
自然变温下小叶蚊母树叶片生理指标及组织结构变化
2018-06-05 10:54姜新强李璐璐刘庆超刘庆华王奎玲
河南农业科学 2018年4期
姜新强,李璐璐,刘庆超,刘庆华,王奎玲*
(1.青岛农业大学 园林与林学院,山东 青岛 266109; 2.北京林业大学 园林学院,北京 100083)
小叶蚊母树(Distyliumbuxifolium)是金缕梅科蚊母树属常绿小灌木,在亚热带地区分布广泛,其叶色浓绿,早春2—4月开花,花朵紫红色,花量大,观赏价值较高,广泛用于室内外园林绿化[1],已在南京等地成功引种栽培[2]。秋冬季节持续低温是小叶蚊母树在北方地区露地栽植应用的主要限制因子。低温能够破坏植物细胞结构和稳定性,造成植物体内代谢失调[3],影响正常的生长发育,严重时造成植物死亡。然而,当植物遭受低温胁迫时,植物可以通过改变其组织器官的形态结构[4]和多种代谢途径来抵抗所受到的低温胁迫,包括渗透保护物质的积累、抗氧化酶系统的激活以及抗逆蛋白质的合成等方面[5-6]。小叶蚊母树具有一定的抗寒性,其叶片半致死温度为-6.23 ℃[7]。明确小叶蚊母树叶片在低温下的生理适应机制对于揭示其耐寒机制和筛选抗寒种质资源具有十分重要的意义。鉴于此,以自杭州引种至青岛地区露地栽培的小叶蚊母树为试验材料,通过测定其在秋冬季节自然变温过程中不同时期叶片组织结构、叶片相对电导率和相关生理指标的变化,探究小叶蚊母树的抗寒性和对自然变温的适应机制,为其北引栽培和培育耐寒种质提供详实的科学依据和理论参考。
1 材料和方法
1.1 试验材料
供试材料为 2 年生小叶蚊母树植株,露地栽植于即墨市普东镇青岛农业大学实验基地中。2015年10月至2016年4月每月中旬在天气晴朗的上午10:00—12:00进行取样。在30株小叶蚊母树中选取生长健壮、受光均匀的3株植株,每株取向阳面枝条顶端向下第4节生长健壮的成熟叶片15片,随机分为3个重复,每个重复取5片叶片用于观察叶片组织结构。试验期间月平均最低温度分别为17、11、4、-2、-7、-2、3 ℃。
1.2 试验方法
1.2.1 相对电导率及生理指标测定 取1.1中叶片用于电导率和生理指标的测定。参照赵世杰等[8]的方法测定小叶蚊母树叶片相对电导率;叶片丙二醛(MDA)、可溶性蛋白和脯氨酸含量测定参考王学奎[9]的方法;超氧化物歧化酶(SOD)活性的测定参考Giannopolitis等[10]的方法,以每单位时间内抑制光还原50%的四氮唑蓝(NBT)为1个酶活性单位,5个生物学重复。
1.2.2 叶片组织结构观测 采用常规石蜡切片法观察小叶蚊母树叶片组织结构,用蒸馏水冲洗叶片,在主脉及两侧取0.5 cm×1.0 cm样品,保存于FAA固定液中,真空泵抽气后备用。切片厚度为8 μm,番红、固绿双重染色后用中性树胶封片,在Olympus CX31(CX31, Olympus, Japan)光学显微镜下观察,每个样品统计2个视野,每个月份统计30张照片。用Image J软件测量叶片中脉厚度,叶片厚度,上、下表皮细胞厚度,栅栏组织厚度,海绵组织厚度。参考Lianopoulou等[11]的方法,计算栅栏组织相对体积比,海绵组织相对体积比,上、下表皮细胞相对体积比。参考马建静等[12]的方法,计算细胞体积与细胞长宽比。
上表皮细胞相对体积比=上表皮细胞厚度/叶片厚度×100%,
下表皮细胞相对体积比=下表皮细胞厚度/叶片厚度×100%,
栅栏组织相对体积比=栅栏组织厚度/叶片厚度×100%,
海绵组织相对体积比=海绵组织厚度/叶片厚度×100%,
细胞体积= π·(细胞宽度/2)2×细胞长度(0.38+0.117×细胞长度/细胞宽度),
细胞长宽比=细胞长度/细胞宽度。
1.3 数据统计
所有数据均通过Excel 2013进行整理,利用SPSS 21.0进行差异显著性分析。采用单因素(one-way ANOVA) 和最小显著差异法(LSD)进行方差分析和多重比较(α=0.05)。
2 结果与分析
2.1 自然变温过程中的细胞膜透性的变化
由图1可知,2016年2月以前,随着月平均最低温度的降低,叶片相对电导率呈现升高的趋势,2016年2月(月平均最低气温为-7 ℃)叶片相对电导率最高,为58.90%,相比于2015年10月升高了276.88%,显著高于其他时期。2016年3月和4月,随着月平均最低温度的上升,小叶蚊母树叶片相对电导率下降,分别降至27.34%和23.23%。
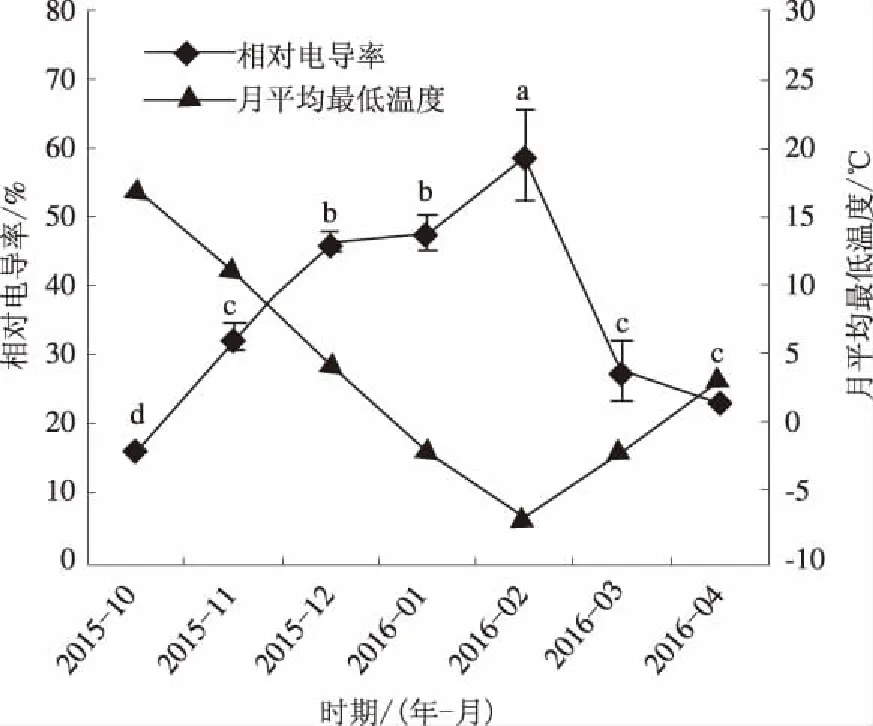
不同小写字母表示不同处理间差异显著(P≤0.05),下同
图12015年10月至2016年4月月平均最低温度和小叶蚊母树叶片相对电导率
2.2 自然变温过程中的叶片生理指标的变化
如图2所示,小叶蚊母树叶片SOD活性、MDA、可溶性蛋白和脯氨酸含量均呈现出先升高后降低的趋势。2016年2月和3月叶片SOD活性显著高于其他月份,2016年3月SOD活性分别为2015年10月和2016年4月的1.67和1.73倍;MDA和脯氨酸含量最大值均出现在2016年2月,分别为14.34 μmol/g和226.16 μg/g,均显著高于其他月份,MDA 含量最低值出现在2016年3月,为5.64 μmol/g,脯氨酸含量最低值为98.73 μg/g,出现在2015年10月;2016年1—2月的叶片可溶性蛋白含量显著高于其他时期。
2.3 自然变温过程中的叶片组织结构的变化
选取小叶蚊母树2015年10月、2016年1月和4月叶片对其组织结构进行切片观察,结果见图3。
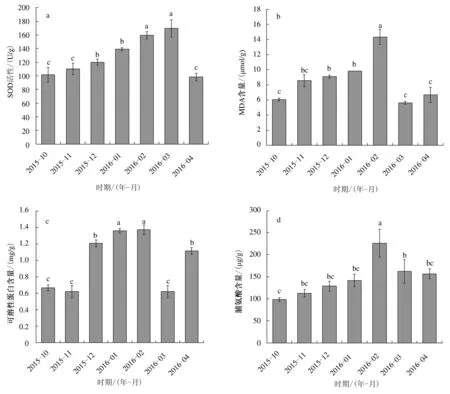
图2 2015年10月至2016年4月小叶蚊母树叶片生理指标的变化
由图3可以看出,小叶蚊母树叶片为不等面叶,不同时期叶片均由上表皮细胞、栅栏组织、海绵组织和下表皮细胞构成,维管束由木质部和韧皮部组成。叶脉维管束为半圆形,韧皮部外侧有木化的韧皮纤维。2015年10月叶片栅栏组织细胞排列较疏松(图3A、D),细胞间隙较大,2016年1月(图3B、E) 和4月(图3C、F) 叶片栅栏组织细胞排列较紧密。2016年1月叶脉皮层组织出现分层现象(图3B),细胞从排列最松散处分开且向四周扩展形成空隙(图3B、C框内),2016年4月空隙变小。
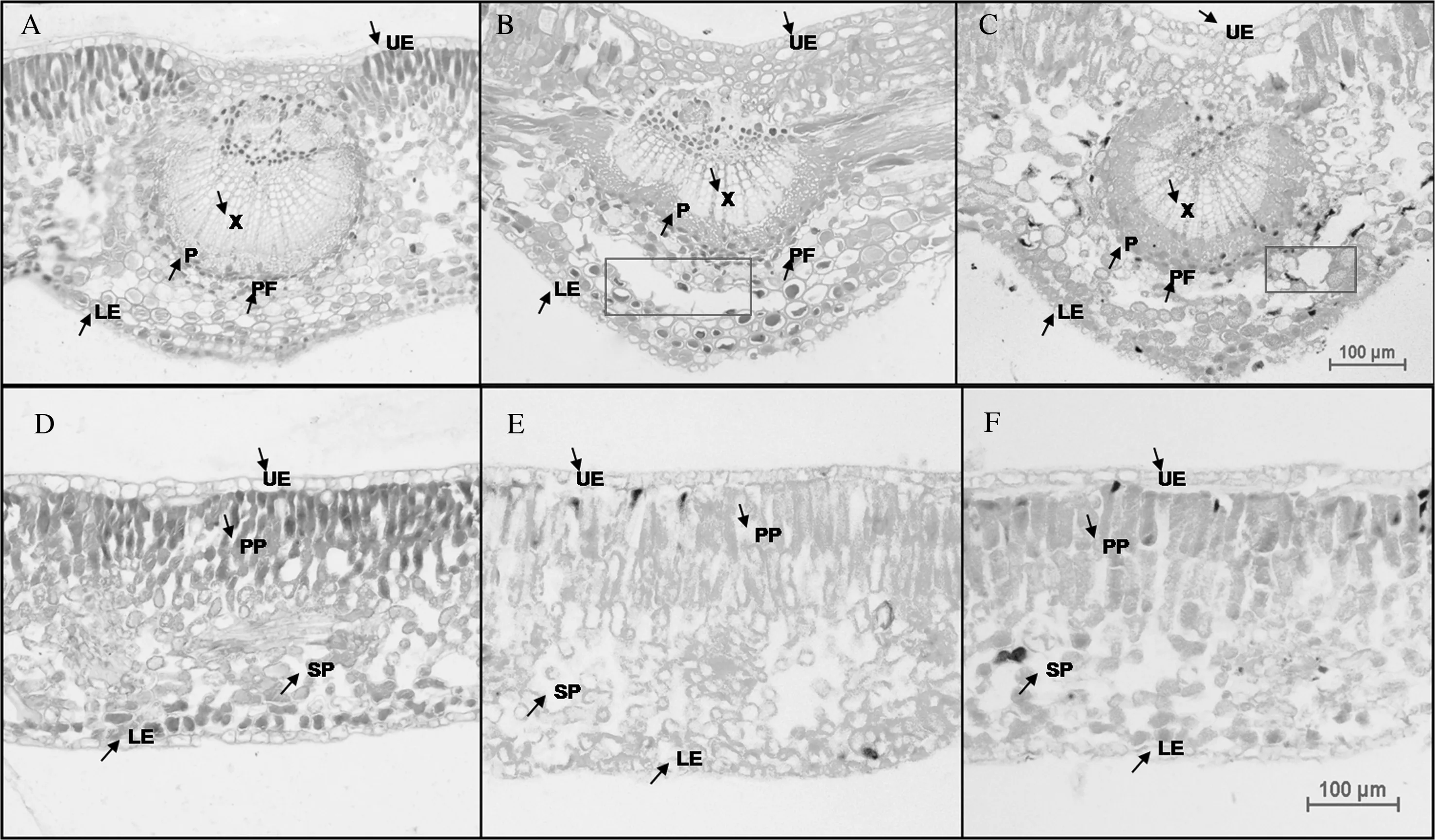
A—C 分别为2015年10月、2016年1月和2016年4月小叶蚊母树叶中脉解剖结构;D—F 分别为2015年10月、2016年1月和2016年4月小叶蚊母树叶叶肉解剖结构。X:木质部细胞; P:韧皮部细胞; UE:上表皮细胞; PP:栅栏组织细胞; PF:纤维细胞; SP:海绵组织细胞; LE:下表皮细胞
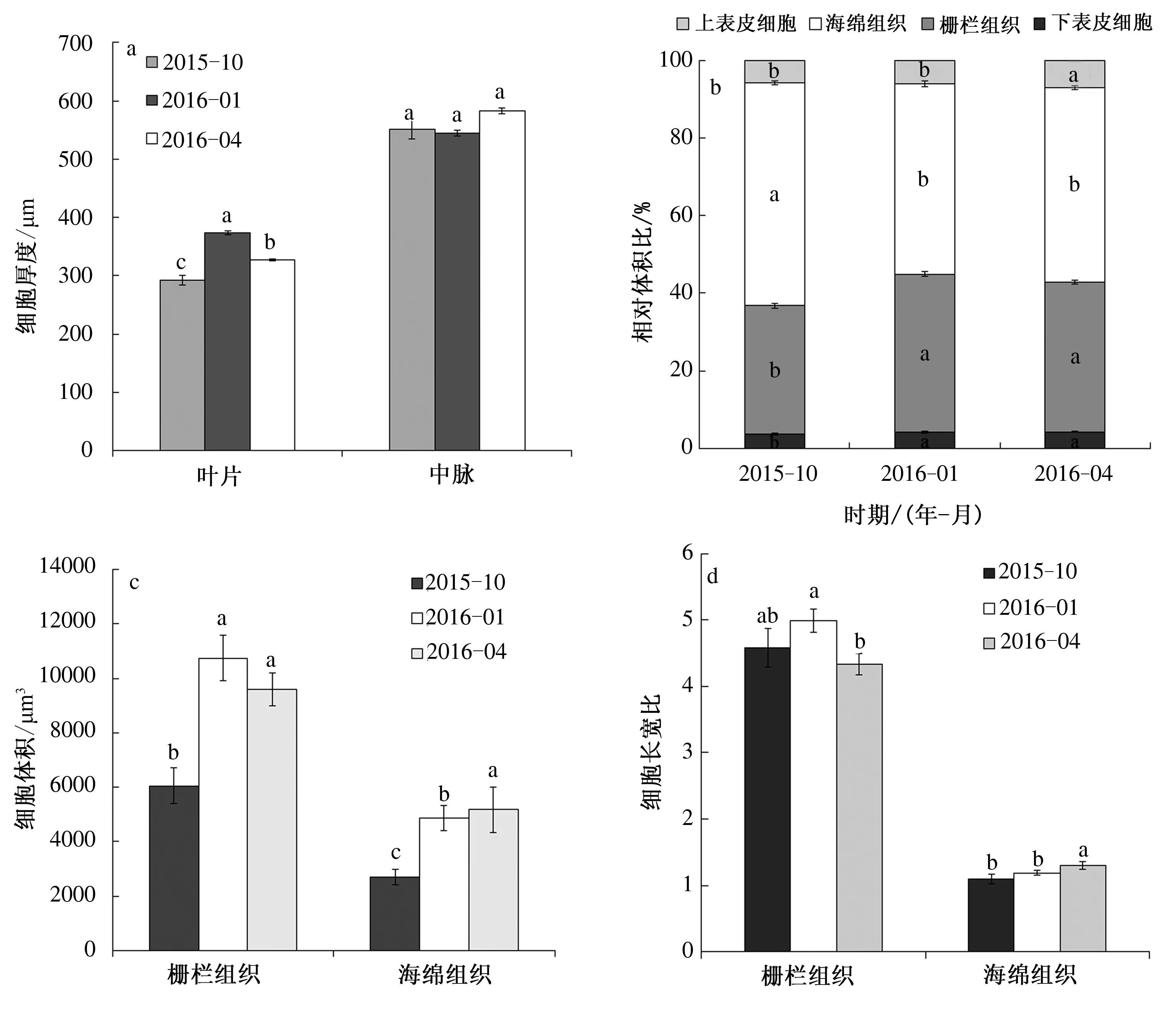
图4 自然变温过程中小叶蚊母树叶片解剖结构特征
对自然变温过程中小叶蚊母树叶片中脉厚度,叶片厚度,上、下表皮细胞厚度,栅栏组织厚度,海绵组织厚度,栅栏组织相对体积比,海绵组织相对体积比,上表皮细胞相对体积比,下表皮细胞相对体积比等11个指标进行了统计分析,结果如图4所示。2016年1月叶片厚度显著高于2015年10月和2016年4月,相比于2015年10月叶片厚度增加了27.7%,达到373.35 μm;2015年10月、2016年1月和2016年4月小叶蚊母树叶中脉厚度无显著性变化,最高值出现在2016年4月,为583.02 μm(图4a)。小叶蚊母树不同时期上、下表皮细胞、栅栏组织和海绵组织在叶片中所占体积不同(图4b)。2015年10月栅栏组织相对体积比显著低于2016年1月和4月,而海绵组织相对体积比显著高于2016年1月和4月;上表皮细胞相对体积比最大值出现在2016年4月,为7.1%,显著高于2015年10月(5.8%) 和2016年1月(6.0%),2016年1月和4月下表皮细胞相对体积比均为4.3%,显著高于2015年10月(3.7%);2016年4月海绵组织体积最大,为5 177.92 μm3(图4c),2016年1月和4月栅栏组织细胞体积显著高于2015年10月(图4c);栅栏组织细胞长宽比最大值出现在2016年1月(1.30),显著高于2016年4月(图4d),与2015年10月之间无显著性差异;2016年4月海绵组织细胞长宽比显著高于2015年10月和2016年1月(图4d)。
3 结论与讨论
温度与植物的生长、发育、开花、结实等生物学过程密切相关,是影响植物生长发育的重要环境因子,贯穿于植物生长的全部过程。在低温胁迫下,植物细胞和组织稳定性遭到破坏,叶片表现出萎篶、冻斑、焦枯,枝条变黑等受害症状[13]。但是经过短暂或长期的低温驯化后,植物也可以通过改变自身细胞、组织结构和生理代谢过程以增强细胞和组织稳定性,提高对低温环境的适应性。本研究对自然变温过程中的小叶蚊母树的叶片形态结构和生理指标变化进行了观测,发现其可以通过改变自身组织结构和积累生理代谢性物质以提高对低温环境的适应能力。
植物叶片是进行光合作用的主要器官,对温度敏感且可塑性较大。叶片可以通过自身组织结构的改变以增强对低温环境适应性[14-15]。研究表明,拟南芥叶片暴露在低温中15 d,叶片解剖结构就会发生不可逆的改变以适应低温环境[16]。对木本植物腰果(Anacardiumoccidentale)[17]、北海道黄杨(Euonymusjaponicus‘Cu Zhi’)[18]和油棕(Elaeisguineensis)[19]的研究均表明,低温能够使叶片结构出现适应性的变化,如叶片厚度增大,组织紧密度增加等以适应低温环境。本研究对小叶蚊母树2015年10月、2016年1月和4月叶组织结构不同参数的统计发现,在温度较低的2016年1月(平均最低温度-7 ℃),小叶蚊母树叶片厚度、栅栏组织细胞体积显著高于温度较高的2015年10月。随着温度的降低,小叶蚊母树叶片结构发生了对低温的适应性变化,并且叶片厚度的增加主要是由于叶片栅栏组织厚度增加。冬季(2016年1月)小叶蚊母树叶片厚度和栅栏组织在叶片中所占比重增大能增强低温环境中叶片对光能的利用效率以增加固碳能力,提高对低温逆境的抵抗能力。栅栏组织厚度的增加是小叶蚊母树适应低温环境的一种适应性表现,这与Poorter等[20]和Scheepens等[21]的研究结果一致。除细胞结构发生改变外,植物叶片对低胁迫的适应性还体现在叶肉细胞生长模式和细胞尺寸上。Stefanowska等[22]研究表明,欧洲油菜(Brassicanapus)叶片在零度以上的低温胁迫下,栅栏组织会由平行于叶表面方向伸长转向垂直于叶表面方向伸长,叶肉细胞体积也会增大。小叶蚊母树不同时期叶肉细胞体积与长宽比变化趋势一致,由此推测冬季植物叶片栅栏组织细胞体积的增大原因可能是由于栅栏组织细胞垂直于叶片方向的生长。
除叶片组织结构之外,植物还可以通过改变自身生理代谢物质以增强自身抗寒性。叶片相对电导率和MDA含量是衡量植物抗寒性的重要指标,相对电导率反映了细胞电解液泄露程度,MDA是细胞膜过氧化的产物,其含量可以反映细胞膜受害程度。2015年11月叶片 MDA 含量显著高于2015年10月,并且在2015年10月至2016年2月期间维持在较高水平,说明在此期间细胞膜受到损伤,MDA 积累。2016年2月小叶蚊母树叶片相对电导率和MDA含量达到最大值,表明在此期间细胞膜受害程度和细胞电解液外泄程度最高,细胞受害最严重。
植物在遭受低温、干旱等非生物胁迫时,会积累渗透调节物质,主动提高细胞对不良环境的抵抗能力[11]。在低温胁迫时,细胞内活性氧含量升高,造成细胞膜过氧化,破坏细胞膜结构和功能[23]。而SOD能有效清除细胞体内产生的活性氧[24],保护细胞膜结构和功能的稳定性。在自然变温期间,小叶蚊母树叶片SOD活性呈先升高后降低的趋势,温度比较低的2016年2月和3月 SOD 活性显著高于其他时期,表明在此时期细胞对活性氧的清除能力提高,细胞自身的生理代谢能力增强;2016年3月MDA含量显著降低,这可能与3月小叶蚊母树叶片中SOD活性较高有关,较高活性的SOD降低了细胞膜脂质过氧化水平。可溶性蛋白含量的增加有利于提高细胞内束缚水含量,从而降低冰点[25]。2016年1月和2月叶片可溶性蛋白含量显著高于其他时期,这是由于植物通过叶片中可溶性蛋白的积累以增加防止细胞内结冰的能力。作为细胞内的渗透调节物质,在低温胁迫下,可溶性蛋白和脯氨酸含量的增加可以增加细胞液浓度,降低细胞内结冰温度[26-27]。除了具有维持细胞内渗透压、结合蛋白质以保护蛋白活性的功能外,脯氨酸还具有清除羟基自由基、保护细胞膜的功能[28]。对冬青(Ilexchinensis)[25]和樱桃(Cerasuspseudocerasus)[29]的研究表明,叶片内脯氨酸含量在低温胁迫下均升高,与本研究结果一致。小叶蚊母树脯氨酸含量的最大值出现在2016年2月,显著高于其他时期,表明在此时期脯氨酸作为一种渗透条件物质,能够提高细胞液浓度,减弱低温对细胞的伤害。
本研究通过揭示小叶蚊母树在北方地区秋冬季节自然变温过程中叶片结构和生理指标的变化,明确了小叶蚊母树叶片能够通过改变组织结构、主动积累渗透调节物质和提高抗氧化物酶活性等机制提高自身对低温环境的适应能力,揭示了小叶蚊母树叶片越冬过程中的低温适应机制。但植物对低温的适应性状是数量性状,并且露地栽培过程中多种环境因素都会对其造成影响,对小叶蚊母树在引种地仅进行了1 a的低温适应性观测,未来还需要对引种植株进行更为全面的观测,以明确其在低温下的适应性。
参考文献:
[1] 何小勇,杨先裕,练发良.小叶蚊母树冬季不同叶龄叶色及其光合特性研究[J].浙江林业科技,2015,35(5):6-10.
[2] 谢晓金,郝日明.南京地区引种的24种常绿阔叶树种耐寒性比较[J].浙江林学院学报,2006,23(3):285-289.
[3] 兰淑芝,李伯清.低温对植物的影响[J].吉林农业,1997(8):19-20.
[4] England J R,Attiwill P M.Changes in stomatal frequency,stomatal conductance and cuticle thickness during leaf expansion in the broad-leaved evergreen species,Eucalyptusregnans[J].Trees,2011,25(6):987-996.
[5] 王达,马纪.植物耐寒机理与耐寒植物新品种培育[J].生物技术通报,2012(1):7-13.
[6] Foyer C H,Descourvières P,Kunert K J.Protection against oxygen radicals: An important defence mechanism studied in transgenic plants[J].Plant Cell and Environment,1994,17(5):507-523.
[7] 张鹏翀,王挺,刘锦,等.电导法配合Logistic方程测定5种高架绿化植物的抗寒性[J].广东农业科学,2013,40(19):49-50.
[8] 赵世杰,史国安,董新纯.植物生理学实验指导[M].北京:中国农业科学技术出版社,2002.
[9] 王学奎.植物生理生化实验原理和技术[M].2版.北京:高等教育出版社,2006.
[10] Giannopolitis C N, Ries S K. Superoxide dismutaseⅡ. Purification and quantitative relationship with water-soluble protein in seedlings[J].Plant Physiology,1997,59(2):315-318.
[11] Lianopoulou V,Bosabalidis A M,Patakas A,etal.Effects of chilling stress on leaf morphology, anatomy, ultrastructure, gas exchange,and essential oils in the seasonally dimorphic plantTeucriumpolium(Lamiaceae)[J].Acta Physiologiae Plantarum,2014,36(8):2271-2281.
[12] 马建静,吉成均,韩梅,等.青藏高原高寒草地和内蒙古高原温带草地主要双子叶植物叶片解剖特征的比较研究[J].中国科学(生命科学),2012,42(2):158-172.
[13] 李璐璐,姜新强,刘庆超,等.茶梅叶片结构对自然变温的适应[J].应用生态学报,2016,27(9):2815-2822.
[14] 李芳兰,包维楷.植物叶片形态解剖结构对环境变化的响应与适应[J].植物学通报,2005,22(S1):118-127.
[15] 覃凤飞,李强,崔棹茗,等.越冬期遮阴条件下3个不同秋眠型紫花苜蓿品种叶片解剖结构与其光生态适应性[J].植物生态学报,2012,36(4):333-345.
[16] Gebrueder B, Berlin F C, Descourivesse P,etal. Protection against oxygen radicals:An important defense mechanism studied in transgenic plants[J].Plant Cell Environ,1994,17(5):507-523.
[17] 梁李宏,梅新,林锋,等.低温胁迫对腰果幼苗叶片组织结构和生理指标的影响[J].生态环境学报,2009,18(1):325-328.
[18] 马妮,孙振元,刘庆华,等.北海道黄杨叶片解剖结构的季节变化[J].北京林业大学学报,2011,33(6):112-118.
[19] 曹红星,黄汉驹,雷新涛,等.不同低温处理对油棕叶片解剖结构的影响[J].热带作物学报,2014,35(3):454-459.
[20] Poorter H,Niinemets U,Poorter L.Causes and consequences of variation in leaf mass per area (LMA):A meta-analysis [J].New Phytologist,2009,182(3):565-588.
[21] Scheepens J F,Frei E S,Stöcklin J.Genotypic and environmental variation in specific leaf area in a widespread Alpine plant after transplantation to different altitudes[J].Oecologia,2010,164(1):141-150.
[23] 康菊清,张岱鹏.低温条件下中国野生拟南芥种群中CBF3与ROS浓度的相关性[J].植物学报,2016,51(5):577-585.
[24] 张玲玲,孙芳芳,温达志.薇甘菊和飞机草抗氧化物和膜脂过氧化物对光照和土壤水分的响应[J].植物生态学报,2009,33(5):974-983.
[25] 曾雯,金晓玲,邢文,等.9个常绿杂交冬青的抗寒能力比较[J].植物生理学报,2016,52(1):55-61.
[26] 周茹宝,张有福,张满效,等.两种圆柏属植物叶片代谢产物季节变化与抗冻性的关系[J].冰川冻土,2008,30(2):351-355.
[27] 王以斌,缪锦来,姜英辉,等.脯氨酸和可溶性糖在南极冰藻低温适应机制中的作用[J].生物技术通报,2016,32(2):198-202.
[28] Walker D J,Romero P,Correal E.Cold tolerance,water relations and accumulation of osmolytes inBituminariabituminosa[J].Biologia Plantarum,2010,54(2):293-298.
[29] 王敏,陈秋芳,石美娟,等.低温胁迫对甜樱桃品种枝条SOD、POD活性和脯氨酸含量的影响[J].山西果树,2011(6):3-5.
猜你喜欢
High Technology Letters(2021年4期)2022-01-09
数学小灵通(1-2年级)(2021年10期)2021-11-05
山东林业科技(2021年4期)2021-08-31
今日农业(2020年20期)2020-11-26
河北果树(2020年4期)2020-11-26
福建林业科技(2020年3期)2020-10-25
现代园艺(2020年17期)2020-08-31
海峡姐妹(2019年1期)2019-03-23
小说月刊(2015年2期)2015-04-23
第二课堂(小学版)(2014年4期)2014-08-02
