
苏南丘陵区典型林分林内外气温变化规律研究
2018-06-05 08:30庄家尧南京林业大学水土保持与生态修复实验室江苏南京210037
西南农业学报 2018年5期
葛 波,庄家尧,常 猛,张 宇(南京林业大学水土保持与生态修复实验室,江苏 南京 210037)
【研究意义】小气候[1]是指在小范围内由于不同的下垫面构成特征,形成的近地面层和大气候产生差异的现象,温度是小气候观测的重要指标,温度的适宜性关乎着生物生长的状况,直接影响到林分的生长速度、枯枝落叶分解速率、土壤微生物活动[2]。目前国内外已经做过许多关于林内外温度变化规律的研究,徐春蕾[3]基于喀斯特峰丛洼地次生林、坡耕地 2 个动态监测样地的连续观测,探讨了林分和小气候的藕合关系,得出林内外气温的相关性较其他气象因子强,太阳辐射强度的不同和林内外温度变化关系密切,太阳辐射的强度又受到林冠结构、太阳高度角、叶面积指数、林分生长状况的影响[4]。刘效东[5]通过观测南亚热带地区初期、过渡期、顶级 3 种不同演替时期的林分,得出随着森林演替的进行,森林的降温效应会更加显著,其中干季森林对林内外气温调节作用更为明显,对干季森林内部水热供需平衡调节意义重大。郝帅[6]在天山云杉林不同海坡处设立观测站,分析的到5-9月份,林内平均温度、日较差均小于林外且林内外温度日变化规律呈现同样的趋势,随着观测海拔的提高,林内温度每百米的递减率高于林外。杜颖对长白山落叶红松林严寒季节的观测数据分析,探讨了林内气温变化的昼夜垂直梯度,夜间红松林有降温作用,白天红松林有升温作用。任引[7]采用DYYZII型地面气象综合有线遥测系统采集中亚带甜储林的气象要素,分析得到由于甜储林林份林冠的盖幕作用,林内年平均温度小于林外,不同的海拔高度甜储林林内外年平均温度均不同,海拔高度是影响林内外温度变化特征的首要因素。【前人研究进展】关于林内外温度变化的研究气候包括了季风气候、大陆性山地气候、海洋性气候地带,但是关于苏南地区林内外温度变化的单独研究较少,主要研究集中在林内外温度对小气候湿度[8]、土层温度的影响[9],但对林分类型对林内外气温最值出现时刻的滞后性研究较少,掌握气温最值出现时刻的滞后作用是减少热岛效应、增加绿岛效应的有效途径[10]。【本研究切入点】本文通过对苏南地区南京市铜山土壤温度资料,以日、月、季、年为尺度,以15 min为记录单位,对苏南地区3种典型林分和裸地24 h的数据进行统计分析,旨在揭示森林对林内外温度改善的作用,【拟解决的关键问题】为苏南地区小气候变化变化研究提供依据,为合理管理森林资源、保护森林生态环境、最大化森林的效益提供理论基础。
1 材料与方法
1.1 试验区概况
试验区位于苏南地区南京东善桥林场铜山分场(31°35′~31°39′N,118°50′~118°52′E),属亚热带季风气候,地形为苏南丘陵。区内气候温暖湿润,四季分明,水热资源比较丰富,生长季长,降水丰沛,年平均气温15.1 ℃,无霜期 229 d 年日照时间199 h,年平均降水量1100 mm,资源比较丰富,年平均降水量1100 mm,土壤类型为黄棕壤,地形为丘陵,海拔在38~388 m,林分类型以杉木(Cunninghamialanceolata),麻栎(Quercusacutissima),毛竹(Phyllostachyedulis)林为主,均为苏南丘陵区典型林分,林下灌木草本主要以山胡椒[Linderagluca(sieb.et Zucc.)Bl.]、盐肤木(Rhuschinensis)、一年蓬(Erigeronannuus)、荩草[Arthraxonhispidus(Thunb.)Makino]、草蘽(RubustephrodesHance)为主。采用样地法对林分进行取样调查,样地大小为20 m×20 m,坡度均为NE。
1.2 研究方法
选择光照条件较好的试验点,试验点设置考虑上下坡气温的差异,在麻栎林、毛竹林、杉木林、裸地上下坡均设立1个小型气温监测点,上坡距离中心点5 m,下坡距离中心点5 m,取上下坡气温平均值作为瞬时数据。
使用EM50数据收集器记录温度,距离地面1.2 m,定期维护仪器,以保证数据的连续性。观测时间为2011年1月至2012年1月,每15 min 记录1次数据,每1个月收集1次数据。四季的划分采用气象划分法,以阳历3-5月为春季,6-8月为夏季,9-11月为秋季,12月至翌年2月为冬季。取1月11日、4月17日、7月9日、10月7日作为典型日变化,用来说明春夏秋冬四季的温度日变化,其中4 d均为晴天,且前后3 d无阴雨天气;取2011年1月至2012年1月各月瞬时数据的平均值作为该月份的平均温度,用来分析林内外温度的季节变化,取2011年1月至2012年1月各林分的瞬时数据计算林内外温度年均值,分析林分对林内外温度年际变化的影响;取2011年1月至2012年1月的瞬时数据对每日出现的最高温、最低温的频率进行统计,分析各林分的滞后作用。
采用EXCEL软件和SPSS 19统计软件对数据进行分析,使用Origin8绘图。

表1 林分基本特征Table 1 Basic character of forest stand
2 结果与分析
2.1 典型林分林内气温日变化特征
图1选取了一年四季中代表月份(1、4、7、10月)典型日林内外气温日变化特征。各个季节的林内和林外的气温日变化有相似的基本规律并且均呈正弦函数变化曲线,春秋夏季0:00-7:00气温逐渐降低,冬季0:00-2:00气温逐渐降低,随着太阳辐射的开始,气温在7:00后逐渐上升,并于14:00达到最高气温,14:00-24:00逐渐下降。春秋夏季的日变化幅度较冬季大,其中春季气温日变化较剧烈,麻栎林毛竹林裸地的日变化幅度较杉木大。整体来说日最高气温裸地最高(4个典型日平均23.2 ℃),其次是麻栎林(20.6 ℃),毛竹林(20.2 ℃)和杉木林(18.7 ℃)日最高气温相对较小。随着林内外太阳辐射不同,呈现夜晚林内>裸地、白天裸地>林内,白天林分由于林冠的反射和吸收作用消弱了到达林内的太阳辐射,夜晚由于林冠对雾、露水的凝结截留作用使林内气温比林外气温高,因此林内气温的日变化幅度较林外小,且森林有白天降温、夜晚保温的作用。
2.2 典型林分林内温度季变化
2.2.1 月气温均值统计 表2是统计了2011-2012年各月份不同林分类型的气温平均值,表3是麻栎林、毛竹林、杉木林各月平均气温和裸地的差值。由表2~3可知,麻栎林、毛竹林、杉木林大部分月平均气温都小于裸地气温,3-10月林内气温均小于林外气温,5-7月份3个树种林内外温差达到外气温的平均值大于杉木林,麻栎林其次。在1、2、11月出现了林内气温大于林外气温的现象,麻栎林升温作用大于杉木,毛竹林其次,这是因为冬季麻栎最大值,说明麻栎林、毛竹林、杉木林在生长的旺季都能有效的降低林内外温度,其中毛竹林降低林内林、毛竹林、杉木林均会落叶,由于冬季林冠郁闭度变小,枯枝落叶量增加,使林下接收太阳辐射,土壤微生物的活动更为活跃,枯落物分解释放出更多的热量,导致地表温度升高,麻栎林林、毛竹林、杉木林落叶量是麻栎林>杉木林>毛竹[11-13]。综合分析12个月的气温,麻栎林、毛竹林、杉木林均为7月份气温最高,分别是26.6、25.6、25.5 ℃,林外裸地月平均气温也是7月份最高,为27.3 ℃。各林分林内各月平均气温比较见图2。
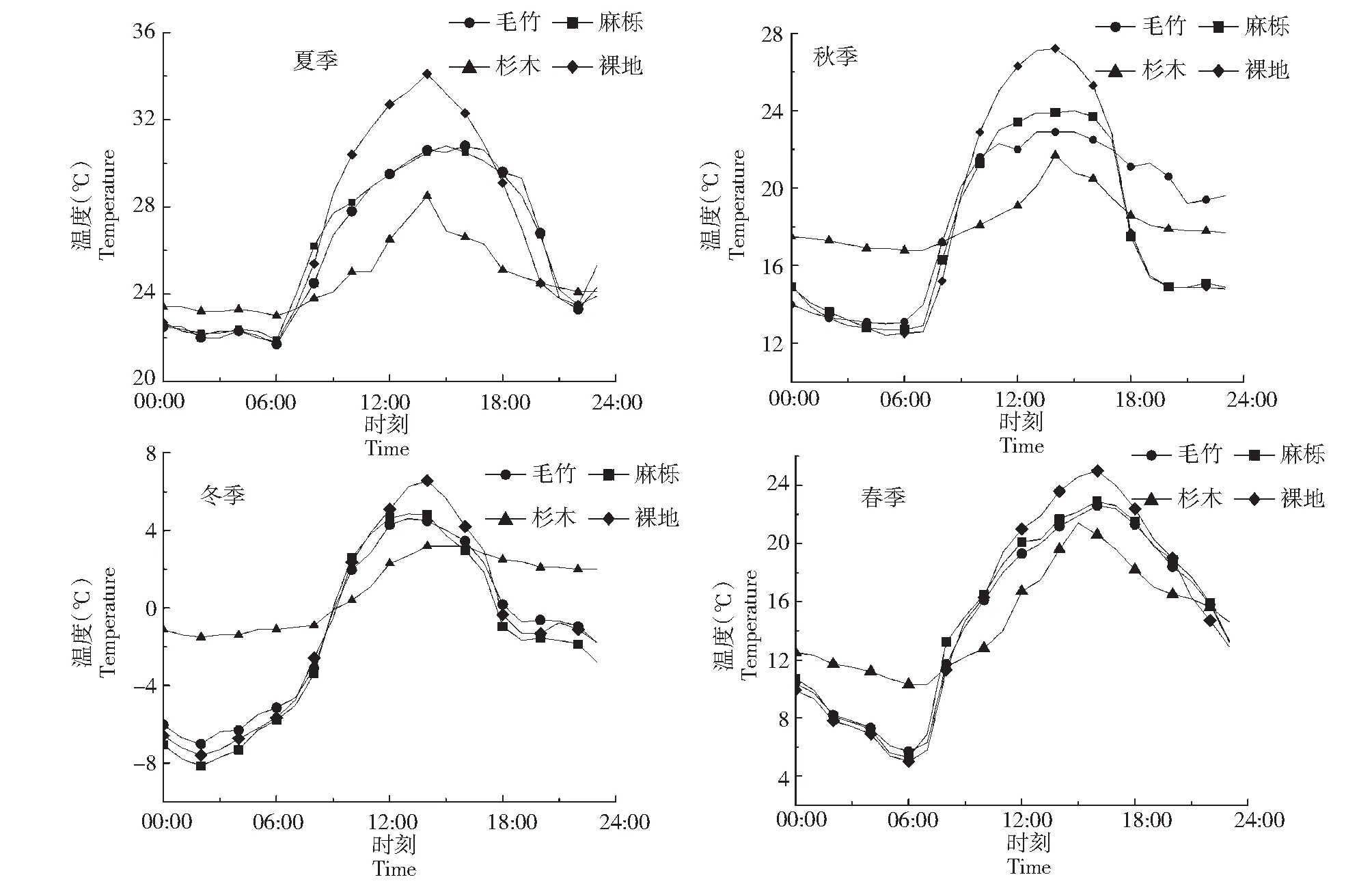
图1 不同季节林内外温度日变化Fig.1 Diurnal variation of temperature inside and outside forest in different seasons
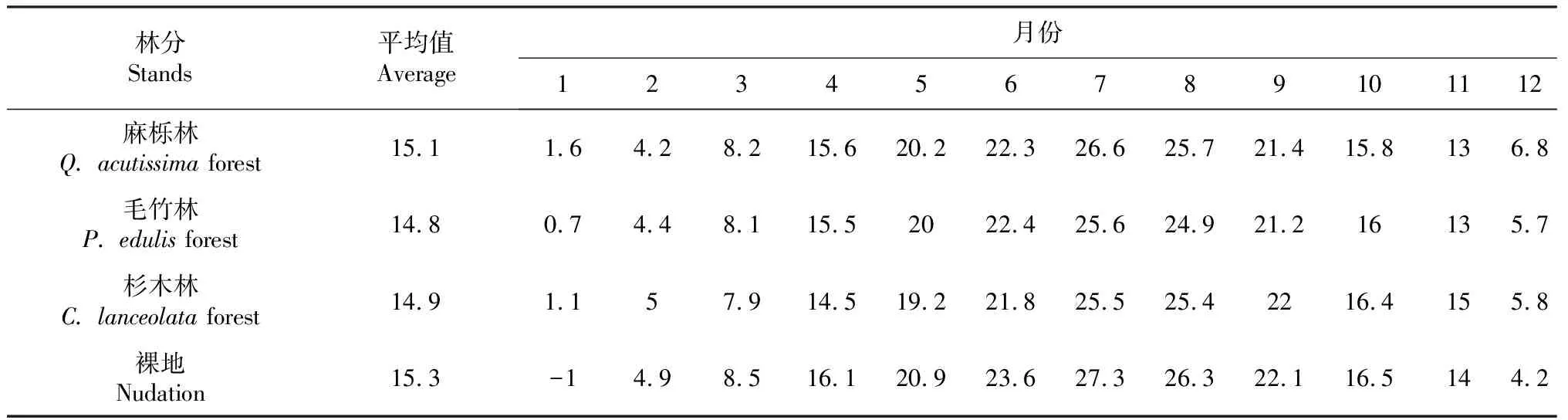
表2 各月不同林分内的平均温度Table 2 Average temperature in different months in each month

表3 各林分与裸地平均温度差值Table 3 Average temperature difference between the stands and bare land

图2 各月各林分内平均温度Fig.2 Monthly average temperature of each forest
2.2.2 季气温均值及标准偏差统计 由图3可以看出,各林分林内温度季均值和2011-2012年各季瞬时数据的标准偏差,在春季麻栎林、毛竹林、杉木林均能降低林内温度,毛竹林的效果最佳(3.3 ℃)杉木林次之(1.2 ℃)麻栎林效果不明显。在夏季各林分均能降低林内温度,其中麻栎林和杉木林效果相当,麻栎林(2.8 ℃)、杉木林(3.5 ℃),毛竹林次之(2 ℃)。在秋季毛竹林能降低林内温度较明显(3.8 ℃),麻栎林和杉木林则效果不明显(0~0.6 ℃),在冬季杉木林能提高林内温度(1.6 ℃)麻栎林和毛竹林均能降低降低林内温度,其中麻栎林降低1 ℃,毛竹林降低1.7 ℃。春夏秋冬四季麻栎林、毛竹林、杉木林的标准偏差均小于裸地,说明林分的存在使气温季节变化更加稳定,波动更小。
2.3 典型林分林内温度的年际变化
2.3.1 年均值及年较差统计 表4是对各林分林内温度年均值和年较差进行统计分析,各个林分的年较差均小于裸地,年较差大小依次为:裸地、麻栎林、毛竹林、杉木林,表明了毛竹林和麻栎林一年内林内温度变化幅度较大,杉木林变化幅度最小。各林分年林内温度均值的大小依次为:裸地、毛竹林、杉木林、麻栎林。表明,各个林分均能降低林内温度,其中,麻栎林林的效果最好,能降低0.5 ℃,毛竹林和杉木林效果一般,能降低年均值0.3、0.4 ℃。

表4 林内外温度年均值Table 4 Average temperature in and outside the forest

图3 各林分林内温度季均值Fig.3 Average temperature of each forest in the forest
2.3.2 滞后分析 通过对各林分和裸地林内温度的最高温及最低温出现的时刻进行频率统计分析(表5,图4~5),裸地林内气温出现最值的时间最早,其次是毛竹林、麻栎林、杉木林,麻栎林约滞后45 min,毛竹林约30 min,杉木林约滞后75 min;麻栎林、毛竹林、杉木林、和裸地出现最低温的时间大致相同,均在23:45,次低温中,裸地最先出现次低温,其次是杉木林、毛竹林、麻栎林,其中杉木林和毛竹林大致相同,约滞后30 min,麻栎林约滞后75 min。苏南地区属于亚热带气候,试验地春秋季节短、冬夏季节长,冬夏气温差显著,且低气温较集中与凌晨零点,因此,试验中最低温出现时间大致相同,凌晨后次低温的出现时间能体现出森林对气温最值出现的时刻的滞后现象。
2.3.3 极值分析 图6~7显示,通过极值分析的方法计算各个林地和裸地各月林内气温最高值最低值的差值,使用裸地温度减去各林分温度,麻栎林、毛竹林、杉木林林内月最高气温基本高于裸地,总的来说杉木林与裸地的林内温度最高温之差最高,麻栎林和毛竹林与裸地林内最高温差异不显著。麻栎林、毛竹林、杉木林和裸地月最低温的差值规律基本相似,杉木林各个月的林内最低温均比裸地高,麻栎林和毛竹林对林内最低温提升作用不显著。表明各林分均有降低林内最高温的作用、提高林内最低温的作用,杉木林作用最为显著,麻栎林次之,毛竹林效果相比最差。

表5 林内温度最高温与最低温出现频率峰值点Table 5 Temperature of the forest temperature peak value of the peak point

图4 林内最大温度出现时刻频率Fig.4 Maximum frequency of temperature
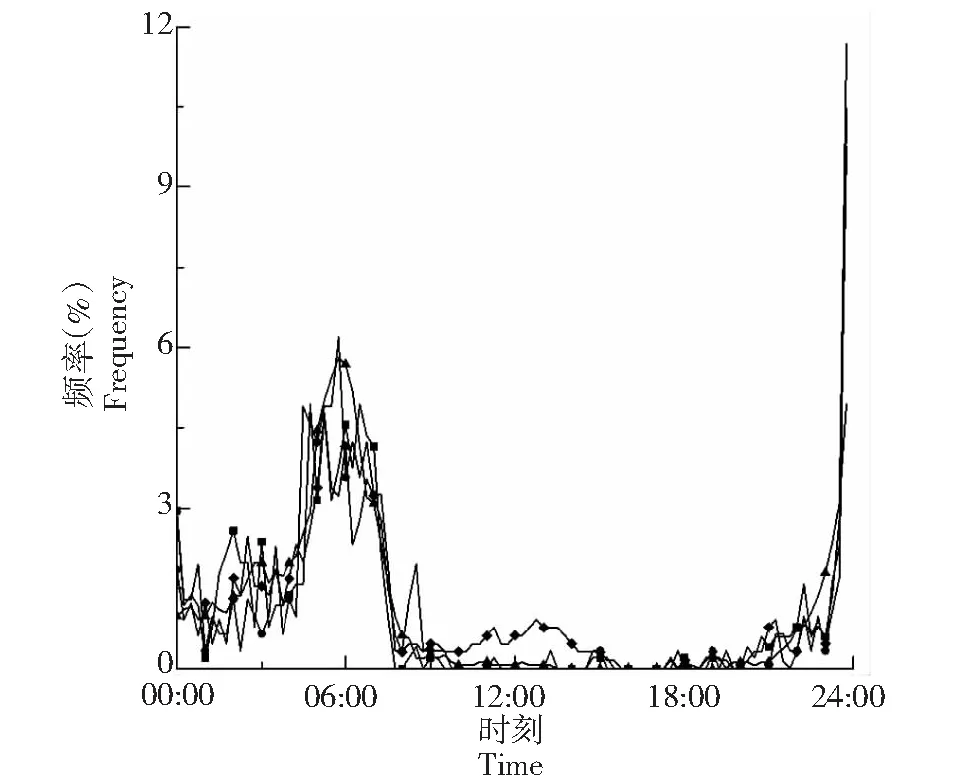
图5 林内最小温度出现时刻频率Fig.5 Minimum temperature frequency of occurrence
3 结 论
(1)在不同的季节中,森林对林内外气温的改变能力不同,其中春季气温日变化较剧烈,随着林内外太阳辐射不同,呈现林内>裸地、裸地>林内、林内>裸地3个阶段;在森林生长旺季的夏季,林内气温日变化较平缓,这与王景升[13]的研究结果相一致,林内由于林冠层的存在和枯枝落叶的覆盖作用大大减少了到达林冠层的太阳辐射,使林内气温的震荡幅度较裸地小。麻栎林、毛竹林的日变化幅度较杉木大,说明麻栎林和毛竹林能使林内气温处于稳定的状态,更加适合林内生物生长。

图6 各林分与裸地月最高温差值Fig.6 Difference monthly maximum temperature in stands and nudation
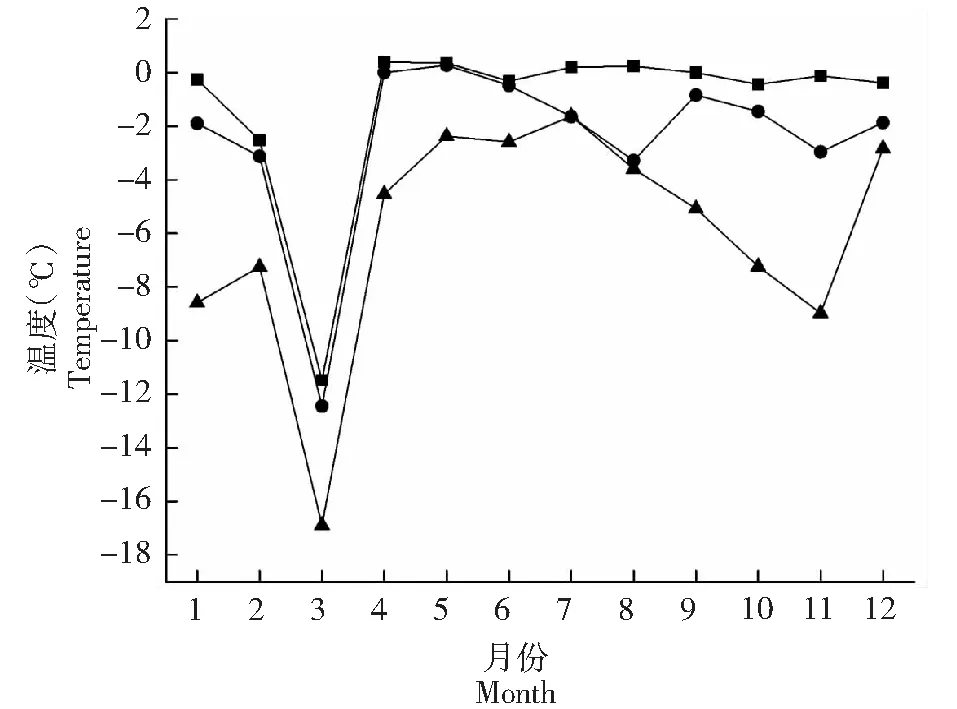
图7 各林分与裸地月最低温差值Fig.7 Difference monthly minimum temperature in stands and nudation
(2)森林在春、夏、秋季均有降低林内气温,冬季除了杉木林能提高林内气温外,麻栎林和毛竹林均能有效的降低林内气温,这是因为冬季杉木林落叶量大,由于林冠下接收太阳辐射的增加和枯落物分解释放热量,导致林内温度的提高。在生长旺季5-7月份森林的林内外温差达到最大值,说明在生长旺季森林能有效的降低林内气温,这与徐云蕾的研究结果相一致。对比3个林分的气温季节变化,可以得出春秋季毛竹林降低林内气温效果最好,夏季麻栎林和杉木林降低林内气温的效果最好,整体来说,毛竹林降低林内气温的效果最好,其次是杉木林,最后是麻栎林。
(3)森林能有效的降低林内气温的年均值,各林分年林内气温均值的大小依次为:裸地、毛竹林、杉木林、麻栎林,分析各个林分的林内气温年较差可知麻栎林降低气温年均值的效果最好,能降低年均值0.5 ℃,麻栎林、毛竹林、杉木林林内气温年较差均小于裸地气温年较差,说明森林内林内气温的变化缓和,林外温度低时森林起到了保温作用,温度高时有降温作用,这与J. Biometeor[14]研究相一致。
(4)森林有滞后林内气温最高温、次低温出现时间的作用,在本文中,最低温出现时刻,麻栎林、毛竹林、杉木林和裸地相一致,主要是因为研究区域的日气温最低时刻一般都是在23:00-24:00,随后才会慢慢升温。整体来说,麻栎林的滞后作用较好,其次是杉木林,毛竹林的效果最差。表明了森林林冠层直接影响到林内最高温、最低温的出现时间。森林对气温的滞后作用可用于缓解城市内部的热岛效应,合理的选择树种达到森林生态效益的最大化。
(5)森林有降低林内气温最高值温,提高林内气温最低值的作用。林内外月气温最高值、最低值的差值与林冠的覆盖率呈正相关关系,杉木林作用最为显著,麻栎林次之,毛竹林效果最差,这与欧阳学军[15]的研究相一致。林冠的存在削弱了太阳辐射到达林内的强度,减少了林内外气温的冷热交换,正是林冠对气温调节的正作用,使森林具有了降低高温,提高低温的作用。
4 讨 论
综合所述,本文所研究的苏南丘陵地区3种典型树种,麻栎林和毛竹林能使林内气温处于相对稳定的状态,春秋季毛竹林降低林内气温的能力优于麻栎林和杉木林,夏季麻栎林和杉木林降低林内气温的能力更好,麻栎林降低气温年均值的效果优于杉木林和毛竹林,麻栎林的滞后林内气温最值出现时间的作用较好,其次是杉木林,毛竹林的效果最差。杉木林降低林内最高温,提高林内最低温的作用较明显。此结论可作为苏南丘陵区森林资源合理分配开发的指导。
参考文献:
[1]唐金生,张如良,张青云. 林分小气候特征的研究[J]. 华东森林经理,2006(2):27-29.
[2]冯增林,毛利慧. 农田小气候改善途径[J]. 现代农村科技,2012(11):78-78.
[3]徐云蕾,曾馥平,宋同清. 喀斯特峰丛洼地次生林小气候特征研究[J]. 农业现代化研究,2012,33(2):239-244.
[4]刘效东,周国逸,陈修治. 南亚热带森林演替过程中小气候的改变及对气候变化的响应[J]. 生态学报,2014,34(10):2755-2764.
[5]郝 帅. 天山中段天山云杉林森林小气候特征研究[D]. 新疆农业大学,2007.
[6]杜 颖,关德新,殷 红. 长白山阔叶红松林的温度效应[J]. 生态学杂志,2007(6):787-792.
[7]任 引,薛建辉. 中亚热带甜槠常绿阔叶林小气候特征分析[J]. 南京林业大学学报(自然科学版),2008(3):14-18.
[8]韩 诚,张金池,庄义琳,等. 苏南丘陵区麻栎林冠层水文效应及其影响因素[J]. 南京林业大学学报(自然科学版),2014,38(2):93-98.
[9]杨 丹,杨 静. 南京城郊毛竹林内外气温及土壤温度特征[J]. 东北林业大学学报,2017,45(3):68-73.
[10]周红妹,丁金才,徐一鸣. 城市热岛效应与绿地分布的关系监测和评估[J]. 上海农业学报,2002(2):83-88.
[11]方德齐. 麻栎象生活习性的初步观察[J]. 昆虫知识,1981(5):207-208.
[12]迟 健. 马尾松和杉木生态习性的比较[J]. 湖南林业科技,1992(2):31-35.
[13]王景升,潘 刚. 急尖长苞冷杉原始林内外温度特征研究[J]. 西藏科技,2004(7):48-52.
[14]Fetcher N Oberbauer S F, Strain B R Vegetation effects on microclimate in lowland tropical forest in Costa Rica[J]. Int. J. Biometeor, 1985, 29(2): 145- 155.
[15]欧阳学军,黄忠良,周国逸,等. 鼎湖山4种主要森林的温度和湿度差异[J]. 热带亚热带植物学报,2003(1):53-58.
猜你喜欢
天津农林科技(2022年2期)2022-04-19
浙江农林大学学报(2020年5期)2020-11-05
防护林科技(2020年6期)2020-08-12
环保科技(2020年2期)2020-05-21
湖州职业技术学院学报(2020年4期)2020-03-30
浙江农林大学学报(2019年6期)2019-11-13
装备环境工程(2019年9期)2019-10-24
绿色科技(2019年6期)2019-04-12
绿色科技(2019年6期)2019-04-12
小小说月刊·下半月(2015年11期)2015-05-14
