
云南茶树地方品种与野生茶遗传多样性的ISSR分析
2018-06-05 08:30孙雪梅赵才美李友勇程在全尚卫琼杨盛美杨兴荣刘本英
西南农业学报 2018年5期
孙雪梅,赵才美,李友勇,程在全,尚卫琼,杨盛美,杨兴荣,矣 兵,刘本英*
(1. 云南省农业科学院茶叶研究所,云南 勐海 666201;2. 云南省茶学重点实验室,云南 勐海 666200;3. 云南省农业科学院生物技术与种质资源研究所,云南 昆明 650223)
【研究意义】茶树[Camelliasinensis(L.)O.Kuntze]是多年生木本植物,在长期的自然演化和人工选择过程中,其遗传背景复杂。【前人研究进展】云南属于中国西南部,具有悠久的种茶历史和独特的气候条件,孕育了丰富的茶树资源,分布广泛,类型多样(包括地方品种、选育品种、育成品种、野生种及近缘植物等),被认为是茶树的原产地,是茶树原始起源中心与物种多样性中心。尤其地方品种和野生茶树资源种类较多,分布广泛。地方品种与野生茶树之间在物种类型、形态特征、生化成分和基因水平均存在丰富的多样性[1-3]。ISSR(inter-simple sequence repeat,简单重复间序列)标记具有操作简便、快捷、可靠性好等特点,并已应用许多作物遗传多样性评价、遗传图谱构建和遗传演化分析等[4]。目前,ISSR技术在茶树资源上已有很多研究[5-6]。【本研究切入点】本研究利用ISSR分子标记技术对云南茶树地方品种与野生茶树进行遗传多样性和亲缘关系分析。【拟解决的关键问题】从分子水平上探讨它们之间的亲缘关系,为更好地开展茶树资源的保护与利用、杂交亲本的选配及茶树的育种工作提供参考。
1 材料与方法
1.1 试验材料
云南茶树种质资源67份(表1),其中包括原产于云南保山品种10份,临沧品种23份(含14份野生茶树),德宏品种11份(含2份野生茶树),西双版纳品种8份,大理品种2份,普洱品种13份,均采自于国家种质勐海大叶茶树资源圃。每份材料取4 ~6个一芽二叶新梢。
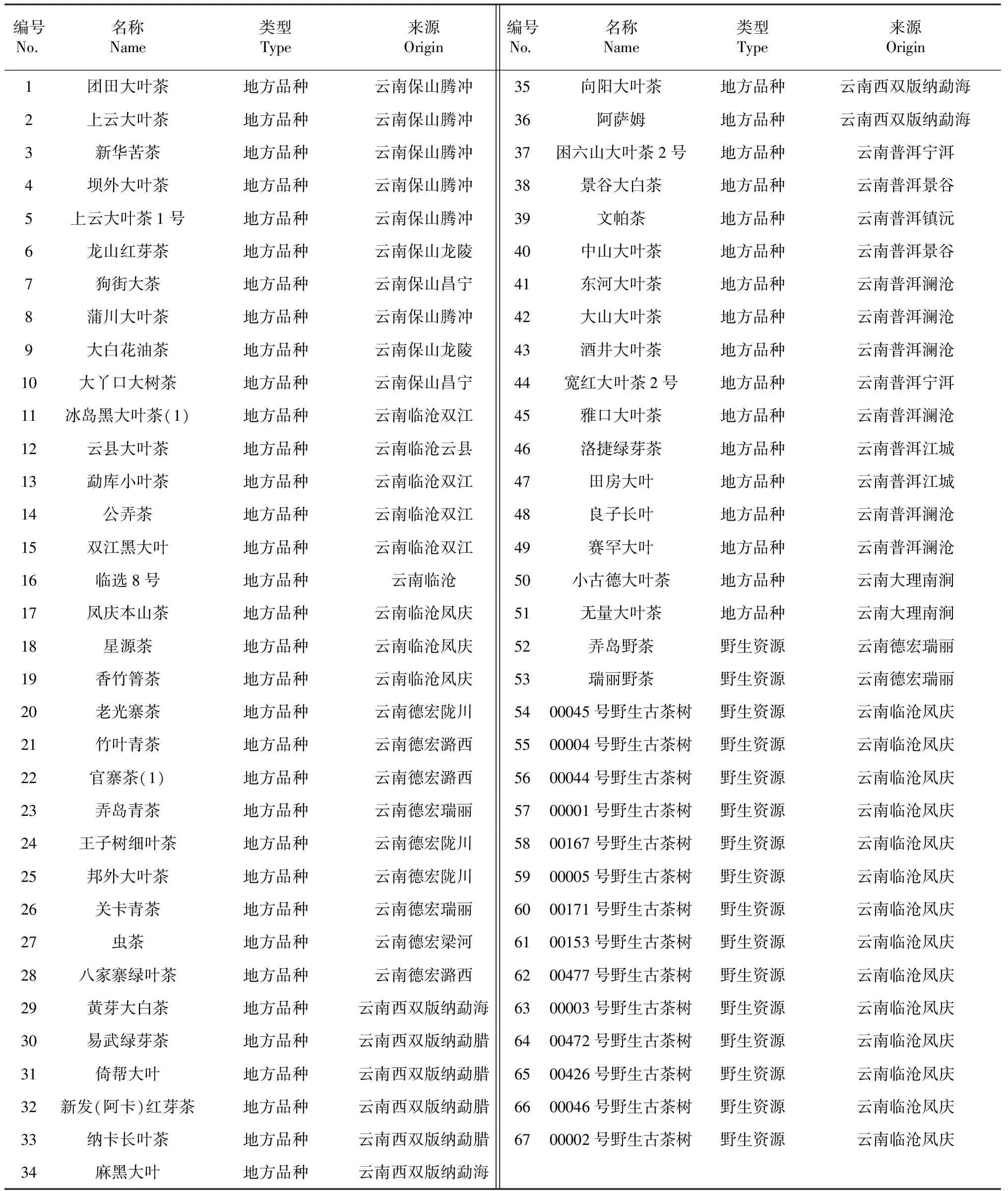
表1 67份云南茶树资源的名称及来源Table 1 Name and origin of 67 tea Yunnan plants
注:以上材料均保存于国家种质勐海大叶茶树资源圃。
Note: The above germplasms were all stored in Menghai National Germplasm Tea Repositories.
1.2 基因组DNA提取
采用改良CTAB法[7]提取供试材料基因组DNA,并用灭菌ddH2O稀释到50 ng/μl,4 ℃保存备用。
1.3 ISSR反应条件和产物检测
参照已有的茶树 ISSR- PCR 体系[8],反应体系总体积为15 μl,PCR组分如下:10×PCR buffer(含Mg2+)1.5 μl,10 nmol/L dNTP 0.3 μl, 5 UTaq酶0.2 μl,10 μmol/L引物0.5 μl,50 ng/μl 模板DNA 3 μl,ddH2O 9.5 μl。PCR扩增程序参照姚明哲方法[7]。PCR反应在96孔热循环仪(德国Biometra公司)上进行。PCR产物在1×TBE缓冲液中用1.5 %琼脂糖凝胶电泳1 h, 凝胶成像仪拍照记录。
1.4 数据分析
用人工读带方法,同一位置的条带在各材料中有带记1,无带则记为0,建立原始数据矩阵。每2份材料间采用Nei公式[9]计算其遗传相似系数(GS)和遗传距离(GD),GSij=2Nij/(Ni+Nj),GDij=1-GSij,其中Nij指2份材料共有的带数,Ni和Nj分别为i和j的扩增总条带数。使用Popgene_32软件分析材料间的基因多样性指数(H)、Shannon指数(Shannon index,I)和组间进化树等。使用软件NTSYS-pc version 2.1[10]进行聚类与主成分分析。
2 结果与分析
2.1 ISSR标记多态性
随机选取10份材料从42条ISSR引物中初步筛选出25条引物分别对全部材料进行扩增,最终筛选出23条引物扩增效果良好(表2),占所合成引物的54.7 %,图1为引物UBC848的扩增结果。使用23条引物对67份材料DNA进行PCR扩增,结果表明,扩增的多态性位点数从5~13不等,23条引物共扩增出223个条带,其中多态性条带数218个,占总条带数97.76 %,平均每个引物扩增多态位点9.5个,平均基因多样性指数(H)和Shannon信息指数(I),分别为0.27和0.42(表2)。
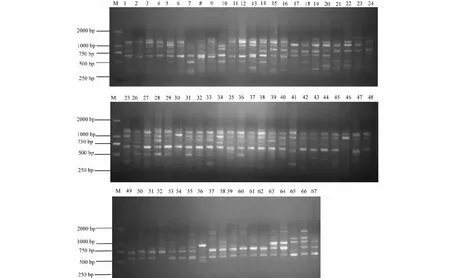
M为2000 bp DNA ladder分子量标准; 1~67分别为表1中的供试材料编号M:2000 bp DNA ladder marker; 1-67:No. of the tea accessions in the table 1图1 引物UBC848的扩增结果Fig.1 Amplication of primer UBC848
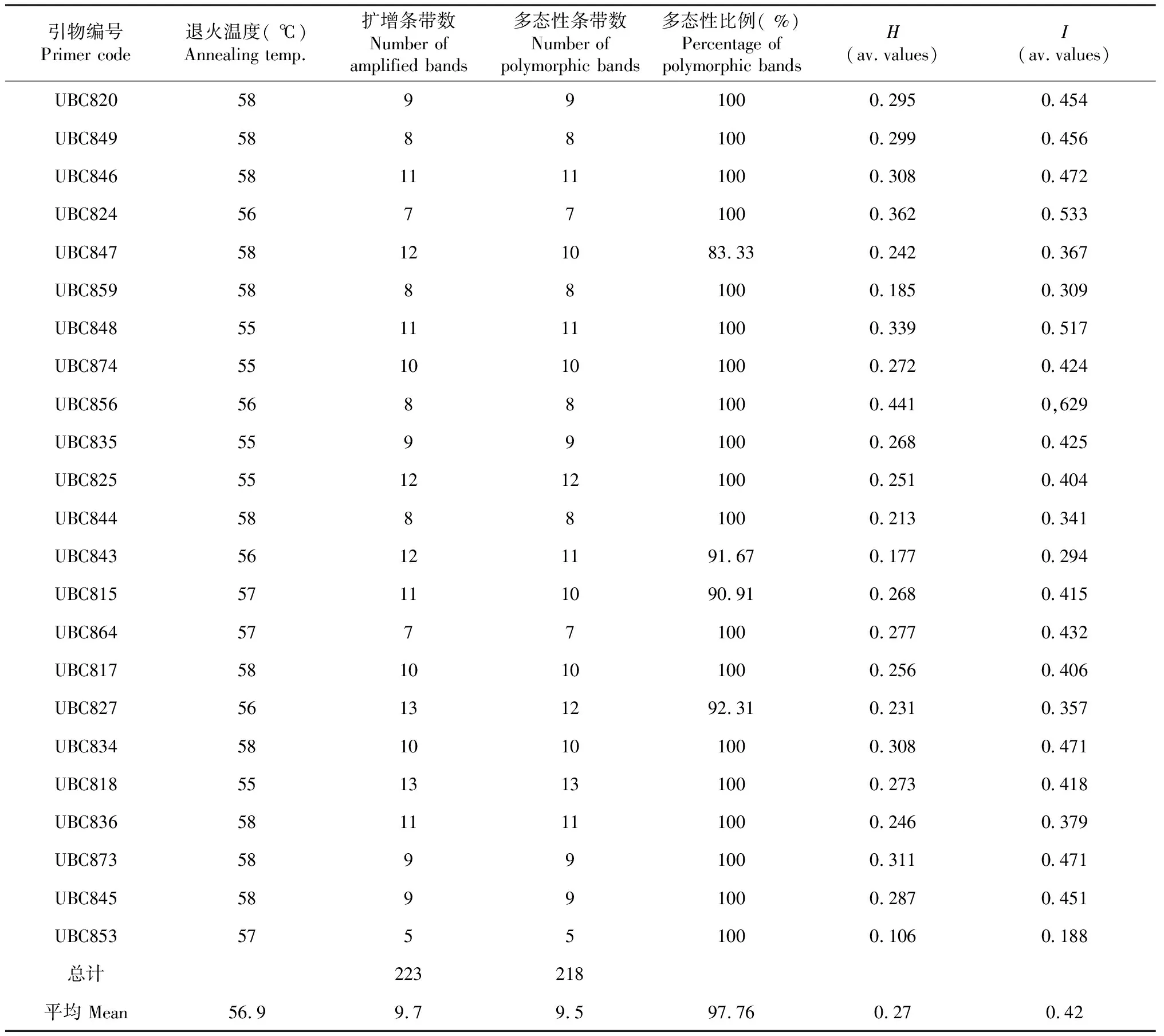
表2 引物扩增结果及多态性Table 2 Amplification result and polymorphism of primers
2.2 供试材料的遗传相似程度
使用软件NTSYS-pc version 2.1分析67份茶叶种质资源矩阵数据,计算出种质资源的遗传相似系数(表3)。它们之间的遗传相似系数变化在0.659~0.736,平均为0.693。普洱地方品种的平均遗传相似系数最高(0.736),表明其遗传多样性较低;而德宏野生茶树的平均遗传相似系数最低(0.659),表明其遗传多样性相对较高。从不同茶树种质的来源及类型上看,保山地方品种与临沧凤庆野生茶树的遗传相似程度最低,其次是临沧地方品种与其野生茶树,说明地方品种与野生茶树之间的亲缘关系较远;而普洱地方品种与德宏地方品种和大理地方品种的遗传程度高,其次是保山地方品种与德宏地方品种,说明来源不同的云南地方品种之间亲缘关系较近,遗传多样性也相对较低。
2.3 供试材料的聚类分析
根据算出的遗传相似系数矩阵,按UPGMA法对供试67份茶树种质进行聚类分析(图2)。在遗传相似系数0.666处(细线处),可将供试材料分为两大类:第I类55份品种,包括全部的地方品种和4份野生茶树(弄岛野茶、00477野生古茶树、00472号野生古茶树和00426野生古茶树),第II类为12份野生茶树;进一步划分(遗传相似系数0.666处)可将第I类组内分为3个亚组,第I 1亚组包括4份品种,为保山地方品种,第I 2亚组49份,其中47份为地方品种,而2份品种(弄岛野茶和00477野生古茶树)为野生茶树,第I 3亚组2份,分别为00472号和00426号野生茶树。第II类组内可分为2个亚组,第II 1亚组为瑞丽野茶单独聚为一类,第II 2亚组11份,均为野生茶树。从图2聚类关系图结果显示,来源于云南不同地方的茶树品种和多数野生茶树各自聚为一类,说明这些茶树品种的地理来源与遗传差异之间并没有明显的差异,而与其品种类型有一定关系,这与传统分类一致。
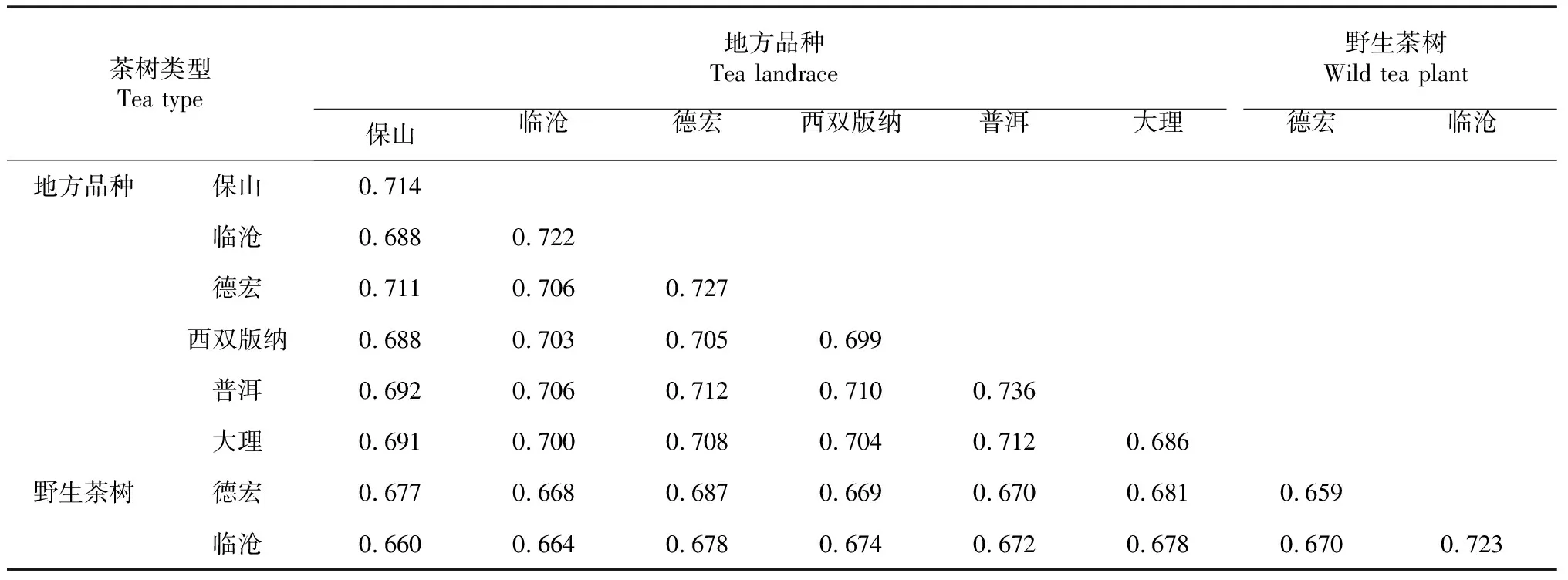
表3 不同来源茶树地方品种和野生品种的平均遗传相似系数Table 3 Average genetic similarity coefficient of different local varieties and wild species in tea
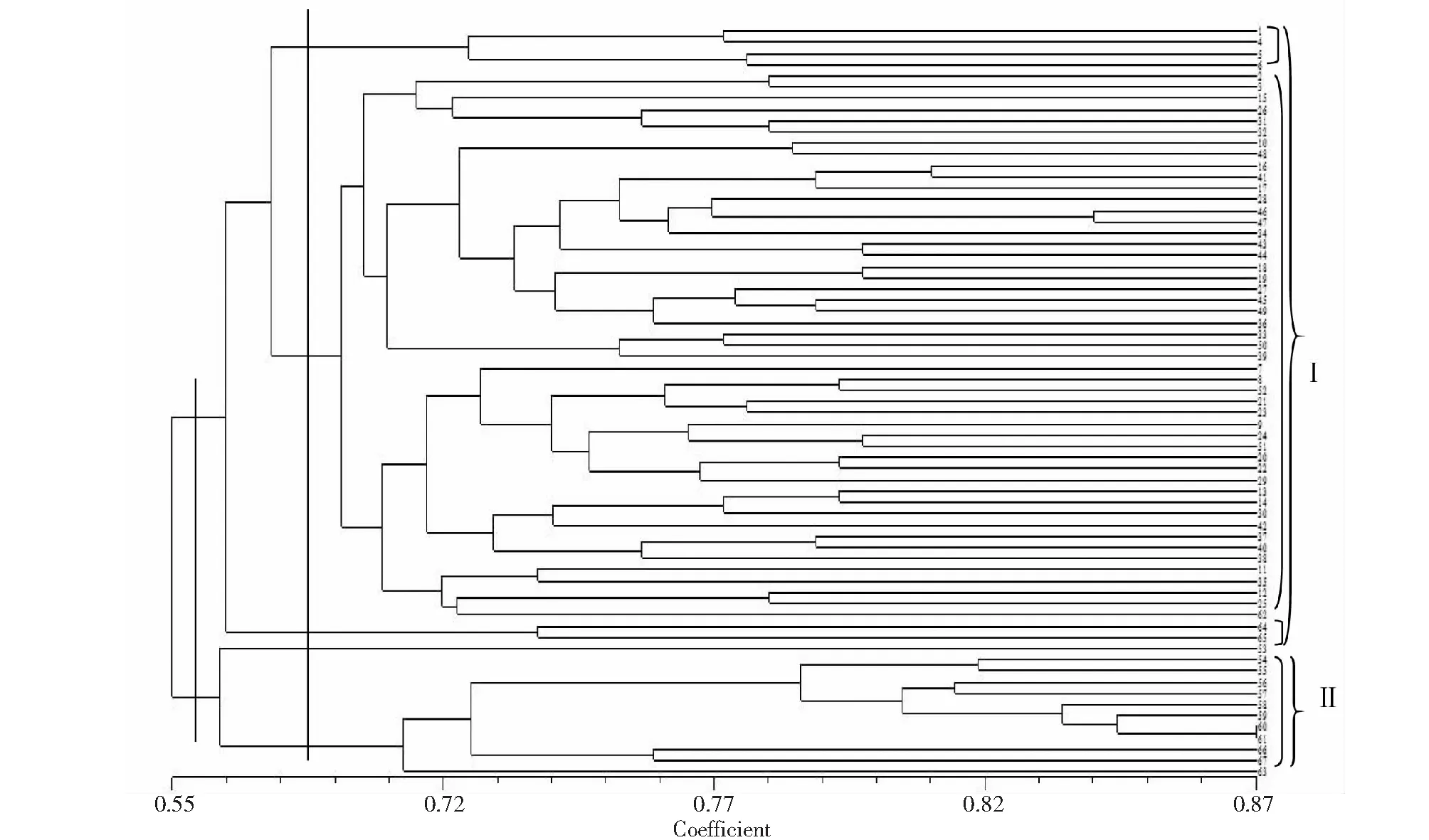
材料编号同表1 Material number was the same as table 1图2 67份茶树种质资源的聚类分析Fig.2 Dendrogram of 67 tea germplasm resources using UPGMA based on ISSR analysis
2.4 主成分分析
基于ISSR原始矩阵,利用NTSYS-pc version 2.1软件进行主成分分析(图3),位置相近者说明关系近,位置远离者则关系远。比较图2~3,主成分分析的结果与系统聚类的结果基本一致,其比聚类分析更直观地显示各个供试材料之间的亲缘关系。
3 讨 论
ISSR标记能够灵敏地揭示品种(系)间的差异,同时该技术所要求的引物较长(16~25 bp),退火温度较高,可重复性高[11]。吴清韩等[12]对48份凤凰单丛茶树资源遗传多样性和亲缘关系分析,多态位点比率为 81.0 %。Liu等[13]利用ISSR标记分析134份云南茶树资源,多态性比例高达98.9 %。本研究用23条引物对67份云南茶树资源进行多样性分析,扩增出223个条带,218个条带具有多态性,多态性比例为97.76 %,这些多态性主要是由于品种间的差异造成的,可以有效地反映品种间的亲缘关系。

图3 67份茶树种质资源的主成分分析Fig.3 Principle component analysis of 67 tea germplasm resources
遗传相似系数是用来比较不同品种或群体间遗传多样性的重要指标。本研究中,各品种间的相似系数变化在0.659~0.736,平均值为0.693,略高于姚明哲等[14]对36个无性系茶树品种和刘本英等[13]对134份云南茶树资源的研究结果,多数地方品种间的遗传相似系数高于野生资源的遗传相似系数,原因在于茶树是异花授粉植物,本身是杂合体,而云南茶树地方品种多数是大叶茶树资源,相互间可能存在基因交流的现象,导致地方品种间的遗传距离小、遗传多样性有降低趋势。周炎花等[15]对51份广西茶树地方品种的遗传多样性表明来自广西不同地区地方品种间的遗传多样性较低,本研究结果与其基本相一致。而云南野生资源遗传背景较复杂,根据周萌等[16]对100份云南野生茶树资源的遗传多样性分析,研究结果表明云南野生茶树的遗传多样性丰富。本研究由于客观条件等原因,所使用的野生茶树来源和数量较少,对试验结果可能有一定影响,但是多数野生茶树的遗传多样性比地方品种遗传多样性高,只有极少部分野生茶树的遗传相似系数略高于地方品种,可能与其复杂的遗传背景和农艺性状有关,有待进一步研究。
4 结 论
本研究聚类分析和主成分分析结果表明,来源不同的云南地方品种和少数野生茶树因地理来源相近和遗传背景相似而聚在同一类群中,而多数野生茶树由于遗传背景相对于其他品种较远聚在单独的一类。说明云南地方品种间的亲缘关系较近,遗传基础相对单一,其中少数野生茶树与地方品种聚为一类,它们可能与地方品种有着直接或间接的亲缘关系,具有相近的遗传背景。通过对云南地方品种与野生茶树遗传多样性的研究,可以为今后在充分利用地方品种的同时,要着重引入野生茶树或其他省份茶树的外源遗传变异,增加茶树种质资源的遗传多样性,培育和选育出优良的茶树新品种。
参考文献:
[1]蒋会兵, 宋维希, 李友勇, 等. 云南茶树种质资源的表型遗传多样性[J]. 作物学报, 2013, 39(11):2000-2008.
[2]陈 亮, 王平盛, 山口聪. 应用RAPD分子标记鉴定野生茶树种质资源研究[J]. 中国农业科学, 2002, 22(1): 19-24.
[3]马 玲, 蒋会兵, 何青元, 等. 云南文山古茶树资源调查和分析[J]. 西南农业学报, 2017, 30(8):1732-1738.
[4]Femamdez M E, Figueiras A M, Benito C. The use of ISSR and RAPD markers for detecting DNA polymorphism, genetype identification and diversity among barley cultivars with known origin[J]. Theor Appl Genet, 2002, 104:845-851.
[5]谭和平, 徐利远, 余桂蓉, 等. 茶树种质资源ISSR分子标记初步研究[J]. 核农学报, 2006, 20(2):113-115.
[6]姚明哲, 陈 亮, 王新超, 等. 我国茶树无性系品种遗传多样性和亲缘关系的ISSR分析[J]. 作物学报, 2007, 33(4):598-604.
[7]刘本英, 王平盛, 周红杰, 等. 云南茶树ISSR-PCR技术体系的建立[J]. 云南农业大学学报, 2006, 21(增刊) :21-25.
[8]姚明哲, 王新超, 陈 亮, 等. 茶树ISSR-PCR反应体系的建立[J]. 茶叶科学, 2004, 24(3):172-176.
[9]Nei M, Li W. Mathematical model for studying genetic variation in terms of restriction endonucleases[J]. Proceedings of the National Academy of Sciences of the United States of America,1979, 76:5269-5273.
[10]Rohlf F J. Statistical power comparisons among alternative morphometric methods[J]. Am J Phys Anthropol, 2000, 111: 463-478.
[11]胡邵庆, 邱英雄, 洪光洪, 等. 桂花品种的ISSR分析[J]. 南京林业大学学报(自然科学版), 2004, 28:71-75.
[12]吴清韩, 庄东红, 朱 慧, 等. 凤凰单丛茶树资源遗传多样性的ISSR分析[J]. 热带作物学报, 2015, 36(3):499-503.
[13]刘本英, 李友勇, 唐一春, 等. 云南茶树资源遗传多样性与亲缘关系的ISSR分析[J]. 作物学报, 2010, 36(3): 391-400.
[14]姚明哲, 陈 亮, 王新超, 等. 我国茶树无性系品种遗传多样性和亲缘关系的ISSR分析[J]. 作物学报, 2007,33(4): 598-604.
[15]周炎花, 乔小燕, 马春雷, 等. 广西茶树地方品种遗传多样性和遗传结构的EST-SSR分析 [J]. 林业科学, 2011, 47(3):59- 67.
[16]周 萌, 李友勇, 孙雪梅, 等. 基于EST-SSR标记的云南野生茶树遗传多样性分析[J]. 江苏农业科学,2013, 41(12):22-27.
猜你喜欢
区域治理(2022年40期)2022-11-27
今日农业(2022年14期)2022-09-15
今日农业(2022年13期)2022-09-15
茶道(2022年3期)2022-04-27
福建茶叶(2020年5期)2020-12-23
福建茶叶(2020年10期)2020-12-22
农技服务(2020年1期)2020-12-17
四川蚕业(2020年4期)2020-02-10
动漫界·幼教365(小班)(2019年10期)2019-10-28
动漫界·幼教365(大班)(2019年10期)2019-10-28
