的水势和含水量,导致水势开始减少的土壤相对湿度值为64%与张喜英等[37]所得值(65%)较接近。本研究土壤水分持续减少处理组相对充分供水组在两个半月的时间中累积亏缺244mm,月均约98mm,与玉米生长期所需月平均降水量适宜值100mm左右[2]相近。植物生理生态特征对环境因子变化的响应,是环境因子联合作用的结果。温度、光照、CO2浓度和土壤养分等变化都会影响水分亏缺的效应,在敏感指标阈值的实际运用中应关注其他环境因子与水分交互作用的影响。干旱监测和预警的目的是最大程度消除干旱带来的损害,但同时又要评估减损措施投入与产出比,以获得更大的经济效益。土壤水分持续减少的不同时间和程度下对玉米开展复水等措施产生的效果有较大差异[8,19,22,25,29],准确获取最佳的复水时间和复水程度信息,以达到最佳的效益今后的研究应重点给予关注。
玉米苗期和拔节期进行适当的干旱锻炼,生长发育不会受影响,并且可以提高玉米的抗旱性,最终还可提高生物量积累,苗期田间持水量控制在60%左右,会促进根系发育,进而有助于地上部分物质生产[1-2]。本试验结果显示,土壤水分持续减少处理组(D)的叶生物量、茎生物量、根生物量、株高、叶数、总叶面积和单叶面积在土壤水分持续减少初期与充分供水组(A)相比有增加的趋势(图2,图4),也表明适度干旱胁迫可以促进玉米苗期和拔节期的营养生长。有研究报道苗期干旱是影响玉米第6叶以上叶片生长的一个重要时期[38],本试验土壤水分从拔节期开始减少,苗期未受干旱,所以叶的影响小一些。拔节期是玉米生长中心逐步从根转到茎再到叶的时期,干旱会严重抑制茎粗[16,23-24,38],拔节期开始土壤水分减少,正是玉米生长中心转向茎的关键时期,所以茎的生物量最先因干旱的胁迫而显著降低。
4 结论
本研究以夏玉米郑单958品种为试验材料,设置充分供水(全生育期维持土壤相对湿度为75%±5%)和土壤水分持续减少(拔节前充分供水维持土壤相对湿度为75%±5%,拔节期开始不再补水)两种水分处理,模拟土壤水分持续减少对夏玉米的影响。通过测定株高和叶面积等形态特征,叶水势和叶绿素含量(含不同叶位)等生理特征以及生物量,辨析土壤水分持续减少过程中夏玉米生理生态特征的变化及其受到胁迫的时间阈值和水分亏缺阈值。结果表明,夏玉米拔节期开始土壤水分持续减少,10d之后生理指标开始陆续受到胁迫,20d之后影响到生物量积累,30d左右形态特征开始受到胁迫。夏玉米各生理生态指标对土壤水分减少胁迫的敏感性顺序如下,生理特征:顶端第1片完全展开叶的含水量和水势>第3片叶的含水量>全部叶片的含水量>茎含水量=第3片叶的叶绿素含量>顶端第1片完全展开叶的叶绿素含量,时间阈值分别为11、13、15、16、16、17d,水分亏缺阈值分别为34、40、47、50、50、53mm,土壤相对湿度阈值分别为64%、62%、61%、60%、60%和59%;生物量积累:茎生物量>根生物量>地上生物量,时间阈值分别为21、22和26d,水分亏缺阈值分别为66、69、82mm,土壤相对湿度阈值分别为56%、56%和53%;形态特征:叶数>株高>总叶面积,时间阈值分别为27、35、38d,水分亏缺阈值分别为86、111、121mm,土壤相对湿度阈值分别为52%、47%和46%。植物生长过程受到干旱持续胁迫后发生一系列变化,首先反映在生理特征,然后影响生物量积累,进而对形态结构特征产生影响。
致谢:感谢王敏政、周怀林、王帆和中国气象局固城生态与农业气象试验站在试验观测过程中给予的帮助。
参考文献(References):
[1] 魏湜, 曹广才, 高洁, 衣莹. 玉米生态基础. 北京: 中国农业出版社, 2010.
[2] 何奇瑾. 我国玉米种植分布与气候关系研究[D]. 北京: 中国气象科学研究院, 南京信息工程大学, 2012.
[3] 李柏贞. 玉米干旱过程及其动态模拟[D]. 北京: 中国气象科学研究院, 2014.
[4] 王接弟. 干旱对玉米光合生理及相关酶基因表达的影响[D]. 石河子: 石河子大学, 2014.
[5] Xu Z Z, Zhou G S, Wang Y L, Han G X, Li Y J. Changes in chlorophyll fluorescence in maize plants with imposed rapid dehydration at different leaf ages. Journal of Plant Growth Regulation, 2008, 27(1): 83- 92.
[6] 张仁和, 薛吉全, 浦军, 赵兵, 张兴华, 郑友军, 卜令铎. 干旱胁迫对玉米苗期植株生长和光合特性的影响. 作物学报, 2011, 37(3): 521- 528.
[7] Lobell D B, Roberts M J, Schlenker W, Braun N, Little B B, Rejesus R M, Hammer G L. Greater sensitivity to drought accompanies maize yield increase in the U.S. Midwest. Science, 2014, 344(6183): 516- 519.
[8] Avramova V, AbdElgawad H, Zhang Z F, Fotschki B, Casadevall R, Vergauwen L, Knapen D, Taleisnik E, Guisez Y, Asard H, Beemster G T S. Drought induces distinct growth response, protection, and recovery mechanisms in the maize leaf growth zone. Plant Physiology, 2015, 169(2): 1382- 1396.
[9] 矫梅燕, 周广胜, 张祖强. 农业应对气候变化蓝皮书: 中国农业气象灾害及其灾损评估报告(No.2). 北京: 社会科学文献出版社, 2017.
[10] Piao S L, Ciais P, Huang Y, Shen Z H, Peng S S, Li J S, Zhou L P, Liu H Y, Ma Y C, Ding Y H, Friedlingstein P, Liu C Z, Tan K, Yu Y Q, Zhang T Y, Fang J Y. The impacts of climate change on water resources and agriculture in China. Nature, 2010, 467(7311): 43- 51.
[11] IPCC. Summary for policymakers // Stocker T F, Qin D, Plattner G K, Tignor M, Allen S K, Boschung J, Nauels A, Xia Y, Bex V, Midgley P M, eds. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, United Kingdom: Cambridge University Press, 2013.
[12] Battisti D S, Naylor R L. Historical warnings of future food insecurity with unprecedented seasonal heat. Science, 2009, 323(5911): 240- 244.
[13] Rurinda J, Mapfumo P, van Wijk M T, Mtambanengwe F, Rufino M C, Chikowo R, Giller K E. Comparative assessment of maize, finger millet and sorghum for household food security in the face of increasing climatic risk. European Journal of Agronomy, 2014, 55: 29- 41.
[14] Wang L Y, Yuan X, Xie Z H, Wu P L, Li Y H. Increasing flash droughts over China during the recent global warming hiatus. Scientific Reports, 2016, 6: 30571, doi: 10.1038/srep30571.
[15] Myers S S, Smith M R, Guth S, Golden C D, Vaitla B, Mueller N D, Dangour A D, Huybers P. Climate change and global food systems: potential impacts on food security and undernutrition. Annual Review of Public Health, 2017, 38: 259- 277.
[16] 刘京宝, 杨克军, 石书兵, 赵霞. 中国北方玉米栽培. 北京: 中国农业科学技术出版社, 2012.
[17] 石耀辉, 周广胜, 蒋延玲, 王慧, 许振柱, 麻雪艳. 贝加尔针茅响应降水变化敏感指标及关键阈值. 生态学报, 2017, 37(8): 2620- 2630.
[18] Çakir R. Effect of water stress at different development stages on vegetative and reproductive growth of corn. Field Crops Research, 2004, 89(1): 1- 16.
[19] 纪瑞鹏, 车宇胜, 朱永宁, 梁涛, 冯锐, 于文颖, 张玉书. 干旱对东北春玉米生长发育和产量的影响. 应用生态学报, 2012, 23(11): 3021- 3026.
[20] 李永华, 卢琦, 吴波, 朱雅娟, 刘殿君, 张金鑫, 靳占虎. 干旱区叶片形态特征与植物响应和适应的关系. 植物生态学报, 2012, 36(1): 88- 98.
[21] 姜鹏, 李曼华, 薛晓萍, 李鸿怡. 不同时期干旱对玉米生长发育及产量的影响. 中国农学通报, 2013, 29(36): 232- 235.
[22] 赵文赛, 孙永林, 刘西平. 干旱-复水-再干旱处理对玉米光合能力和生长的影响. 植物生态学报, 2016, 40(6): 594- 603.
[23] 郑盛华, 严昌荣. 水分胁迫对玉米苗期生理和形态特性的影响. 生态学报, 2006, 26(4): 1138- 1143.
[24] 谭国波, 赵立群, 张丽华, 赵洪祥, 方向前, 孟祥盟, 闫伟平, 徐长洪, 韩喜国, 边少锋. 玉米拔节期水分胁迫对植株性状、光合生理及产量的影响. 玉米科学, 2010, 18(1): 96- 98.
[25] 张淑杰, 张玉书, 纪瑞鹏, 刘庆婺. 水分胁迫对玉米生长发育及产量形成的影响研究. 中国农学通报, 2011, 27(12): 68- 72.
[26] 姚春霞, 张岁岐, 燕晓娟. 干旱及复水对玉米叶片光合特性的影响. 水土保持研究, 2012, 19(3): 278- 283.
[27] Siddique M R B, Hamid A, Islam M S. Drought stress effects on water relations of wheat. Botanical Bulletin of Academia Sinica, 2000, 41(1): 35- 39.
[28] 丁雷, 李英瑞, 李勇, 沈其荣, 郭世伟. 梯度干旱胁迫对水稻叶片光合和水分状况的影响. 中国水稻科学, 2014, 28(1): 65- 70.
[30] Shen X F, Dong Z X, Chen Y. Drought and UV-B radiation effect on photosynthesis and antioxidant parameters in soybean and maize. Acta Physiologiae Plantarum, 2015, 37(2): 25.
[31] 张玉书, 米娜, 陈鹏狮, 纪瑞鹏. 土壤水分胁迫对玉米生长发育的影响研究进展. 中国农学通报, 2012, 28(3): 1- 7.
[32] 谭凯炎, 郭建平. 固城生态与农业气象试验站定位观测与研究数据集. 北京: 气象出版社, 2015.
[33] 李伶俐. 夏玉米水分胁迫敏感性及水分适宜度研究[D]. 南京: 南京信息工程大学, 2015.
[34] 麻雪艳, 周广胜. 夏玉米苗期主要生长指标的土壤水分临界点确定方法. 生态学杂志, 2017, 36(6): 1761- 1768.
[35] 孙红春. 不同棉花品种对水分胁迫的形态、生理生化反应[D]. 保定: 河北农业大学, 2015.
[36] 陈家宙, 王石, 张丽丽, 吕国安. 玉米对持续干旱的反应及红壤干旱阈值. 中国农业科学, 2007, 40(3): 532- 539.
[37] 张喜英, 裴冬, 由懋正. 几种作物的生理指标对土壤水分变动的阈值反应. 植物生态学报, 2000, 24(3): 280- 283.
[38] 杨国虎, 李建生, 罗湘宁, 王承莲. 干旱条件下玉米叶面积变化及地上干物质积累与分配的研究. 西北农林科技大学学报: 自然科学版, 2005, 33(5): 27- 32.
猜你喜欢
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
新疆农业科学(2020年1期)2020-02-14
今日农业(2019年11期)2019-08-13
现代园艺(2017年19期)2018-01-19
高原山地气象研究(2016年2期)2016-11-10
中国麻业科学(2015年5期)2015-12-28
塔里木大学学报(2014年3期)2014-03-11
自然资源遥感(2014年2期)2014-02-27
自然资源遥感(2014年2期)2014-02-27
天津农林科技(2011年3期)2011-05-14
