
成都地区番茄主要害虫和天敌种类及优势类群发生规律研究
2018-06-05 08:30蒲德强王小强刘虹伶戢沛城龚一耘叶仕伦周肆维李跃建
西南农业学报 2018年5期
蒲德强,王小强,刘虹伶,戢沛城,龚一耘,叶仕伦,黄 静,周肆维,张 梅,祝 进,李跃建
(1. 四川省农业科学院经济作物育种栽培研究所,四川 成都 610300;2. 四川省农业科学院植物保护研究所,四川 成都 610066;3. 四川省农业厅,四川 成都 610041;4. 四川省农业科学院园艺研究所,四川 成都 610066)
【研究意义】成都因独特的地理位置和生态条件使其成为四川地区主要蔬菜种植区域,而番茄Lycopersionesculentum作为茄科类蔬菜的一种,因可生食、煮食、加工且含有丰富的营养在成都地区被广泛种植。【前人研究进展】研究发现成都地区蔬菜重要害虫包括昆虫纲7目33科86种[1],但对番茄田相关研究未见报道。【本研究切入点】在实际生产中,菜田生态系统因受到蔬菜品种、生长周期、换茬及反季节栽培等因素影响造成其昆虫群落结构稳定性差,昆虫种类及数量变化幅度大,而成都地区适宜的温湿度为蔬菜病虫害发生提供了有利条件,从而给蔬菜害虫防治及其天敌利用带来很大困难[2]。【拟解决的关键问题】本研究旨在通过2015-2016年2年间对成都地区番茄田主要昆虫群落调查,及主要害虫和天敌消长动态研究,从而为准确及时地对番茄虫害采取防治措施及提高番茄产量和质量提供理论依据。
1 材料与方法
1.1 试验地概况
本试验于2015-2016年连续2年在四川省农业科学院经济作物育种栽培研究所实验基地(成都市青白江区城厢镇)进行,供试番茄品种为红盾,试验田面积1000 m2(2015年试验田周边主要种植大豆和棉花,2016年试验地周边主要种植烟草和棉花),以当地普遍采用的深沟高箱方式栽培,除在番茄移栽时选用0.5 %阿维菌素颗粒剂1 kg/667m2进行穴施以防治地下害虫外,番茄整个生长期内均不施用任何化学农药,农事操作按常规进行。
1.2 调查方法
调查采样时间为番茄移栽后20 d至采摘结束,2015年5月至9月和2016年6月至9月,每10~12 d调查1次(雨天顺延),每次调查时间为昆虫活动频繁的10:00-16:00,采用固定五点调查法,每点系统调查15株并记录每株出现昆虫的数量、种类,将田间无法鉴定的昆虫种类保存于75 %乙醇溶液带回实验室鉴定。因蜘蛛类亦为大部分昆虫主要天敌,螨类为农作物主要害虫,故将其统计在内。
1.3 标本鉴定
昆虫标本在体视显微镜(型号:Zeiss Discovery V8, Germany)下根据中国动物志、中国经济昆虫志等鉴定,部分不能鉴定的种请国内相应类群的昆虫分类学家鉴定。
1.4 数据处理
部分气象数据来自成都市植保站,所有数据均通过Excel 2007及Spss 19.0统计分析
2 结果与分析
2.1 成都地区番茄害虫和天敌种类
鉴定结果表明,成都地区番茄田节肢动物主要包括昆虫纲8目19科37种及蛛形纲2目4科5种共42种,其中害虫主要集中在半翅目、鳞翅目及直翅目等昆虫纲6目13科23种及蛛形纲1目1科1种共24种共占比58 %(表1),天敌主要集中在脉翅目、双翅目及蜘蛛目等昆虫纲5目7科14种及蛛形纲1目3科4种共18种共占比42 %(表2)。

表1 成都地区番茄害虫种类Table 1 The species of pests on tomato in Chengdu
2.2 成都地区番茄主要害虫类群发生规律
如图1~2所示,2015 -2016年成都地区番茄主要害虫为粉虱、蚜虫及蛾类,其中以粉虱为重,相比蚜虫及蛾类,粉虱在2年间均出现了较大幅度动态变化,在2015年7月22日及2016年8月5日分别达到高峰;2年间5-8月的温度均在20~34 ℃变化,这一温度为害虫的发生提供有利条件。同时,2年间5-8月的平均温度及降雨天数均无较大差异,但是大于2级风力天数2016年较2015年多8 d,尤其表现在2016年7-8月 (表3)。
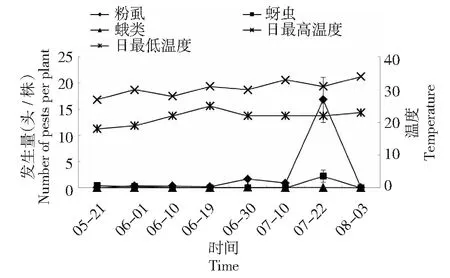
蛾类指烟青虫和斜纹夜蛾,其他的食叶蛾类较少,未计入,下同Moths including only Heliothis assulta and Prodenia litura.The same as below图1 2015年成都地区番茄主要害虫发生动态及温度变化规律Fig.1 The occurrence dynamics of dominant pests on tomato and law of temperature change in Chengdu in 2015
2.3 优势天敌发生规律
本试验对蛛形纲蜘蛛目天敌调查表明,2015-2016年连续2年捕食性蜘蛛始见期均与番茄虫害始见期保持一致,并随着害虫数量变化而出现动态波动,在2015年7月19日及2016年7月25日分别达到最大值1.5、0.8头/株。
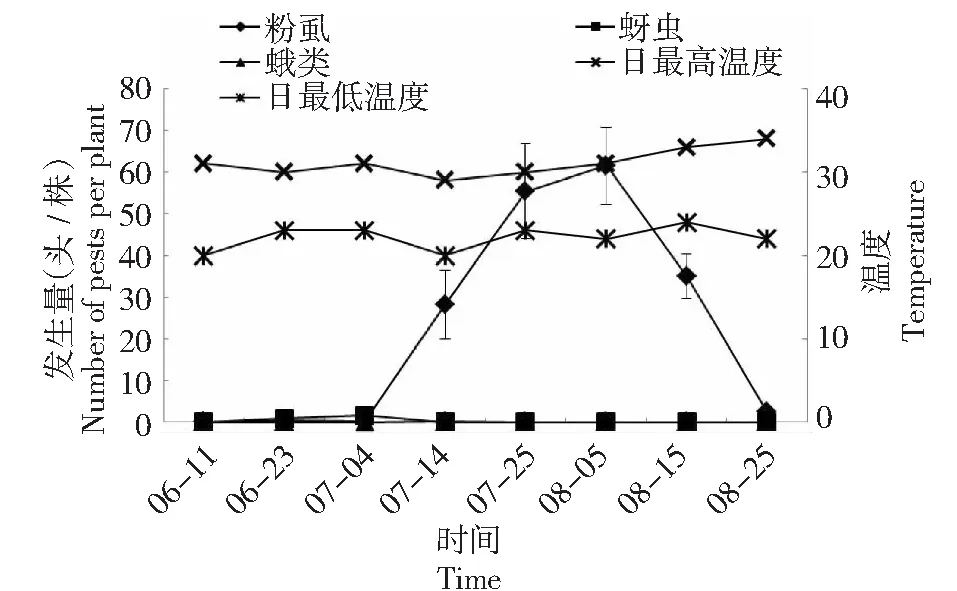
图2 2016年成都地区番茄主要害虫发生动态及温度变化规律Fig.2 The occurrence dynamics of dominant pests on tomato and law of temperature change in Chengdu in 2016
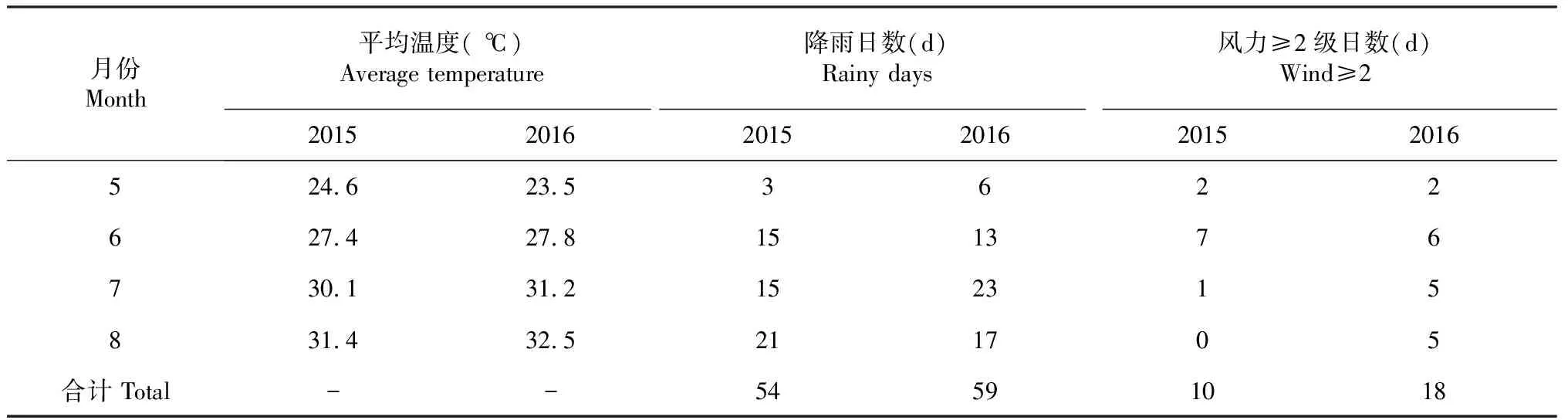
表3 成都地区2015-2016年5-8月主要气象参数Table 3 The meteorological parameter of 5-8 months in 2015 and 2016 in Chengdu
2.4 捕食性蜘蛛与主要害虫相关性分析
以捕食性蜘蛛为因变量,主要害虫为自变量建立线性回归方程表明,2015年害虫始发期,捕食性蜘蛛与其表现为极显著的线性关系(P<0.01),但随着害虫数量的不断加剧,捕食性蜘蛛与主要害虫的线性关系逐渐减弱甚至表现为线性关系消失;2016年害虫始发期与末期,捕食性蜘蛛与主要害虫均表现出极显著(P<0.01)或显著(P<0.05)的线性关系,而在害虫盛发期,捕食性蜘蛛与主要害虫的线性关系不显著(P>0.05,表4)。
3 讨 论
昆虫作为动物界中最大类群,与人类有着密切关系,尤以农业害虫类表现为甚,因而引起国内外大多数昆虫学家对其展开种群生态学,个体生物学及分子生物学等相关方面的研究[3-5]。黄保宏等[6]研究表明,昆虫群落是了解群落结构特征,更好地发挥自然因素的生态调控作用的重要突破口,不少研究人员对花椒园、梅园、桃园、大豆田、棉田、麦田及油菜田等[7-13]昆虫群落结构及动态进行了研究,而且昆虫群落生态学是害虫综合治理 (Integrated Pest Management,IPM) 策略和实践的理论基础和前提,其中群落结构已成为评价害虫综合治理效果的指标之一[14-15]。因此,探讨番茄田中昆虫群落在时空上的运动变化规律及机制,是开展番茄田有害生物综合生态调控的理论基础,烟粉虱作为小型害虫,在其虫源周围一定距离迁移受风力影响明显[16],本研究发现,2015-2016年2年间成都地区番茄粉虱的发生量差异明显,这可能与捕食性蜘蛛2年间发生量不同有一定关系,另外,2年间5-8月的温度均在20~34 ℃变化,这一温度为害虫的发生提供有利条件,2年间5-8月的平均温度及降雨天数均无较大差异,但是大于2级风力天数2016年较2015年多8 d,尤其表现在害虫盛发期的7-8月,从而为粉虱在一定范围内的转移提供了有利条件;而且2015年试验田周边主要以大豆和棉花栽培为主,2016年试验田周边主要以棉花、烟草育种栽培为主,从而为粉虱提供了更适宜的寄主,以上各因素使得2016年试验田粉虱发生量明显高于2015年,但关于成都地区气象因子与粉虱类害虫发生动态的关联性还需进一步深入研究。
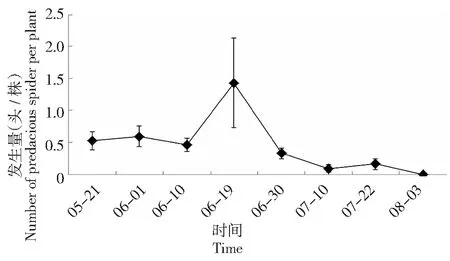
图3 2015年成都地区番茄害虫主要天敌——捕食性蜘蛛发生动态Fig.3 The occurrence dynamics of predacious spiders on tomato in Chengdu in 2015
蜘蛛作为蛛形纲蜘蛛目动物,因其可捕食大量农林害虫而成为我国开展以虫治虫研究的对象之一,姜瑞德等[17]研究发现捕食性蜘蛛是茶园害虫主要天敌之一,亦是飞虱的重要天敌,仅取食假眼小绿叶蝉Empoascavitis的蜘蛛类就有20科80种[18]。
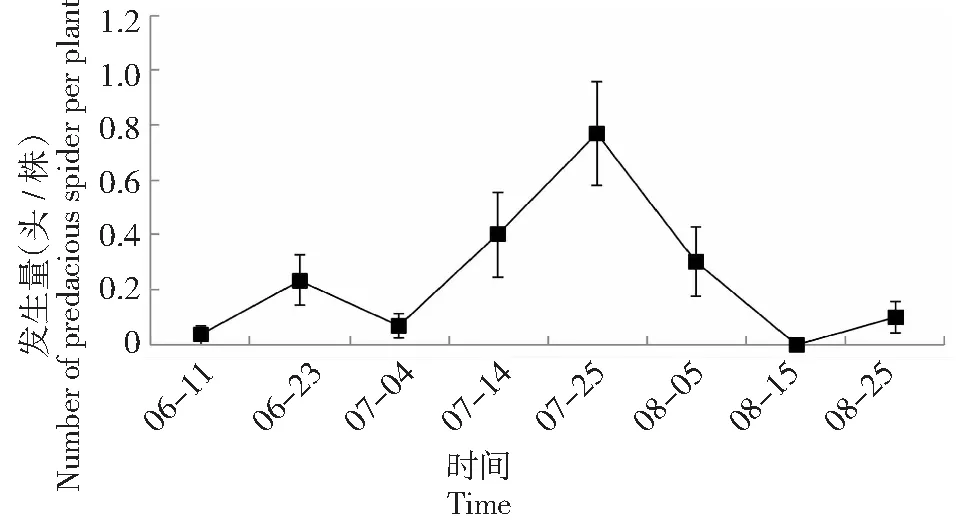
图4 2016年成都地区番茄害虫主要天敌——捕食性蜘蛛发生动态Fig.4 The occurrence dynamics of predacious spiders on tomato in Chengdu in 2016

表4 捕食性蜘蛛与主要害虫的线性关系Table 4 The linear relationship between predacious spiders and dominant pests
吴进才等[19]研究表明田间蜘蛛密度增大时其对飞虱的取食量反而降低,这可能与种间竞争及飞虱在适宜环境下短期内爆发有关,赵静等[20]通过利用BlockKriging进行空间插值分析表明蜘蛛种群在较大程度上与烟盲蝽种群存在跟随关系,王成树等[1]研究发现蔬菜害虫和天敌在不受人为防治的干扰下,天敌群落和害虫群落在结构上有动态的消长规律。本研究发现,番茄田捕食性蜘蛛发生时序动态亦追随害虫发生动态且均表现出动态的消长规律,2年间在害虫始发期均表现为捕食性蜘蛛与害虫显著的线性关系,随着害虫数量的不断增加,捕食性蜘蛛对其的控制能力有所减弱,但捕食性蜘蛛发生规律并不与害虫发生规律完全同步,这可能与捕食性蜘蛛食谱范围广,活动能力强等因素有关。
我国是世界上最早开展人工生物防治的国家,但直到1988年美国利用澳洲瓢虫Rodoliacardinalis成功控制吹绵蚧Iceryapurchasi后,生物防治才被广大植保工作者接受并重视[21],中科院院士蒲蛰龙教授提出的 “以发挥害虫天敌效能为主的害虫综合防治”成为我国开展以虫治虫研究新的开端[22],大批科研工作者对瓢虫,食蚜蝇,草蛉及寄生蜂等天敌对猎物选择,胚胎发育,人工繁殖等方面进行研究[18, 23]。本研究明确了成都地区番茄昆虫群落结构及主要类群发生规律,下一步将针对粉虱和蛾类害虫系统开展生物防治研究。另外,在番茄采摘后期,朱砂叶螨发生量较大,危害严重,值得关注。
4 结 论
成都地区番茄田主要包括昆虫纲8目19科37种及蛛形纲2目4科5种共42种,其中害虫占比58 %,天敌占比42 %。2015-2016年2年间成都地区番茄粉虱的发生量差异明显,这与捕食性蜘蛛的发生量及气象因子存在关系。
致谢:感谢浙江大学昆虫科学研究所吴琼老师鉴定部分标本,中国科学院动物研究所张魁艳老师帮忙鉴定部分食蚜蝇标本,西北大学谭江丽博士帮忙鉴定部分胡蜂标本。
参考文献:
[1]谢大实. 成都蔬菜害虫初步名录[J]. 四川大学学报(自然科学版), 1955(7):70-74.
[2]王成树,陈树仁. 蔬菜害虫及其天敌昆虫群落多样性和相关性研究[J]. 生物多样性, 1999, 7(2):106-111.
[3]Alon S, Leon B, Yoel M. Influence of salinity concentration on aquatic insect community structure: a mesocosm experiment in the dead sea basin regin[J]. Hydrobiologia, 2005, 584:1-10.
[4]Makoto K, Tamiji I, Abang AH,et al. Seasonality and vertical structure of light-attracted insect communities in a dipterocarp forest in sarawak[J]. Population Ecology, 1995, 37(1):59-79.
[5]马世骏. 昆虫种群的空间、数量、时间结构及其动态[J]. 昆虫学报, 1964,13(1):39-55.
[6]黄保宏,邹运鼎,毕守东,等. 梅园昆虫群落的时间结构及动态[J]. 应用与环境生物学报, 2005, 11(1):187-191.
[7]张承祚,姚利忠,李 强,等. 云南不同海拔花椒园昆虫群落结构及动态[J]. 应用生态学报, 2006, 16(2):307-312.
[8]黄保宏,邹运鼎,毕守东,等. 梅园昆虫群落特征、动态及优势种生态位[J]. 应用生态学报, 2005, 16(2):307-312.
[9]邹运鼎,毕守东,周夏芝,等. 桃园害虫及天敌群落动态研究[J]. 应用生态学报, 2003, 14(5):717-720.
[10]候中一. 大豆田昆虫群落结构及天敌对蚜虫的调控功能[D]. 哈尔滨: 东北农业大学, 2007.
[11]吕文彦,秦雪峰,徐艳聆,等. 不同类型棉田昆虫群落结构研究[J]. 中国生态农业学报, 2009, 17(2):330-334.
[12]高书晶,庞 保,周 晓,等. 麦田昆虫群落结构及多样性的季节动态[J]. 应用昆虫学报, 2006, 43(3):295-299.
[13]卢 申,江文娟,李桂亭,等. 油菜田节肢动物群落结构及其模糊聚类分析[J]. 中国农学通报, 2008, 24(11):365-370.
[14]句荣辉,沈佐锐. 昆虫种群动态模拟模型[J]. 生态学报, 2005, 25(10):2709-2716.
[15]刘兴平,李 冬,张林平,等. 退化红壤区不同退耕还林模式昆虫群落结构及其多样性特征分析[J]. 环境昆虫学报, 2011, 33(4):471-477.
[16]张网定,周福才,田 晨,等. 基于气象因子的烟粉虱发生预测[J]. 内蒙古民族大学学报(自然科学版), 2015, 30(1):39-42.
[17]姜瑞德,王继青,张 涛,等. 7种农药对茶园假眼小绿叶蝉和捕食性蜘蛛的影响研究[J]. 安全与环境学报, 2011, 11(2):12-14.
[18]戴 轩. 假眼小绿叶蝉的天敌蜘蛛种类及其物种多样性研究. 环境昆虫学报, 1999, 21(4):165-169.
[19]吴进才,姜永厚,邱慧敏,等. 田间蜘蛛集团对飞虱的每日捕食量与室内、盆栽水稻的捕食作用比较[J]. 生态学报, 2002, 22(8):1260-1269.
[20]赵 静,赵 鑫,王玉军,等. 烟盲蝽及其天敌蜘蛛空间格局的地统计学分析[J]. 生态学报, 2010, 30(15):4196-4205.
[21]陈学新. 21世纪我国害虫生物防治研究的发展、问题与展望[J]. 应用昆虫学报, 2010, 47(4):615-625.
[22]古德祥,张古忍,张润杰,等. 中国南方昆虫生物防治50周年回顾[J]. 昆虫学报, 2000, 43(3):327-335.
[23]庞 虹. 捕食性瓢虫的利用[J]. 环境昆虫学报, 1996, 18(4):30-36.
猜你喜欢
果农之友(2022年4期)2022-05-09
广东蚕业(2021年12期)2022-01-18
落叶果树(2020年2期)2020-04-20
小朋友·快乐手工(2018年3期)2018-04-22
现代园艺(2018年23期)2018-01-18
小学阅读指南·低年级版(2017年6期)2017-06-12
中国新闻周刊(2015年31期)2015-08-28
小朋友·快乐手工(2015年1期)2015-03-13
