
不同居群新疆野苹果生理响应差异与抗寒性的关系
2018-05-31 01:47王泽华
经济林研究 2018年2期
王泽华 ,秦 伟
(1.新疆农业大学 林业研究所,新疆 乌鲁木齐 830052;2.湖南省农业委员会 湖南农业杂志社,湖南 长沙 410005)
苹果具有很高的经济价值,也是重要的水果资源、观赏资源及砧木资源,而中国的苹果产量和栽培面积均居世界首位[1]。在苹果属植物中,与栽培苹果亲缘关系最近的野生种是新疆野苹果[2-4],Richards等人研究认为,中亚地区是栽培苹果的起源中心,他们提出,新疆野苹果是栽培苹果的祖先种[5-6]。新疆野苹果是第三纪孑遗物种[7],主要分布在天山山脉,包括中国的新疆维吾尔自治区和哈萨克斯坦的阿拉木图州与塔尔迪库尔干州以及吉尔吉斯斯坦的伊塞克湖州等地区[2]。我国最常用的苹果砧木分别为山定子、西府海棠和新疆野苹果,其中的山定子抗逆性好但不耐盐碱,西府海棠亲和力好但抗逆性不如新疆野苹果,而新疆野苹果具有抗寒性、抗旱性、抗盐碱、抗病虫[9]多种优良的抗逆性状[8],且其与栽培苹果有较好的亲和力,时至今日,新疆野苹果已经成为我国西部地区的重要砧木资源[10]。目前,对植物生理生化指标与抗寒性关系的研究报道有很多[11-14],但有关低温胁迫下不同居群新疆野苹果生理响应差异的研究报道却较少,在之前的研究中,本课题组研究了低温胁迫下不同居群新疆野苹果休眠枝条的生理响应差异[15-16]。为给新疆野苹果优良抗寒种质资源的筛选提供参考依据,本研究以不同居群新疆野苹果实生苗的叶片为试材,对其低温生理响应差异进行了比较分析,现将研究结果分析报道如下。
1 材料与方法
1.1 材 料
2015年12月下旬分别在新疆维吾尔自治区伊犁州新源县的阿勒马勒乡、巩留县莫乎尔乡、霍城县大西沟乡、托里县多拉特乡、额敏县霍吉尔特乡等5个新疆野苹果自然居群采样,在每个自然居群内随机抽样带回实验室,待果实后熟后剥取种子,次年1月每个居群各取200粒种子进行层积处理,4月将层积处理好的种子播种于营养袋中,每袋播种4粒种子,均置于新疆农业大学校内实验田中让其自然生长,8月选取生长良好且无病虫害的植株进行低温处理,处理温度分别设为25(即室温,作为对照CK)、10、5、0、-5和-10 ℃,降温的速度和解冻时温度的回升速度均为4 ℃/h,降温时温度达到处理温度后维持24 h,然后解冻,达到室温后取出静置1 h,取中上部的叶片以测定相关指标。
1.2 方 法
采用电导法测定相对电导率,采用蒽酮比色法测定可溶性糖含量,采用茚三酮显色法测定脯氨酸含量,采用硫代巴比妥酸显色法测定丙二醛含量,采用考马斯亮蓝法测定可溶性蛋白含量,采用NBT光化还原法测定SOD活性,采用愈创木酚法测定POD活性,采用紫外吸收法测定CAT活性[17-18]。所有指标的测定均设3次重复。
1.3 数据处理
采用Excel 2010和SPSS 19.0软件进行数据处理与显著性分析。
2 结果与分析
2.1 不同低温胁迫对不同居群新疆野苹果相对电导率的影响
不同低温胁迫下不同居群新疆野苹果的相对电导率如图1 所示。由图1可知,在低温胁迫下,不同居群新疆野苹果的相对电导率均随着处理温度的下降而增加,但增加幅度不同。在处理温度降至0 ℃前,不同居群新疆野苹果相对电导率的增势均较缓;当温度降至0 ℃时,新源县、巩留县和霍城县野苹果相对电导率的增幅均较大,均超过10%,而托里县和额敏县野苹果的相对电导率在-5 ℃时才有较大增幅,这说明在处理温度降到0 ℃时,新源县、巩留县和霍城县野苹果细胞膜均受到明显伤害,而在处理温度降到-5 ℃时,托里县和额敏县野苹果细胞膜才受到明显伤害。显著性差异分析结果表明,在处理温度降到5 ℃后,新源县、巩留县和霍城县野苹果的相对电导率一直极显著高于托里县和额敏县野苹果的相对电导率(P<0.01)。
2.2 不同低温胁迫对不同居群新疆野苹果枝条可溶性糖含量的影响
不同低温胁迫下不同居群新疆野苹果枝条的可溶性糖含量如图2 所示。由图2可知,在低温胁迫下,不同居群新疆野苹果枝条的可溶性糖含量均呈先增后降的变化趋势:不同居群新疆野苹果的可溶性糖含量在处理温度降至0 ℃时均达到最大值,托里县、额敏县、巩留县、霍城县、新源县野苹果枝条的可溶性糖含量最大值由高到低依次为4.55%、4.33%、3.85%、3.72%、3.47%;与对照(CK)比较,托里县、额敏县、巩留县、新源县、霍城县野苹果枝条可溶性糖含量达到最高时的增量由多到少分别为2.13%、2.01%、1.11%、0.87%、0.84%。显著性差异分析结果表明,当处理温度为0 ℃时,托里县野苹果枝条的可溶性糖含量显著高于额敏县野苹果(P<0.05),托里县和额敏县野苹果枝条的可溶性糖含量差极显著高于其他3个居群(P<0.01),其他3个居群间的含量差异不显著(P>0.05)。

图1 不同低温胁迫下不同居群新疆野苹果的相对电导率Fig.1 Relative electrical conductivity of different Malus sieversii populations under different treatment temperatures
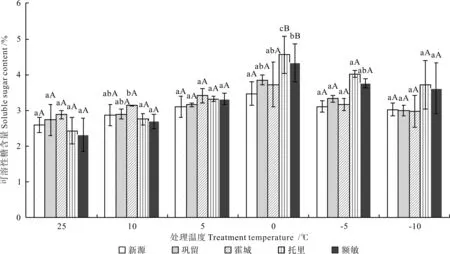
图2 不同低温胁迫下不同居群新疆野苹果枝条的可溶性糖含量Fig.2 Soluble sugar content of different Malus sieversii populations under different treatment temperatures
2.3 不同低温胁迫对不同居群新疆野苹果枝条脯氨酸含量的影响
不同低温胁迫下不同居群新疆野苹果枝条的脯氨酸含量如图3 所示。由图3可知,在低温胁迫下,不同居群新疆野苹果的脯氨酸含量均随处理温度的下降而升高。与对照(CK)比较,不同居群新疆野苹果的脯氨酸含量达到最高值时的增量由多到少依次为:托里县野苹果(69.63 µg·g-1)、额敏县野苹果(52.69 µg·g-1)、巩留县野苹果(41.51 µg·g-1)、霍城县野苹果(40.15 µg·g-1)、新源县野苹果(29.14 µg·g-1)。显著性差异分析结果表明,在处理温度降至0 ℃后,托里县野苹果的脯氨酸含量显著高于新源县野苹果(P<0.05),其余居群间的含量差异不显著(P>0.05)。
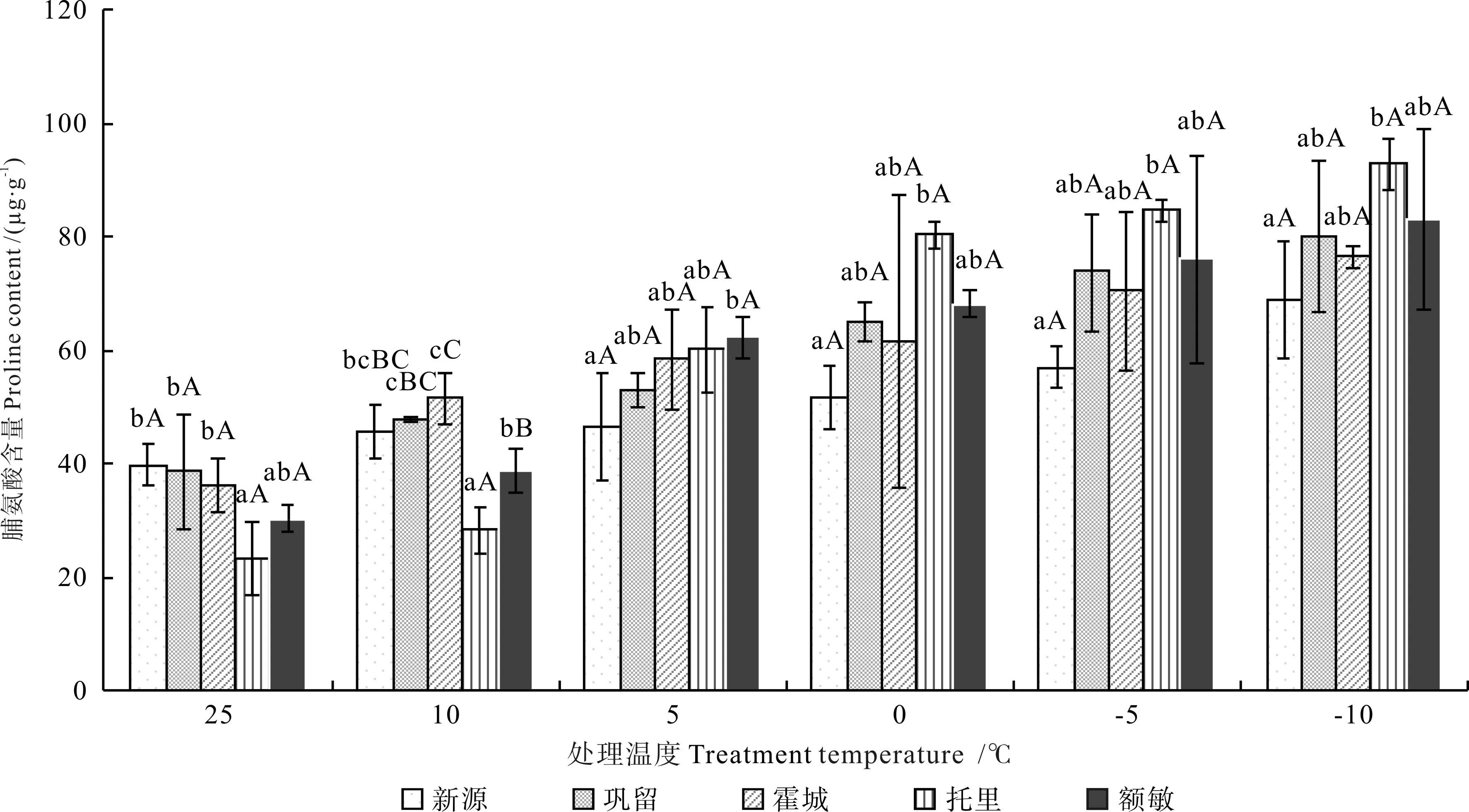
图3 不同低温胁迫下不同居群新疆野苹果枝条的脯氨酸含量Fig.3 Proline content of different Malus sieversii populations under different treatment temperatures
2.4 不同低温胁迫对不同居群新疆野苹果枝条丙二醛含量的影响
不同低温胁迫下不同居群新疆野苹果枝条的丙二醛含量如图4 所示。由图4可知,在低温胁迫下,不同居群新疆野苹果枝条的MDA含量均呈现先升后降的变化趋势,在处理温度降至-5 ℃时均达到最大值,其由高到低依次为:巩留县野苹果(46.02 μmol·g-1)、新源县野苹果(43.10 μmol·g-1)、霍城县野苹果(42.64 μmol·g-1)、额敏县野苹果(41.59 μmol·g-1)、托里县野苹果(39.22 μmol·g-1)。与对照(CK)比较,不同居群新疆野苹果枝条的MDA含量达到最高时的增量由多到少依次为:新源县野苹果(33.81 μmol·g-1)、巩留县野苹果(32.67 μmol·g-1)、霍城县野苹果(31.90 μmol·g-1)、额敏县野苹果(29.02 μmol·g-1)、托里县野苹果(27.45 μmol·g-1)。显著性差异分析结果表明,当处理温度为-5 ℃时,巩留县野苹果枝条的MDA含量显著高于托里县野苹果(P<0.05),而其他居群间的含量差异不显著(P>0.05)。
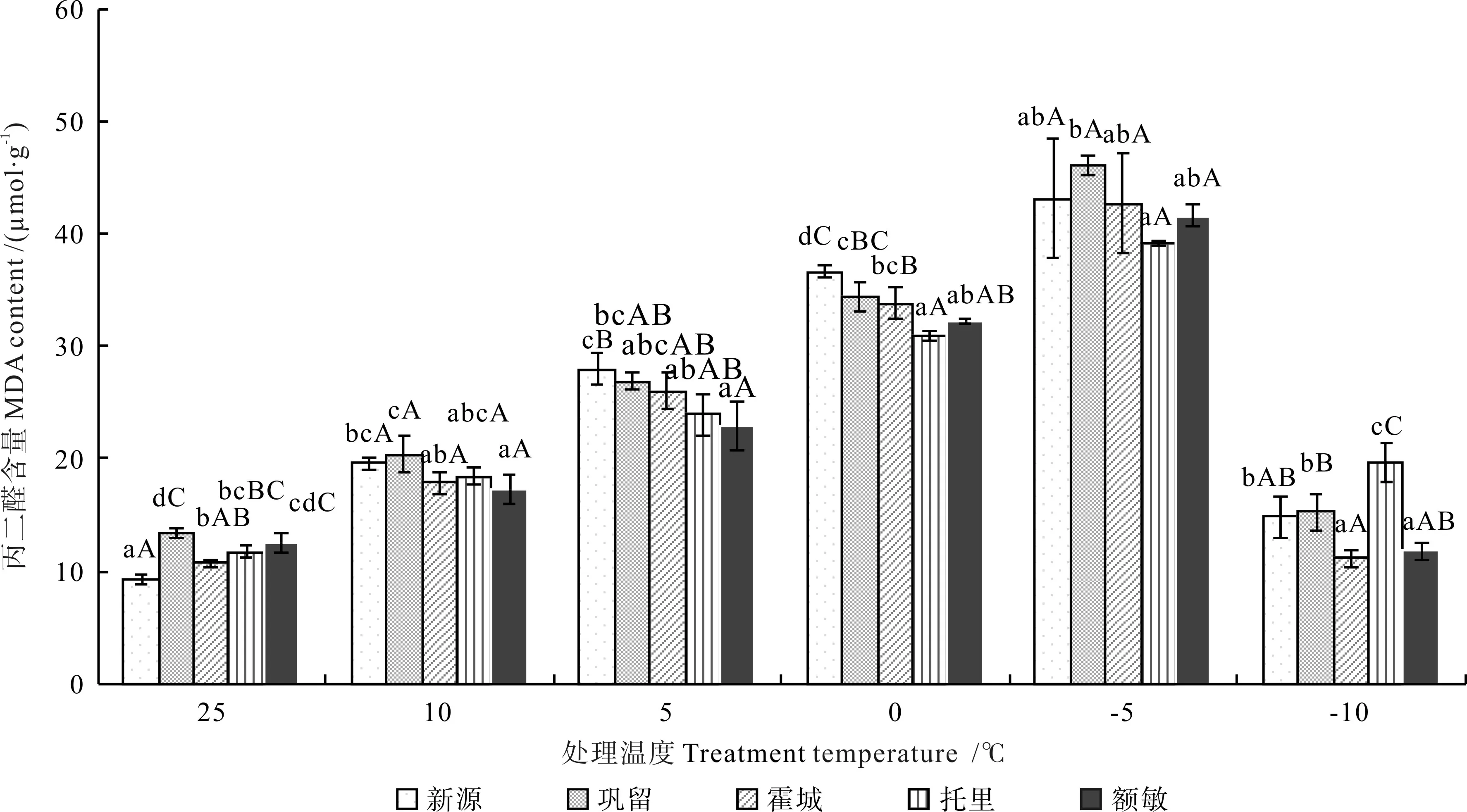
图4 不同低温胁迫下不同居群新疆野苹果枝条的丙二醛含量Fig.4 MDA of different Malus sieversii populations under different treatment temperatures
2.5 不同低温胁迫对不同居群新疆野苹果枝条可溶性蛋白的影响
不同低温胁迫下不同居群新疆野苹果枝条的可溶性蛋白含量如图5 所示。由图5可知,在低温胁迫下,不同居群新疆野苹果枝条的可溶性蛋白含量均随着处理温度的下降而升高。与对照(CK)比较,不同居群新疆野苹果枝条的脯氨酸含量达到最高值时的增量由多到少依次为:托里县野苹果(5.84 mg·g-1)、额敏县野苹果(5.59 mg·g-1)、霍城县野苹果(4.64 mg·g-1)、巩留县野苹果(4.58 mg·g-1)、新源县野苹果(4.33 mg·g-1)。显著性差异分析结果表明,当处理温度为-10 ℃时,托里县野苹果的可溶性蛋白含量极显著低于新源县、巩留县和霍城县野苹果(P<0.01),而霍城县野苹果的可溶性蛋白极显著高于其他4个居群(P<0.01)。

图5 不同低温胁迫下不同居群新疆野苹果枝条的可溶性蛋白含量Fig.5 Soluble protein content of different Malus sieversii populations under different treatment temperatures
2.6 不同低温胁迫对不同居群新疆野苹果枝条SOD活性的影响
不同低温胁迫下不同居群新疆野苹果枝条的SOD活性如图6 所示。由图6可知,在低温胁迫下,不同居群新疆野苹果的SOD活性均呈先增后降的变化趋势,在处理温度降至0 ℃时均达到最大值,这说明供试材料在温度降至0 ℃前其细胞自身保护能力增强,而低于0 ℃这种保护能力则变弱。不同居群新疆野苹果的SOD活性达到最高值时其由高到低依次为:托里县野苹果(180.59 U·g-1h-1)、额敏县野苹果(169.78 U·g-1h-1)、霍城县野苹果(160.40 U·g-1h-1)、巩留县野苹果(155.87 U·g-1h-1)、新源县野苹果(126.57 U·g-1h-1)。与对照(CK)比较,不同居群新疆野苹果的SOD活性达到最高时的增量由多到少依次为:托里县野苹果(149.16 U·g-1h-1)、额敏县野苹果(139.04 U·g-1h-1)、巩留县野苹果(131.83 U·g-1h-1)、霍城县野苹果(129.54 U·g-1h-1)、新源县野苹果(90.38 U·g-1h-1)。显著性差异分析结果表明,当处理温度为0 ℃时,托里县和额敏县野苹果的SOD活性均显著高于新源县野苹果(P<0.05),而在其他处理温度下,各居群间的含量差异均不显著(P>0.05)。
2.7 不同低温胁迫对不同居群新疆野苹果枝条POD活性的影响
不同低温胁迫下不同居群新疆野苹果枝条的POD活性如图7 所示。由图7可知,在低温胁迫下,不同居群新疆野苹果的POD活性均呈现先增后降的变化趋势,在处理温度降至-5 ℃时均达到最大值,这说明,新疆野苹果在10~-5 ℃时细胞自身保护能力增强,而在低于-5 ℃时这种保护能力则变弱。不同居群新疆野苹果枝条的POD活性达到最高值时其由高到低依次为:托里县野苹果(269.44 μg·mg-1)、额敏县野苹果(255.06 μg·mg-1)、巩留县野苹果(231.92 μg·mg-1)、霍城县野苹果(186.79 μg·mg-1)、新源县野苹果(125.55 μg·mg-1)。与对照(CK)比较,不同居群新疆野苹果的POD活性达到最高时的增量由多到少依次为:托里县野苹果(234.60 μg·mg-1)、额敏县野苹果(210.31 μg·mg-1)、巩留县野苹果(176.24 μg·mg-1)、霍城县野苹果(125.81 μg·mg-1)、新源县野苹果(84.31 μg·mg-1)。显著性差异分析结果表明,当处理温度为-5 ℃时,托里县和额敏县野苹果的POD活性均极显著高于新源县和霍城县野苹果(P<0.01),巩留县野苹果的POD活性极显著高于新源县野苹果(P<0.01)。

图6 不同低温胁迫下不同居群新疆野苹果枝条的SOD活性Fig.6 SOD of different Malus sieversii populations under different treatment temperatures

图7 不同低温胁迫下不同居群新疆野苹果枝条的POD活性Fig.7 POD of different Malus sieversii populations under different treatment temperatures
2.8 不同低温胁迫对不同居群新疆野苹果枝条CAT活性的影响
不同低温胁迫下不同居群新疆野苹果枝条的CAT活性如图8所示。由图8可知,在低温胁迫下,不同居群新疆野苹果枝条的CAT活性均呈现先增后降的变化趋势,在处理温度降至0 ℃时均达到最大值,这说明不同居群新疆野苹果在10~0 ℃时其细胞自身保护能力迅速增强,而低于0 ℃时这种保护能力则变弱。不同居群新疆野苹果枝条的CAT活性最高值由高到低依次为:额敏县野苹果(192.22 U·g-1min-1)、托里县野苹果(176.74 U·g-1min-1)、霍城县野苹果(153.26 U·g-1min-1)、巩留县野苹果(148.28 U·g-1min-1)、新源县野苹果(133.56 U·g-1min-1)。与对照(CK)比较,不同居群新疆野苹果CAT活性达到最高值时的增量由多到少依次为:额敏县野苹果(151.50 U·g-1min-1)、托里县野苹果(136.52 U·g-1min-1)、巩留县野苹果(108.90 U·g-1min-1)、新源县野苹果(102.89 U·g-1min-1)、霍城县野苹果(102.37 U·g-1min-1)。显著性差异分析结果表明,当处理温度为0 ℃时,托里县和额敏县野苹果的CAT活性均极显著高于其他3个居群(P<0.01),巩留县野苹果的CAT活性显著高于新源县野苹果(P<0.05),新源县和霍城县野苹果的CAT活性之间差异不显著(P>0.05)。

图8 不同低温胁迫下不同居群新疆野苹果枝条的CAT活性Fig.8 CAT of different Malus sieversii populations under different treatment temperatures
2.9 不同低温胁迫下不同居群新疆野苹果枝条各抗寒指标的主成分分析
对不同居群新疆野苹果枝条各个抗寒指标测定值进行了主成分分析(结果见表1),从中提取出2个主成分,其累计贡献率为88.79%,包含了所测定的8个指标中的绝大部分信息,所以,可以利用这2个主成分对不同居群新疆野苹果进行抗寒性评价。在主成分1中,各个指标作用由大到小依次是CAT活性、可溶性糖含量、相对电导率、脯氨酸含量、SOD活性、POD活性、MDA含量和可溶性蛋白含量。
表2给出了不同居群新疆野苹果休眠枝条在所提取的主成分中的得分,分值越高,说明其抗寒性越强,由此可以推测,不同居群新疆野苹果的抗寒性由强到弱依次为:托里县野苹果、额敏县野苹果、巩留县野苹果、霍城县野苹果、新源县野苹果。
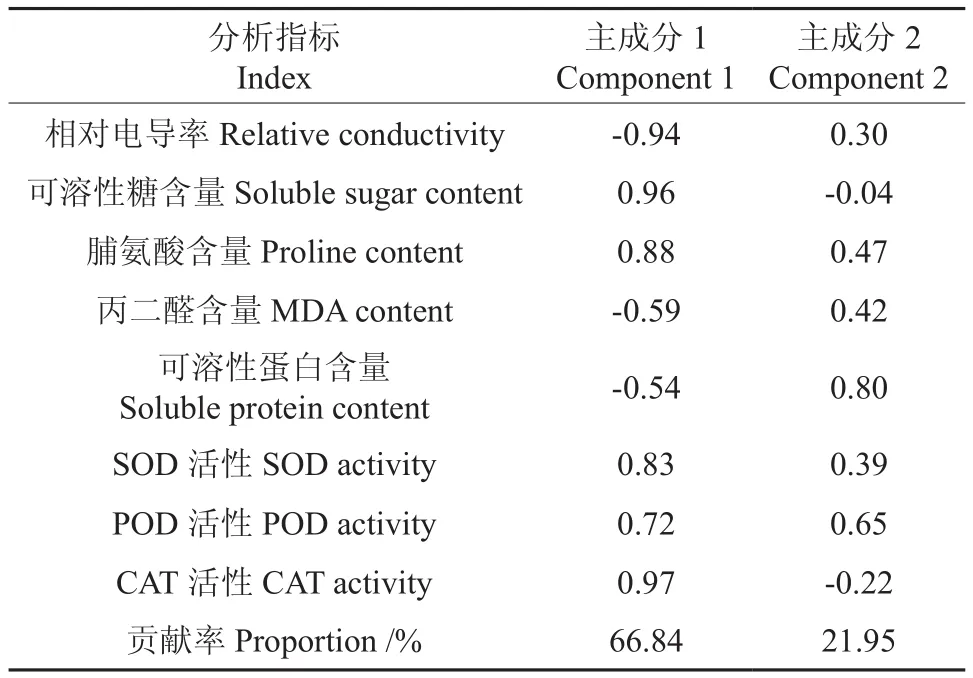
表1 新疆野苹果枝条各抗寒指标的主成分系数及贡献率Table 1 The component coefficient of the cold resistance indexes and proportion of Malus sieversii

表2 不同居群新疆野苹果主成分的得分Table 2 Factor scores of of Malus sieversii of different habitats
3 讨 论
相对电导率已广泛应用于植物抗寒性研究中,它能直观显示植物在低温胁迫下细胞膜透性的变化情况[19-21]。植物遭受低温伤害时,细胞的膜透性增大,进而导致生理代谢紊乱[22]。用相对电导率反映新疆野苹果细胞质膜透性的变化情况,能直观了解其受伤害的程度,推断不同居群新疆野苹果抗寒性的强弱顺序,探究其抗寒性差异。
可溶性糖、脯氨酸和可溶性蛋白均为植物体内重要的渗透调节物质。可溶性糖能通过渗透调节来增强细胞膜的稳定性[18]。研究中发现,不同居群新疆野苹果枝条的可溶性糖含量均随处理温度的下降而呈先升高后下降的变化趋势,表明新疆野苹果的抗冻能力在低温胁迫下先升高,但低温胁迫到达一定程度后则会变低,这与王勇等人对核桃抗寒性的研究结果一致[23]。脯氨酸能够增强植物的保水能力,从而增强植物的抗寒性[24]。研究中发现,不同居群新疆野苹果枝条的脯氨酸含量均随处理温度的下降而上升,表明其在受到低温胁迫时会调动内源物质来抵抗冻害,这与李建设等人对茄子幼苗抗寒性的研究结果一致[25]。可溶性蛋白不仅能降低植物细胞冰点,还能增加其水合度,增强其保水能力,减小细胞原生质受到脱水伤害[26],徐小军等人对甜瓜幼苗的研究结果表明,可溶性蛋白含量与试材的抗寒性呈正相关[27]。本研究结果表明,不同居群新疆野苹果的可溶性蛋白含量均随处理温度的下降而上升,但上升幅度不同,表明其抗寒性存在差异,这与王依等对不同品种葡萄的研究结果一致[28]。
丙二醛是植物在受到低温胁迫时膜脂过氧化作用的产物之一,其含量增加会破坏细胞膜系统,甚至导致细胞死亡[13,26]。本研究结果表明,不同居群新疆野苹果的MDA含量均随处理温度的下降而呈现先升后降的变化趋势,这与谭艳玲等人[29]对铁皮石斛的部分研究结果相近。
植物在受到低温胁迫时体内自由基和活性氧含量的增加会影响细胞膜系统的稳定性,最终导致膜结构被破坏,SOD、POD和CAT是植物体内的3种保护酶,都是酶促防御系统的重要组成部分,它们能维持膜的稳定,增强植物的抗寒性[30]。本研究中发现,不同居群新疆野苹果的SOD、POD和CAT酶活性均随处理温度的下降而呈先升后降的变化趋势,这表明,不同居群新疆野苹果的保护酶均受到低温的诱导,但处理温度降至一定程度后,3种保护酶的活性均开始下降,这说明其保护能力开始变弱,这与令凡等人对油橄榄的研究结果一致[31]。武雁军等人对厚皮甜瓜的研究结果表明,抗寒性强的品种其酶活性较高[14]。显著性差异分析结果表明,不同居群新疆野苹果的酶活性间有差异,因此可以通过不同居群新疆野苹果3种酶活性的大小来判断其抗寒性的强弱。
综上所述,不同居群新疆野苹果的抗寒性差异可以通过测定其相对电导率、可溶性糖含量、脯氨酸含量、MDA含量、可溶性蛋白含量、SOD活性、POD活性和CAT活性来推测。但是,植物的抗寒性受到多种因素的影响,只分析单一指标来反映其抗寒性并不可靠,不能用某一个指标评价植物的抗寒性[12,32,33]。目前,本课题组在前期已经研究了不同居群新疆野苹果叶片解剖结构的差异[8]、休眠枝条的生理响应差异[15-16]及与抗寒相关基因CBF1、CBF2、CBF3和CBF4的表达差异[34],这几个方面的研究结论均一致,且与本文的研究结论也一致,说明不同居群新疆野苹果的抗寒性存在差异,也证实了研究结论的可靠性,但要深刻而准确地揭示不同居群新疆野苹果的抗寒性差异,了解其差异产生的过程,还需在分子生物学层面进行更加深入的研究。
4 结 论
本研究对不同居群新疆野苹果枝条的相对电导率、可溶性糖含量、脯氨酸含量、丙二醛含量、可溶性蛋白含量、SOD活性、POD活性和CAT活性进行了测定,结合主成分分析结果对其抗寒性进行了综合评判,结果表明,不同居群新疆野苹果的抗寒性由强到弱依次为:托里县野苹果、额敏县野苹果、巩留县野苹果、霍城县野苹果、新源县野苹果。
[1]Wei X,Liu F,Chen C,Ma F,et al.TheMalus domesticasugar transporter gene family: identifications based on genome and expression profiling related to the accumµlation of fruit sugars[J].Frontiers in Plant Science,2014,5(Z1): 152-157.
[2]Chen X S,Feng T,Zhang Y M,et al.Genetic diversity of volatile components in Xinjiang Wild Apple (Malus sieversii)[J].Journal of Genetics and Genomics,2007,34(2): 171-179.
[3]阎国荣.主分量分析法在新疆野苹果与数种栽培品种亲缘关系研究中的应用[J].新疆环境保护 1997,19(1): 41- 45..
[4]Forte A V,Ignatov A N,Ponomarenko V V,et al.Phylogeny of theMalus(apple tree) species,inferred from the morphological traits and molecular DNA analysis[J].Genetics,2002,38(10):1150-1161.
[5]Richards C M,Volk G M,Reilley A A,et al.Genetic diversity and population structure inMalus sieversii,a wild progenitor species of domesticated apple[J].Tree Genetics & Genomes,2009,5(2): 339-347.
[6]Zhang Y M,Feng T,Zhang C Y,et al.Advances in research of theMalus sieversii(Lebed.) Roem.[J].Acta Horticulturae Sinica,2009,36(3): 447- 452.
[7]Zhang C Y,Chen X S,Zhang Y M,et al.Method of constructing core collection forMalus sieversiiin Xinjiang,China using molecular markers[J].Journal of Integrative Agriculture,2009,8(3): 276-284.
[8]王泽华,秦 伟,闫娟娟,等.不同居群新疆野苹果叶片解剖结构与抗寒性的关系[J].经济林研究,2016,34(3):163-168.
[9]王 磊.新疆野苹果和新疆野杏[J].新疆农业科学,1989(6):33-34.
[10]徐佳宁,刘 钢,马 盼,等.新疆野苹果逆境响应机制研究进展[J].山东农业科学,2014,46(8): 138-141.
[11]杨宁宁,孙万仓,刘自刚,等.北方冬油菜抗寒性的形态与生理机制[J].中国农业科学,2014,47(3):452-461.
[12]刘杜玲,张博勇,孙红梅,等.早实核桃不同品种抗寒性综合评价[J].园艺学报,2015,42(3): 545-553.
[13]陈新华,郭 婧,祁 雷,等.低温胁迫对甜樱桃一年生枝条的影响[J].果树学报,2014,31(增刊):124-128.
[14]武雁军,刘建辉.低温胁迫对厚皮甜瓜幼苗抗寒性生理生化指标的影响[J].西北农林科技大学学报(自然科学版),2007,35(3): 140-143.
[15]王泽华,秦 伟,马依努尔姑·吐地.不同生境新疆野苹果休眠枝条抗寒性鉴定[J].新疆农业大学学报,2016,39(2):106-111.
[16]王泽华,秦 伟,马依努尔姑·吐地.不同天然居群新疆野苹果休眠枝条可溶性蛋白和保护酶活性对低温的响应[J].新疆农业大学学报,2016,39(3):186-190.
[17]张莉梅,张子晗,喻方圆.低温层积过程中野鸦椿种子生理生化变化的研究[J].中南林业科技大学学报,2016,36(11):36-40.
[18]赵树亮.越橘越冬期枝条生理特性研究[D].哈尔滨:东北农业大学,2014.
[19]韩 轩,蔡 平,伊卫东,等.内蒙古8份冷蒿种质资源材料抗寒性的初步研究[J].草地学报,2014,22(3):669-672.
[20]林 艳,郭伟珍,徐振华,等.大叶女贞抗寒性及冬季叶片相对电导率变化研究[J].天津农业科学,2012,18(5):145-149.
[21]陈 曦,张 婷,刘志洋.低温胁迫对十种宿根花卉电导率的影响[J].北方园艺,2010(10): 121-122.
[22]葛亚英.两种苦首苔科花卉低温半致死温度的测定及耐寒生理变化研究[D].杭州:浙江大学,2010.
[23]王 勇,乔永胜,梅 霞,等.不同低温下核桃枝条抗寒性生理生化指标分析[J].中国农学通报,2013,29(10):40-44.
[24]Wang YL,Liao K,Liu J,et al.Changes of contents of osmosis substances and antioxidant enzyme activity in grape vines during cold exercise period before over-wintering[J].Journal of Fruit Science,2006,23(3):375- 378.
[25]李建设,耿广东,程智慧.低温胁迫对茄子幼苗抗寒性生理生化指标的影响[J].西北农林科技大学学报(自然科学版),2003,31(1):90-92.
[26]高京草,王慧霞,李西选.可溶性蛋白、丙二醛含量与枣树枝条抗寒性的关系研究[J].北方园艺,2010(23):18-20.
[27]徐小军,张桂兰,周亚峰,等.甜瓜幼苗耐冷性相关生理指标的综合评价[J].果树学报,2015,32(6):1187-1194.
[28]王 依,靳 娟,罗强勇,等.4个酿酒葡萄品种抗寒性的比较[J].果树学报,2015,32(4):612-619.
[29]谭艳玲,张艳嫣,高冬冬,等.低温胁迫对铁皮石斛抗坏血酸过氧化物酶活性及丙二醛和脯氨酸含量的影响[J].浙江大学学报(农业与生命科学版),2012,38(4):400-406.
[30]王玲丽.低温胁迫下不同百合的抗寒生理研究[D].武汉:华中农业大学,2012.
[31]令 凡,焦 健,李朝周,等.不同油橄榄品种对低温胁迫的生理响应及抗寒性综合评价[J].西北植物学报,2015,35(3):508-515.
[32]吕优伟,贺家圆,白小明,等.9个野生早熟禾对低温胁迫的生理响应及苗期抗寒性评价[J].草地学报,2014,22(2):326-333.
[33]余 萍,丁志彬,程龙霞,等.不同基质对欧洲鹅耳枥1 年生播种苗生长及生理特性影响[J].中南林业科技大学学报,2016,36(9):44-50,56.
[34]Wang Z H,Tian J,Geng W J,et al.Characterization of CBF1,CBF2,CBF3,and CBF4 genes ofMalus sieversiiand analysis of their expression in different habitats[J].European Journal of Horticultural Science,2017,82(2): 81-89.
猜你喜欢
河北科技师范学院学报(2022年2期)2022-08-26
热带作物学报(2022年5期)2022-06-01
浙江中医药大学学报(2021年6期)2021-07-12
中国粮油学报(2020年12期)2021-01-09
天津农业科学(2017年6期)2017-06-10
中国果菜(2016年9期)2016-03-01
中国马铃薯(2015年5期)2016-01-09
园艺与种苗(2015年10期)2015-02-27
农产品市场周刊(2014年15期)2014-08-22
植物资源与环境学报(2012年2期)2012-09-06
